
An In Vitro Orbital Flow Model to Study Mechanical Loading Effects on Osteoblasts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Osteoblasts Cultures
2.2. Orbital Shaker and Rocker Flow Models
2.3. Osteoblast Proliferation Assay
2.4. Differentiation Assay
2.5. Gene Expression
2.6. Immunofluorescence Labeling of Cells
2.7. Statistical Analysis
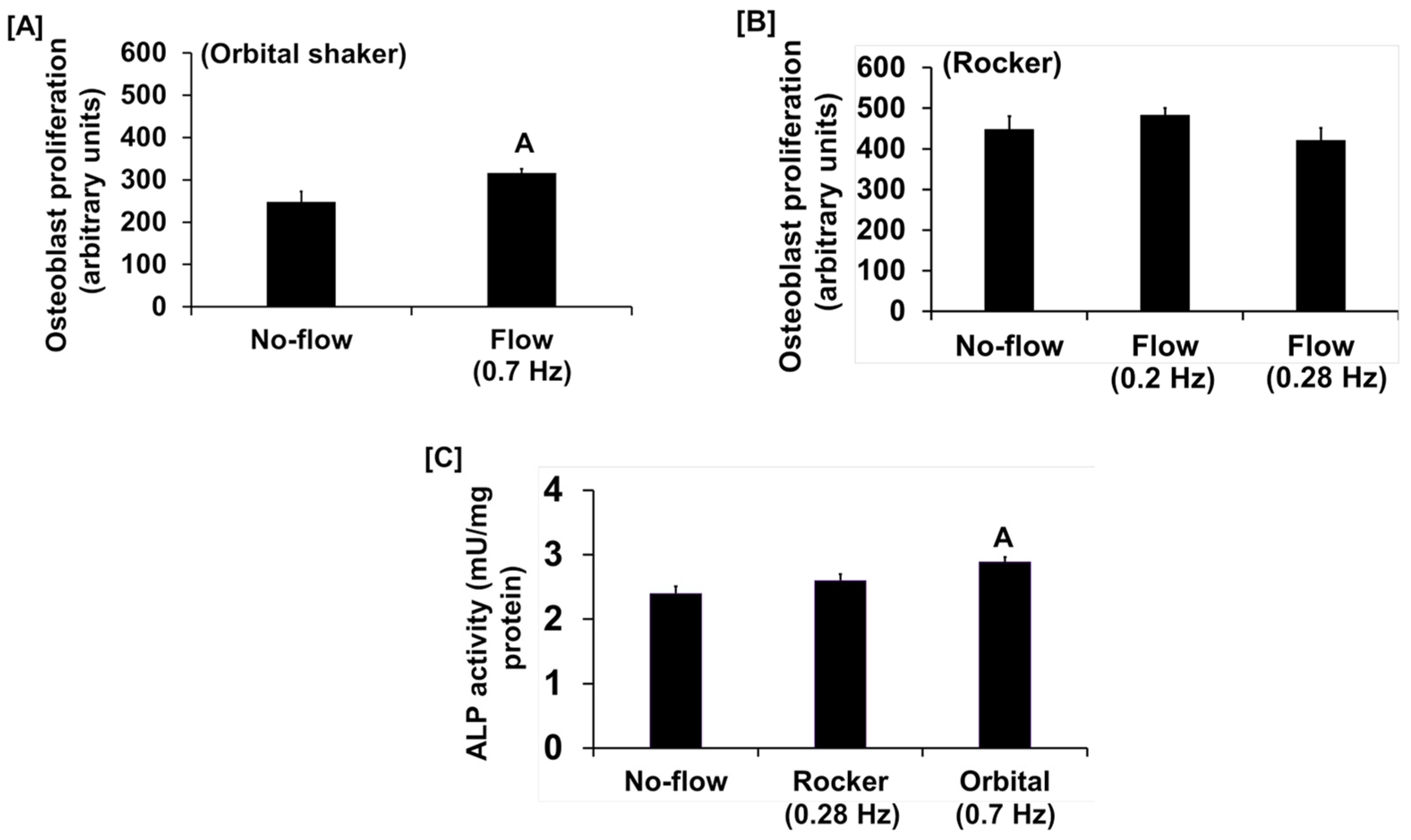
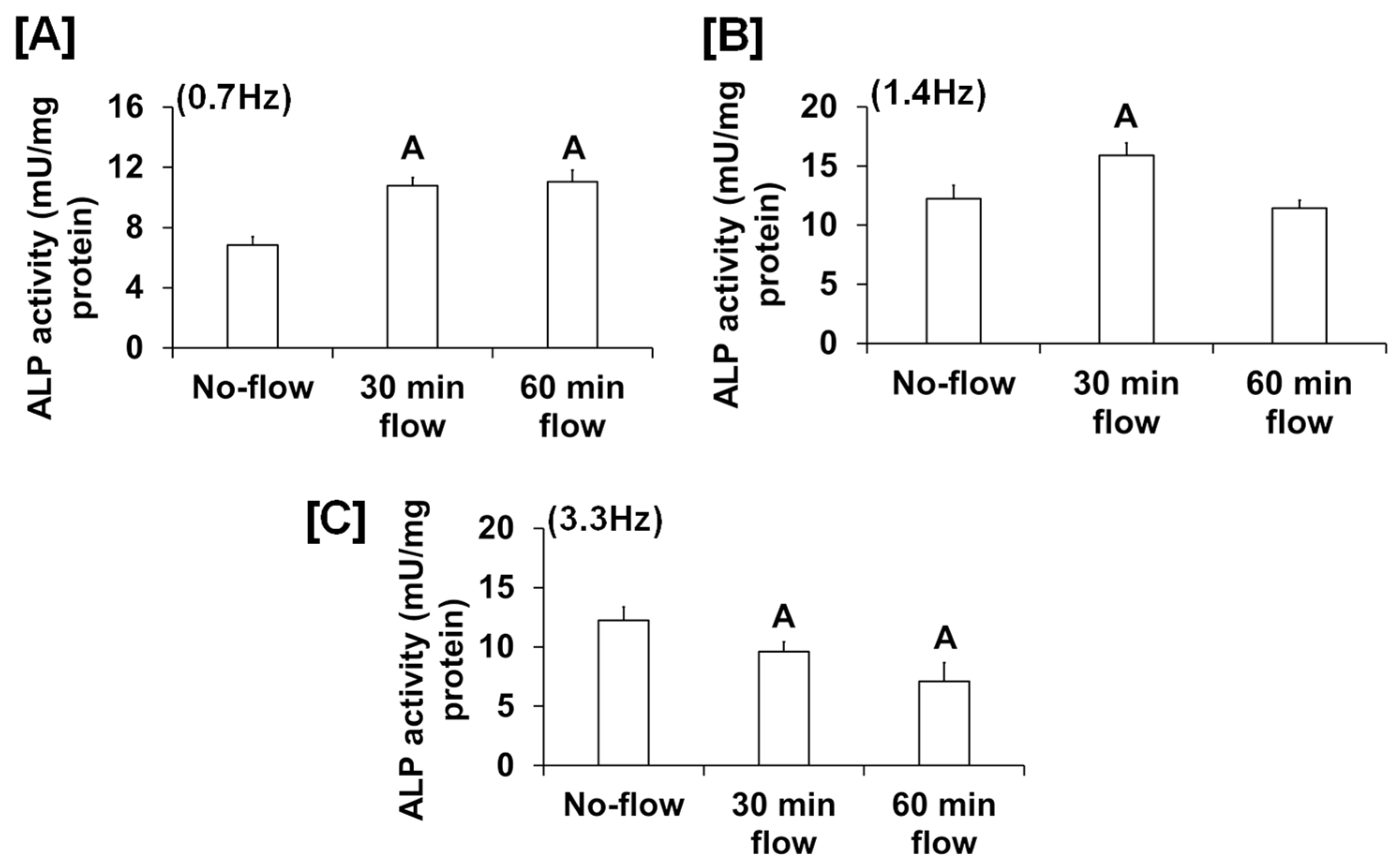
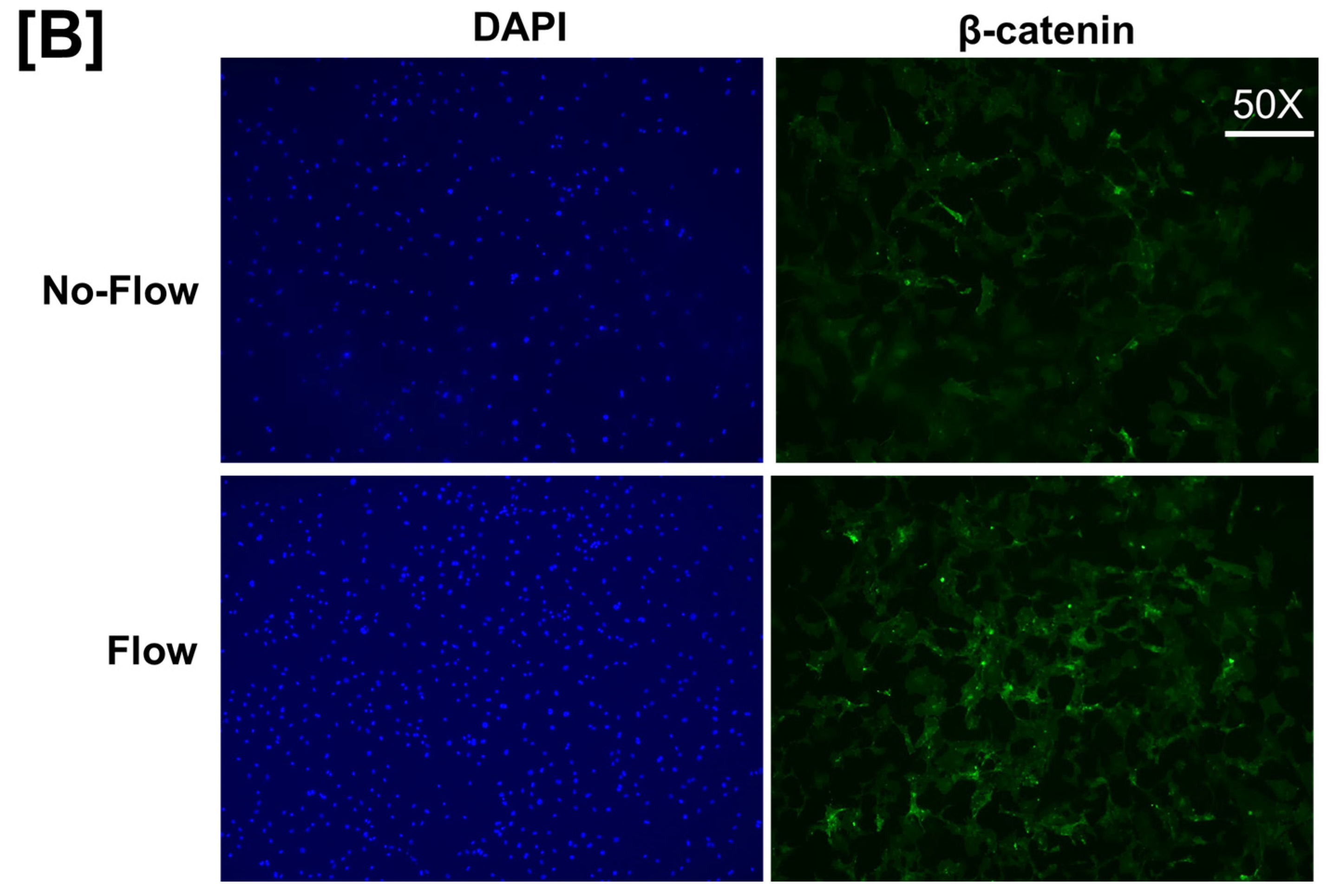
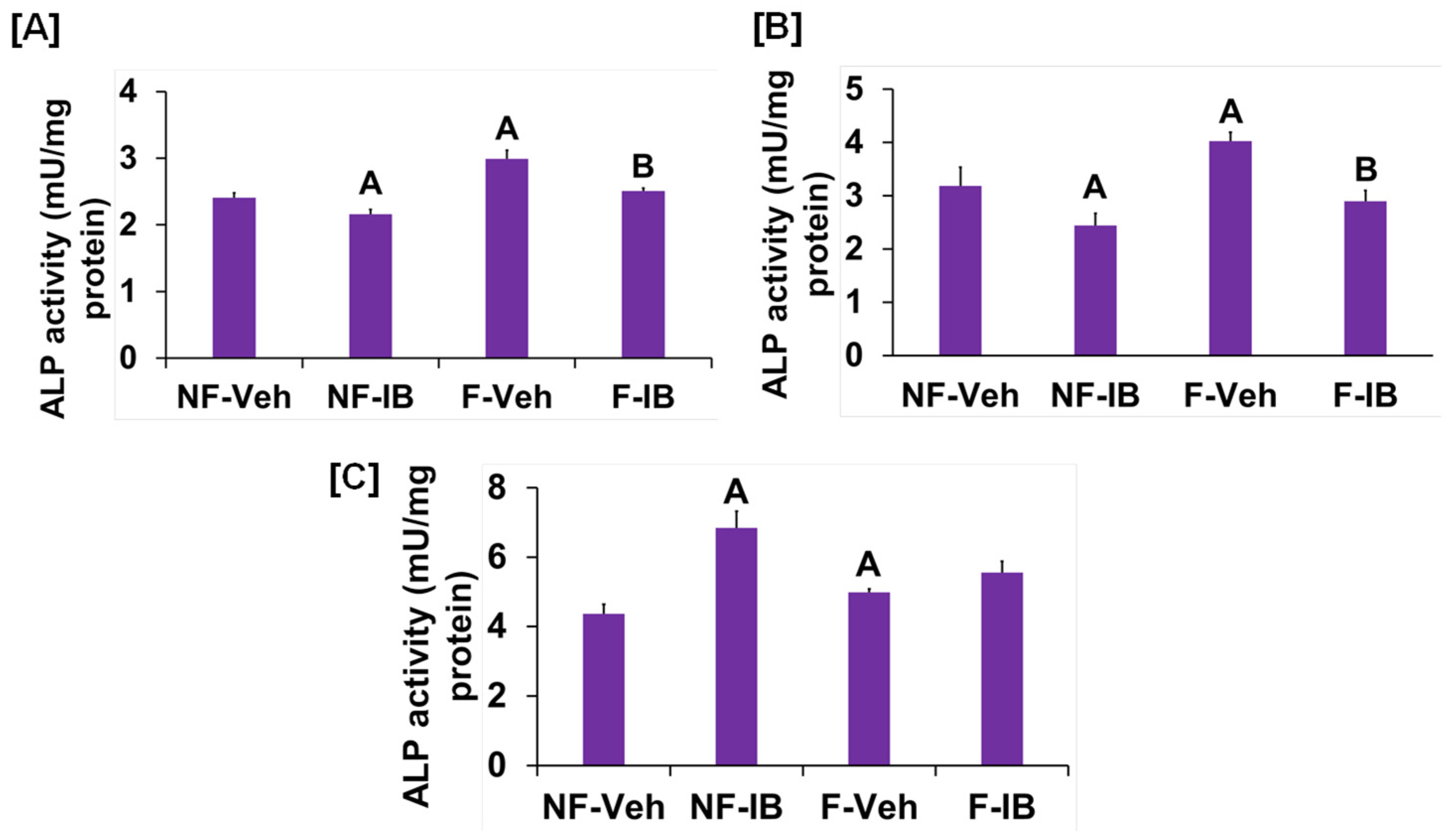
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergmann, P.; Body, J.J.; Boonen, S.; Boutsen, Y.; Devogelaer, J.P.; Goemaere, S.; Kaufman, J.; Reginster, J.Y.; Rozenberg, S. Loading and skeletal development and maintenance. J. Osteoporos. 2010, 2011, 786752. [Google Scholar] [CrossRef]
- Chang, X.; Xu, S.; Zhang, H. Regulation of bone health through physical exercise: Mechanisms and types. Front. Endocrinol. 2022, 13, 1029475. [Google Scholar] [CrossRef]
- Howe, T.E.; Shea, B.; Dawson, L.J.; Downie, F.; Murray, A.; Ross, C.; Harbour, R.T.; Caldwell, L.M.; Creed, G. Exercise for preventing and treating osteoporosis in postmenopausal women. Cochrane Database Syst. Rev. 2011, 7, CD000333. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.T.; Lanyon, L.E. Regulation of bone mass by mechanical strain magnitude. Calcif. Tissue Int. 1985, 37, 411–417. [Google Scholar] [CrossRef]
- Kesavan, C.; Wergedal, J.E.; Lau, K.H.; Mohan, S. Conditional disruption of IGF-I gene in type 1alpha collagen-expressing cells shows an essential role of IGF-I in skeletal anabolic response to loading. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1191–E1197. [Google Scholar] [CrossRef]
- Hughes, J.M.; Guerriere, K.I.; Popp, K.L.; Castellani, C.M.; Pasiakos, S.M. Exercise for optimizing bone health after hormone-induced increases in bone stiffness. Front. Endocrinol. 2023, 14, 1219454. [Google Scholar] [CrossRef]
- Song, L. Effects of Exercise or Mechanical Stimulation on Bone Development and Bone Repair. Stem Cells Int. 2022, 2022, 5372229. [Google Scholar] [CrossRef] [PubMed]
- Eimori, K.; Endo, N.; Uchiyama, S.; Takahashi, Y.; Kawashima, H.; Watanabe, K. Disrupted Bone Metabolism in Long-Term Bedridden Patients. PLoS ONE 2016, 11, e0156991. [Google Scholar] [CrossRef] [PubMed]
- Jaul, E.; Malcov, T.; Menczel, J. Osteoporosis in tube-fed bed-ridden elderly female patients. J. Am. Geriatr. Soc. 2009, 57, 1318–1320. [Google Scholar] [CrossRef]
- Tafaro, L.; Napoli, N. Current and Emerging Treatment of Osteoporosis. In Orthogeriatrics: The Management of Older Patients with Fragility Fractures, 2nd ed.; Falaschi, P., Marsh, D., Eds.; Springer: Cham, Switzerland, 2021; pp. 257–272. [Google Scholar]
- Benedetti, M.G.; Furlini, G.; Zati, A.; Letizia Mauro, G. The Effectiveness of Physical Exercise on Bone Density in Osteoporotic Patients. BioMed Res. Int. 2018, 2018, 4840531. [Google Scholar] [CrossRef]
- Hong, A.R.; Kim, S.W. Effects of Resistance Exercise on Bone Health. Endocrinol. Metab. 2018, 33, 435–444. [Google Scholar] [CrossRef]
- Battaglino, R.A.; Lazzari, A.A.; Garshick, E.; Morse, L.R. Spinal cord injury-induced osteoporosis: Pathogenesis and emerging therapies. Curr. Osteoporos. Rep. 2012, 10, 278–285. [Google Scholar] [CrossRef]
- Michael Delaine-Smith, R.; Javaheri, B.; Helen Edwards, J.; Vazquez, M.; Rumney, R.M. Preclinical models for in vitro mechanical loading of bone-derived cells. Bonekey Rep. 2015, 4, 728. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaka, T.; Matsugaki, A.; Nakano, T. Control of osteoblast arrangement by osteocyte mechanoresponse through prostaglandin E2 signaling under oscillatory fluid flow stimuli. Biomaterials 2021, 279, 121203. [Google Scholar] [CrossRef] [PubMed]
- Farley, J.R.; Hall, S.L.; Tanner, M.A.; Wergedal, J.E. Specific activity of skeletal alkaline phosphatase in human osteoblast-line cells regulated by phosphate, phosphate esters, and phosphate analogs and release of alkaline phosphatase activity inversely regulated by calcium. J. Bone Miner. Res. 1994, 9, 497–508. [Google Scholar] [CrossRef]
- Mohan, S.; Kesavan, C. T-cell factor 7L2 is a novel regulator of osteoblast functions that acts in part by modulation of hypoxia signaling. Am. J. Physiol. Endocrinol. Metab. 2022, 322, E528–E539. [Google Scholar] [CrossRef]
- Dardik, A.; Chen, L.; Frattini, J.; Asada, H.; Aziz, F.; Kudo, F.A.; Sumpio, B.E. Differential effects of orbital and laminar shear stress on endothelial cells. J. Vasc. Surg. 2005, 41, 869–880. [Google Scholar] [CrossRef]
- Wittkowske, C.; Reilly, G.C.; Lacroix, D.; Perrault, C.M. In Vitro Bone Cell Models: Impact of Fluid Shear Stress on Bone Formation. Front. Bioeng. Biotechnol. 2016, 4, 87. [Google Scholar] [CrossRef]
- Silva, M.J.; Brodt, M.D.; Lynch, M.A.; Stephens, A.L.; Wood, D.J.; Civitelli, R. Tibial loading increases osteogenic gene expression and cortical bone volume in mature and middle-aged mice. PLoS ONE 2012, 7, e34980. [Google Scholar] [CrossRef]
- McKenzie, J.A.; Bixby, E.C.; Silva, M.J. Differential gene expression from microarray analysis distinguishes woven and lamellar bone formation in the rat ulna following mechanical loading. PLoS ONE 2011, 6, e29328. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yu, W.; Liu, Y.; Yuan, X.; Yuan, R.; Guo, Q. Expression of HIF-1alpha in cycling stretch-induced osteogenic differentiation of bone mesenchymal stem cells. Mol. Med. Rep. 2019, 20, 4489–4498. [Google Scholar] [PubMed]
- Lau, K.H.; Kapur, S.; Kesavan, C.; Baylink, D.J. Up-regulation of the Wnt, estrogen receptor, insulin-like growth factor-I, and bone morphogenetic protein pathways in C57BL/6J osteoblasts as opposed to C3H/HeJ osteoblasts in part contributes to the differential anabolic response to fluid shear. J. Biol. Chem. 2006, 281, 9576–9588. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Ito, M.; Naoe, Y.; Lacy-Hulbert, A.; Ikeda, K. Integrin alphav in the mechanical response of osteoblast lineage cells. Biochem. Biophys. Res. Commun. 2014, 447, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Bakker, A.D.; Gakes, T.; Hogervorst, J.M.; de Wit, G.M.; Klein-Nulend, J.; Jaspers, R.T. Mechanical Stimulation and IGF-1 Enhance mRNA Translation Rate in Osteoblasts via Activation of the AKT-mTOR Pathway. J. Cell. Physiol. 2016, 231, 1283–1290. [Google Scholar] [CrossRef]
- Song, F.; Wang, Y.; Jiang, D.; Wang, T.; Zhang, Y.; Ma, H.; Kang, Y. Cyclic Compressive Stress Regulates Apoptosis in Rat Osteoblasts: Involvement of PI3K/Akt and JNK MAPK Signaling Pathways. PLoS ONE 2016, 11, e0165845. [Google Scholar] [CrossRef]
- Robinson, J.A.; Chatterjee-Kishore, M.; Yaworsky, P.J.; Cullen, D.M.; Zhao, W.; Li, C.; Kharode, Y.; Sauter, L.; Babij, P.; Brown, E.L.; et al. Wnt/beta-catenin signaling is a normal physiological response to mechanical loading in bone. J. Biol. Chem. 2006, 281, 31720–31728. [Google Scholar] [CrossRef]
- Kopf, J.; Petersen, A.; Duda, G.N.; Knaus, P. BMP2 and mechanical loading cooperatively regulate immediate early signalling events in the BMP pathway. BMC Biol. 2012, 10, 37. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohan, S.; Surisetty, R.; Kesavan, C. An In Vitro Orbital Flow Model to Study Mechanical Loading Effects on Osteoblasts. Biology 2024, 13, 646. https://doi.org/10.3390/biology13090646
Mohan S, Surisetty R, Kesavan C. An In Vitro Orbital Flow Model to Study Mechanical Loading Effects on Osteoblasts. Biology. 2024; 13(9):646. https://doi.org/10.3390/biology13090646
Chicago/Turabian StyleMohan, Subburaman, Ritika Surisetty, and Chandrasekhar Kesavan. 2024. "An In Vitro Orbital Flow Model to Study Mechanical Loading Effects on Osteoblasts" Biology 13, no. 9: 646. https://doi.org/10.3390/biology13090646
APA StyleMohan, S., Surisetty, R., & Kesavan, C. (2024). An In Vitro Orbital Flow Model to Study Mechanical Loading Effects on Osteoblasts. Biology, 13(9), 646. https://doi.org/10.3390/biology13090646