
Bird Richness and Abundance in Urban Areas: Simulation-Based Conservation Strategies for an Italian Town


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Bird Surveys
2.2. Sampling Completeness
2.3. Environmental Filtering
2.4. Generalized Additive Models
2.5. What-If Simulations
3. Results
3.1. Bird Surveys
3.2. Sampling Completeness
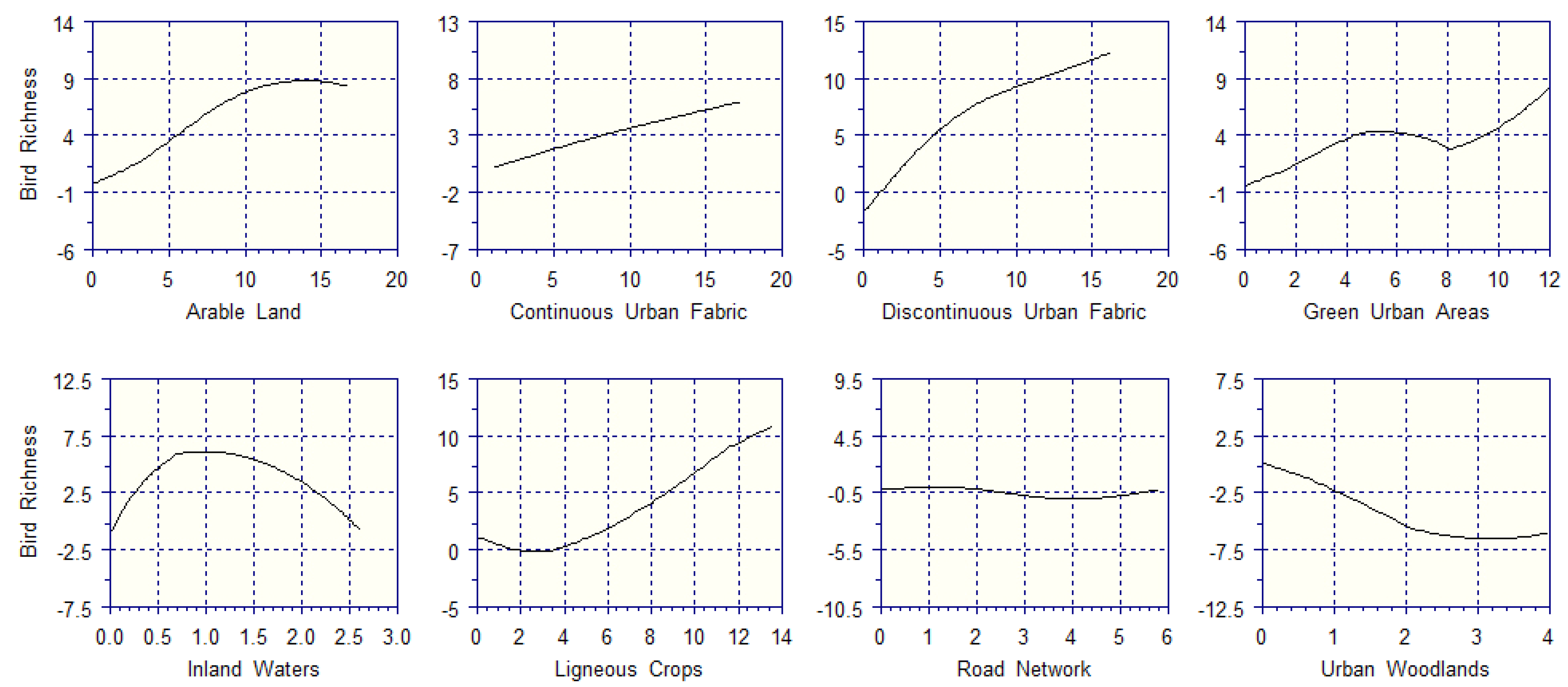
3.3. Environmental Filtering
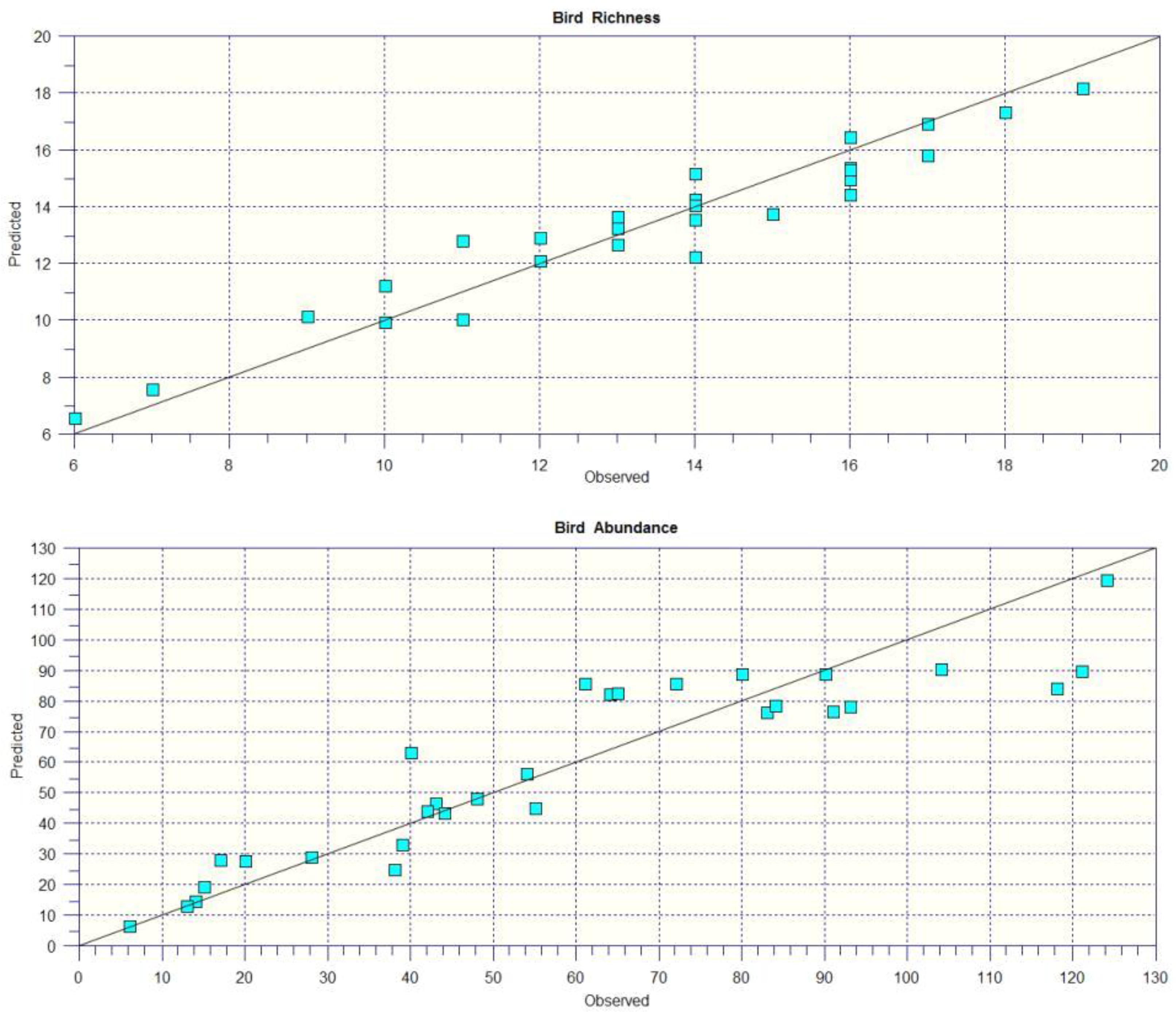
3.4. GAMs
3.5. What-If Simulations
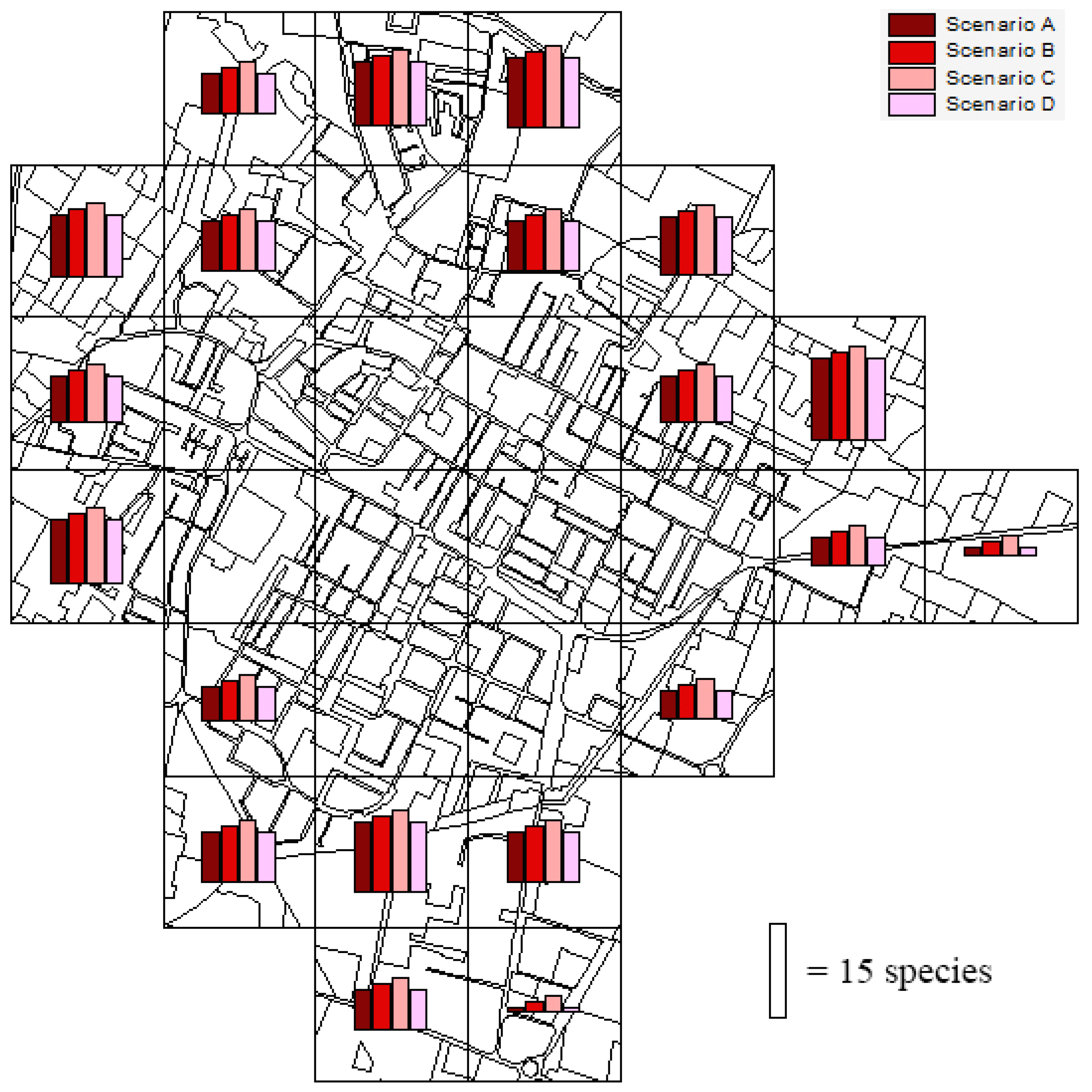
3.5.1. Urban Expansion
3.5.2. Urban Greening
4. Discussion
Entailments for Avian Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischer, S.E.; Edwards, A.C.; Weber, P.; Garnett, S.T.; Whiteside, T.G. The Bird Assemblage of the Darwin Region (Australia): What Is the Effect of Twenty Years of Increasing Urbanisation? Diversity 2021, 13, 294. [Google Scholar] [CrossRef]
- Bisikirskienė, L.; Brazaitis, G.; Šimkevičius, K.; Brazaitytė, G. How Are Urban Birds Affected by Surrounding Forests and Agricultural Landscapes? Forests 2023, 14, 2119. [Google Scholar] [CrossRef]
- Evans, B.S.; Reitsma, R.; Hurlbert, A.H.; Marra, P.P. Environmental filtering of avian communities along a rural-to-urban gradient in Greater Washington, DC, USA. Ecosphere 2018, 9, e02402. [Google Scholar] [CrossRef]
- Hensley, C.B.; Trisos, C.H.; Warren, P.S.; MacFarland, J.; Blumenshine, S.; Reece, J.; Katti, M. Effects of Urbanization on Native Bird Species in Three Southwestern US Cities. Front. Ecol. Evol. 2019, 7, 71. [Google Scholar] [CrossRef]
- Simkin, R.D.; Seto, K.C.; McDonald, R.I.; Jetz, W. Biodiversity Impacts and Conservation Implications of Urban Land Expansion Projected to 2050. Proc. Natl. Acad. Sci. USA 2022, 119, e2117297119. [Google Scholar] [CrossRef] [PubMed]
- Murgui, E.; Hedblom, M. Ecology and Conservation of Birds in Urban Environments; Springer: New York, NY, USA, 2017. [Google Scholar]
- Evans, K.L.; Chamberlain, D.E.; Hatchwell, B.J.; Gregory, R.D.; Gaston, K.J. What Makes an Urban Bird? Glob. Chang. Biol. 2011, 17, 32–44. [Google Scholar] [CrossRef]
- Blair, R.B. Land use and avian species diversity along an urban gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar] [CrossRef]
- Gil, D.; Brumm, H. Avian Urban Ecology; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the City: Fundamental questions for understanding the ecology of urban green spaces for biodiversity conservation. Bioscience 2017, 67, 799–807. [Google Scholar] [CrossRef]
- Andrade, R.; Bateman, H.L.; Franklin, J.; Allen, D. Waterbird community composition, abundance, and diversity along an urban gradient. Landsc. Urban Plan. 2018, 170, 103–111. [Google Scholar] [CrossRef]
- Minias, P.; Janiszewski, T. Ground nesting in passerine birds: Evolution, biogeography and life history correlates. Oikos 2023, 6, e09870. [Google Scholar] [CrossRef]
- Storchová, L.; Hořák, D. Life-history characteristics of european birds. Glob. Ecol. Biogeogr. 2018, 27, 400–406. [Google Scholar] [CrossRef]
- Sol, D.; González-Lagos, C.; Moreira, D.; Maspons, J.; Lapiedra, O. Urbanisation tolerance and the loss of avian diversity. Ecol. Lett. 2014, 17, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Jokimäki, J.; Suhonen, J.; Jokimäki-Kaisanlahti, M.-L.; Carbó-Ramírez, P. Effects of urbanization on breeding birds in european towns: Impacts of species traits. Urban Ecosyst. 2016, 19, 1565–1577. [Google Scholar] [CrossRef]
- Fernández-Juricic, E. Can human disturbance promote nestedness? a case study with breeding birds in urban habitat fragments. Oecologia 2002, 131, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L. Spatial similarity of urban bird communities: A multiscale approach. J. Biogeogr. 2003, 30, 1183–1193. [Google Scholar] [CrossRef]
- Kath, J.; Maron, M.; Dunn, P.K. Interspecific competition and small bird diversity in an urbanizing landscape. Landsc. Urban Plan. 2009, 92, 72–79. [Google Scholar] [CrossRef]
- Gilbert, G.; Gibbons, D.W.; Evans, J. Bird Monitoring Methods: A Manual of Techniques for Key UK Species; Royal Society for the Protection of Birds: Bedfordshire, UK, 1988. [Google Scholar]
- Cramp, S.E.; Simmons, K.E.L.; Brooks, D.J.; Perrins, C.M. Handbook of the Birds of Europe, the Middle East and North Africa: The Birds of the Western Palearctic (Vol. 1–9); Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Berthold, P. Bird Migration: A General Survey; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Thompson, S.K. Sampling; Wiley: New York, NY, USA, 1992. [Google Scholar]
- Peterson, A.T.; Slade, N.A. Extrapolating inventory results into biodiversity estimates and the importance of stopping rules. Divers. Distrib. 1998, 4, 95–105. [Google Scholar] [CrossRef]
- Soberón, M.J.; Llorente, B.J. The use of species accumulation functions for the prediction of species richness. Conserv. Biol. 1993, 7, 480–488. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Burham, K.P.; Overton, W.S. Estimation of the size of a closed population when capture probabilities vary among individuals. Biometrika 1978, 65, 625. [Google Scholar]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B 1994, 345, 101–118. [Google Scholar]
- Connor, E.F.; Simberloff, D. The Assembly of Species Communities: Chance or Competition? Ecology 1979, 60, 1132–1140. [Google Scholar] [CrossRef]
- Connor, E.F.; Simberloff, D. Competition, scientific method, and null models in ecology. Am. Sci. 1986, 74, 155–162. [Google Scholar]
- Ladau, J.; Schwager, S.J. Robust Hypothesis Tests for Independence in Community Assembly. J. Math. Biol. 2008, 57, 555. [Google Scholar] [CrossRef]
- Stone, L.; Roberts, A. The checkerboard score and species distributions. Oecologia 1990, 85, 74–79. [Google Scholar] [CrossRef]
- Hastie, T.J.; Tibshirani, R.J. Generalised Additive Models; Chapman and Hall: New York, NY, USA, 1990. [Google Scholar]
- Palacio, F.X.; Ibañez, L.M.; Maragliano, R.E.; Montalti, D. Urbanization as a driver of taxonomic, functional, and phylogenetic diversity losses in bird communities. Can. J. Zool. 2018, 96, 1114–1121. [Google Scholar] [CrossRef]
- Donald, P.F.; Sanderson, F.J.; Burfield, I.J.; Van Bommel, F.P.J. Further evidence of continent-wide impacts of agricultural intensification on european farmland birds, 1990–2000. Agric. Ecosyst. Environ. 2006, 116, 189–196. [Google Scholar] [CrossRef]
- Sol, D.; Bartomeus, I.; González-Lagos, C.; Pavoine, S. Urbanisation and the loss of phylogenetic diversity in birds. Ecol. Lett. 2017, 20, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Sol, D.; Trisos, C.; Múrria, C.; Jeliazkov, A.; González-Lagos, C.; Pigot, A.L.; Ricotta, C.; Swan, C.M.; Tobias, J.A.; Pavoine, S. The worldwide impact of urbanisation on avian functional diversity. Ecol. Lett. 2020, 23, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Korányi, D.; Gallé, R.; Donkó, B.; Chamberlain, D.E.; Batáry, P. Urbanization does not affect green space bird species richness in a mid-sized city. Urban Ecosyst. 2021, 24, 789–800. [Google Scholar] [CrossRef]
- Chamberlain, D.; Kibuule, M.; Skeen, R.; Pomeroy, D. Trends in bird species richness, abundance and biomass along a tropical urbanization gradient. Urban Ecosyst. 2017, 20, 629–638. [Google Scholar] [CrossRef]
- Filloy, J.; Zurita, G.A.; Bellocq, M.I. Bird diversity in urban ecosystems: The role of the biome and land use along urbanization gradients. Ecosystems 2019, 22, 213–227. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Major, R.E.; Lyons, M.B.; Martin, J.M.; Kingsford, R.T. The effects of local and landscape habitat attributes on bird diversity in urban greenspaces. Ecosphere 2018, 9, e02347. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Poore, A.G.B.; Major, R.E.; Cornwell, W.K.; Wilshire, J.H.; Lyons, M.B. How to build a bio-diverse city: Environmental determinants of bird diversity within and among 1581 cities. Biodivers. Conserv. 2021, 30, 217–234. [Google Scholar] [CrossRef]
- Evans, K.L.; Newson, S.E.; Gaston, K.J. Habitat influences on urban avian assemblages. Ibis 2009, 151, 19–39. [Google Scholar] [CrossRef]
- Kang, W.; Minor, E.S.; Park, C.-R.; Lee, D. Effects of Habitat Structure, Human Disturbance, and Habitat Connectivity on Urban Forest Bird Communities. Urban Ecosyst. 2015, 18, 857–870. [Google Scholar] [CrossRef]
- Zhou, D.; Chu, L.M. How Would Size, Age, Human Disturbance, and Vegetation Structure Affect Bird Communities of Urban Parks in Different Seasons? J. Ornithol. 2012, 153, 1101–1112. [Google Scholar] [CrossRef]
- Haaland, C.; van den Bosch, C.K. Challenges and Strategies for Urban Green-Space Planning in Cities Undergoing Densification: A Review. Urban For. Urban Green. 2015, 14, 760–771. [Google Scholar] [CrossRef]
- Mexia, T.; Vieira, J.; Príncipe, A.; Anjos, A.; Silva, P.; Lopes, N.; Freitas, C.; Santos-Reis, M.; Correia, O.; Branquinho, C.; et al. Ecosystem Services: Urban Parks under a Magnifying Glass. Environ. Res. 2018, 160, 469–478. [Google Scholar] [CrossRef]
- Reif, J.; Hanzelka, J. Continent-wide gradients in open-habitat insectivorous bird declines track spatial patterns in agricultural intensity across Europe. Glob. Ecol. Biogeogr. 2020, 29, 1988–2013. [Google Scholar] [CrossRef]
- Donald, P.F.; Green, R.E.; Heath, M.F. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 25–29. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimator | Predicted Bird Richness (Session 1) | % Completeness (Session 1) | Predicted Bird Richness (Session 2) | % Completeness (Session 2) |
|---|---|---|---|---|
| First-order jackknife | 37.80 | 84.66 | 37.83 | 87.23 |
| Second-order jackknife | 38.68 | 82.73 | 37.99 | 86.86 |
| Chao1 | 38.43 | 83.27 | 37.18 | 88.76 |
| Michaelis–Menten | 32.11 | 99.66 | 33.68 | 97.98 |
| Sampling Session 1 | Sampling Session 2 | |
|---|---|---|
| Assumption 1: 0% of species interacting | ||
| p-value | p < 0.001 | p = 0.037 |
| 95% confidence interval for p-value | 0 < p < 0.001 | 0.003 < p < 0.074 |
| Assumption 2: 50% of species interacting | ||
| p-value | p < 0.001 | p = 0.059 |
| 95% confidence interval for p-value | 0 < p < 0.001 | 0.031 < p < 0.084 |
| Assumption 3: 100% of species interacting | ||
| p-value | p < 0.001 | p = 0.064 |
| 95% confidence interval for p-value | 0 < p < 0.001 | 0.040 < p < 0.094 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrarini, A.; Bagni, L.; Gustin, M. Bird Richness and Abundance in Urban Areas: Simulation-Based Conservation Strategies for an Italian Town. Biology 2025, 14, 37. https://doi.org/10.3390/biology14010037
Ferrarini A, Bagni L, Gustin M. Bird Richness and Abundance in Urban Areas: Simulation-Based Conservation Strategies for an Italian Town. Biology. 2025; 14(1):37. https://doi.org/10.3390/biology14010037
Chicago/Turabian StyleFerrarini, Alessandro, Luca Bagni, and Marco Gustin. 2025. "Bird Richness and Abundance in Urban Areas: Simulation-Based Conservation Strategies for an Italian Town" Biology 14, no. 1: 37. https://doi.org/10.3390/biology14010037
APA StyleFerrarini, A., Bagni, L., & Gustin, M. (2025). Bird Richness and Abundance in Urban Areas: Simulation-Based Conservation Strategies for an Italian Town. Biology, 14(1), 37. https://doi.org/10.3390/biology14010037