
Microbial Seafloor Weathering of Hydrothermal Sulfides: Insights from an 18-Month In Situ Incubation at the Wocan-1 Hydrothermal Field


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
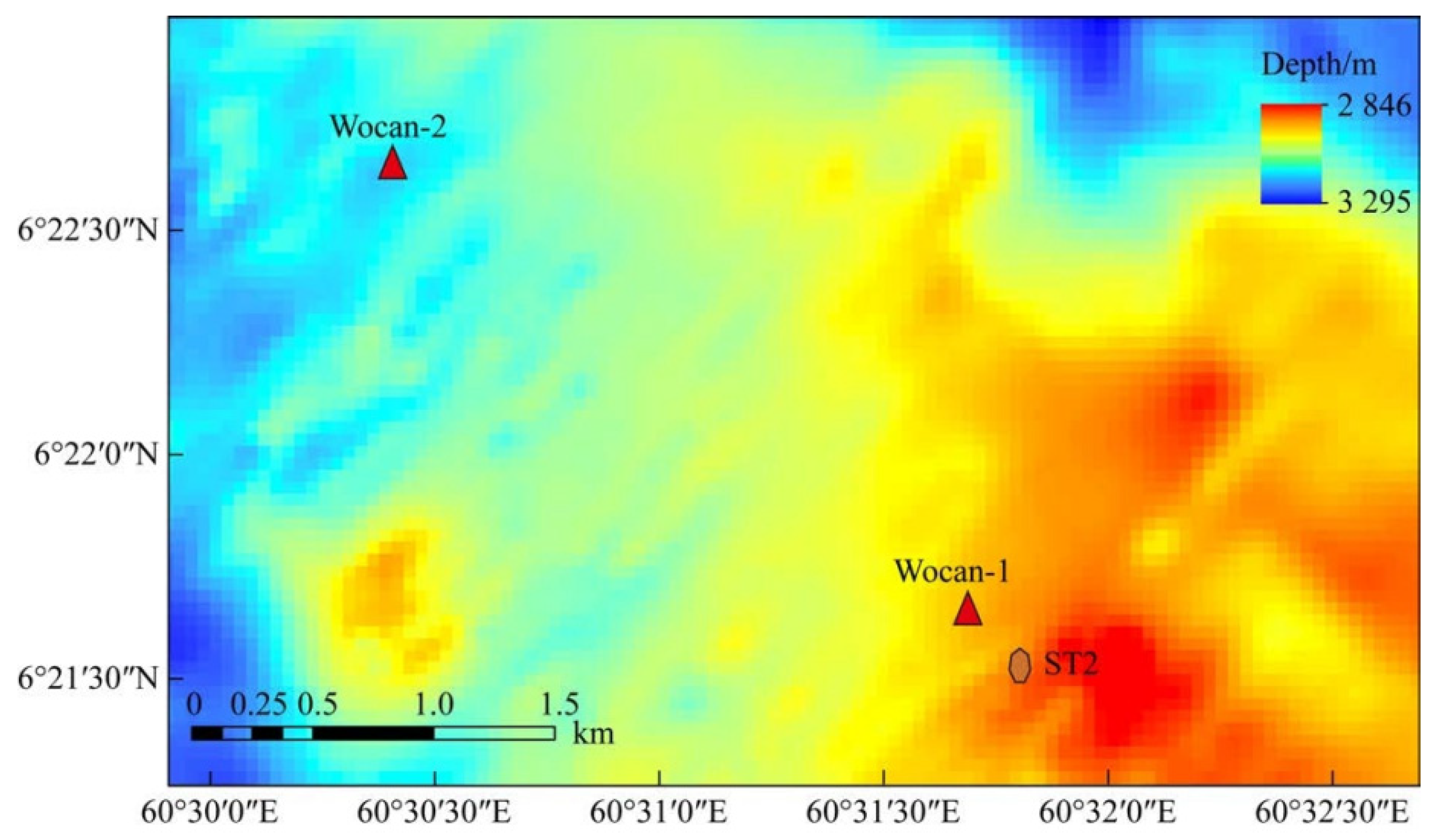
2.1. Sample Selection
2.2. In Situ Incubation Experiments on the Seafloor
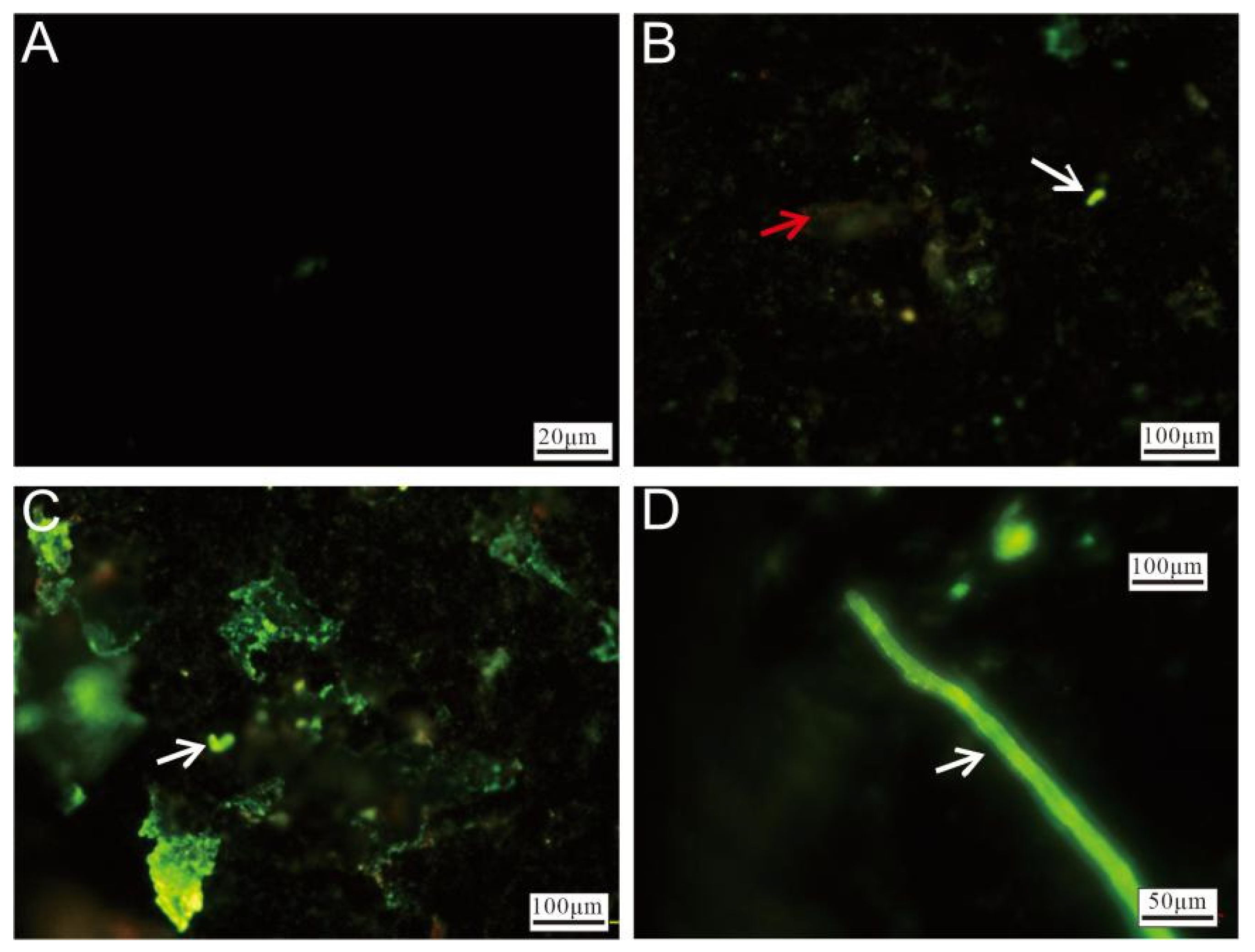
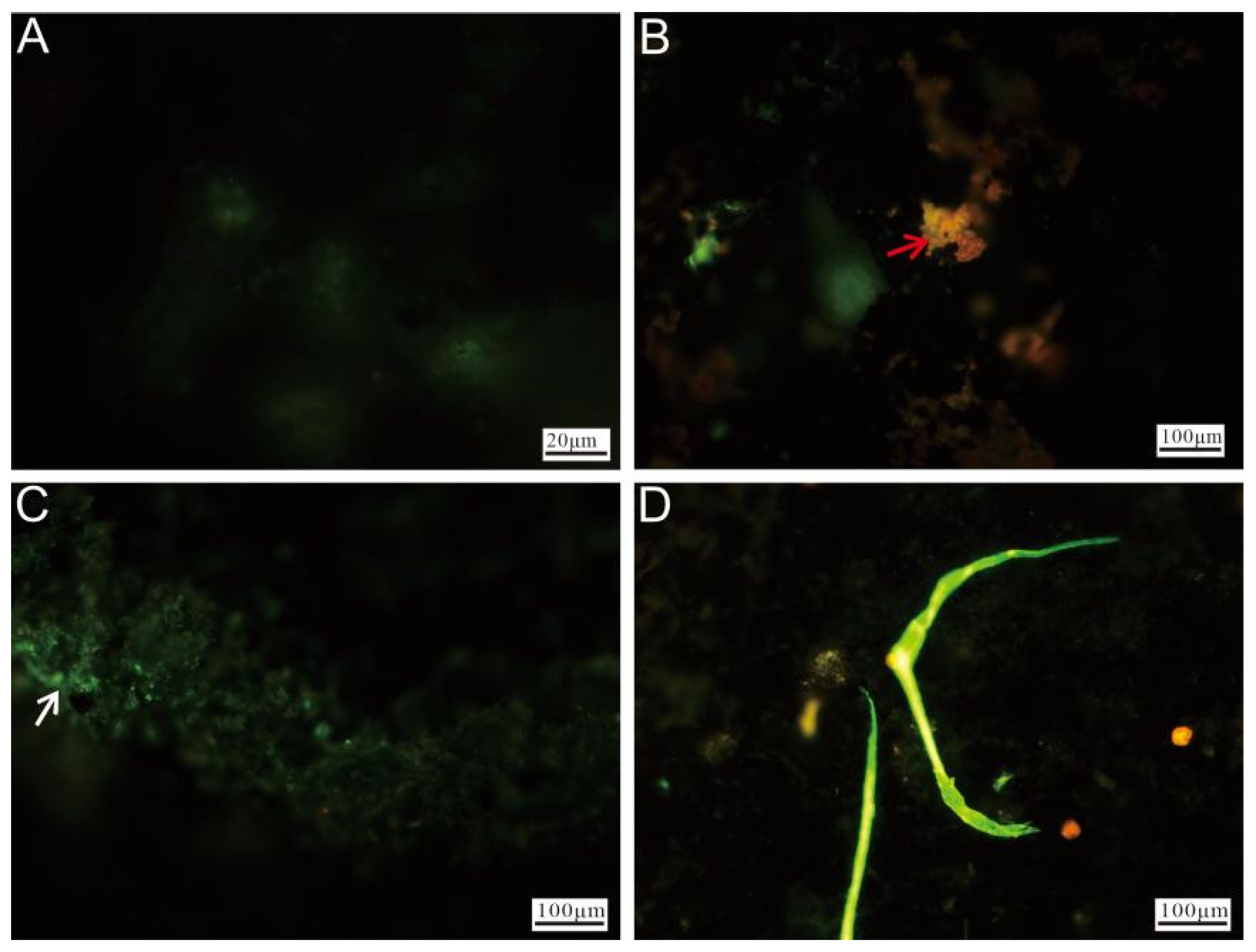
2.3. Cell Staining and Microstructure Analysis
2.4. FT-IR Analysis
3. Results
3.1. Surface Morphology Changes Before and After the Incubation Experiment
3.2. Observations Under SEM
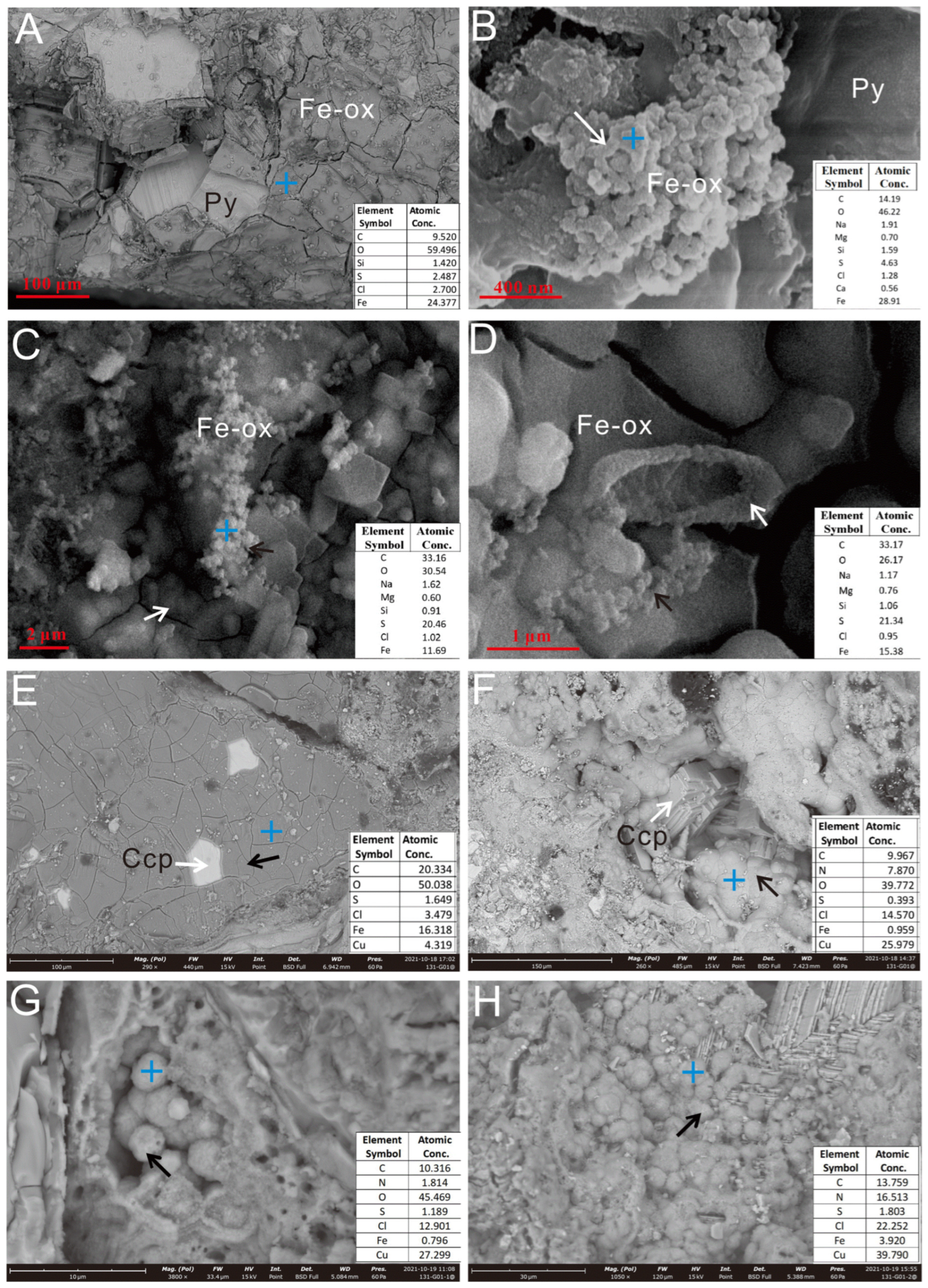
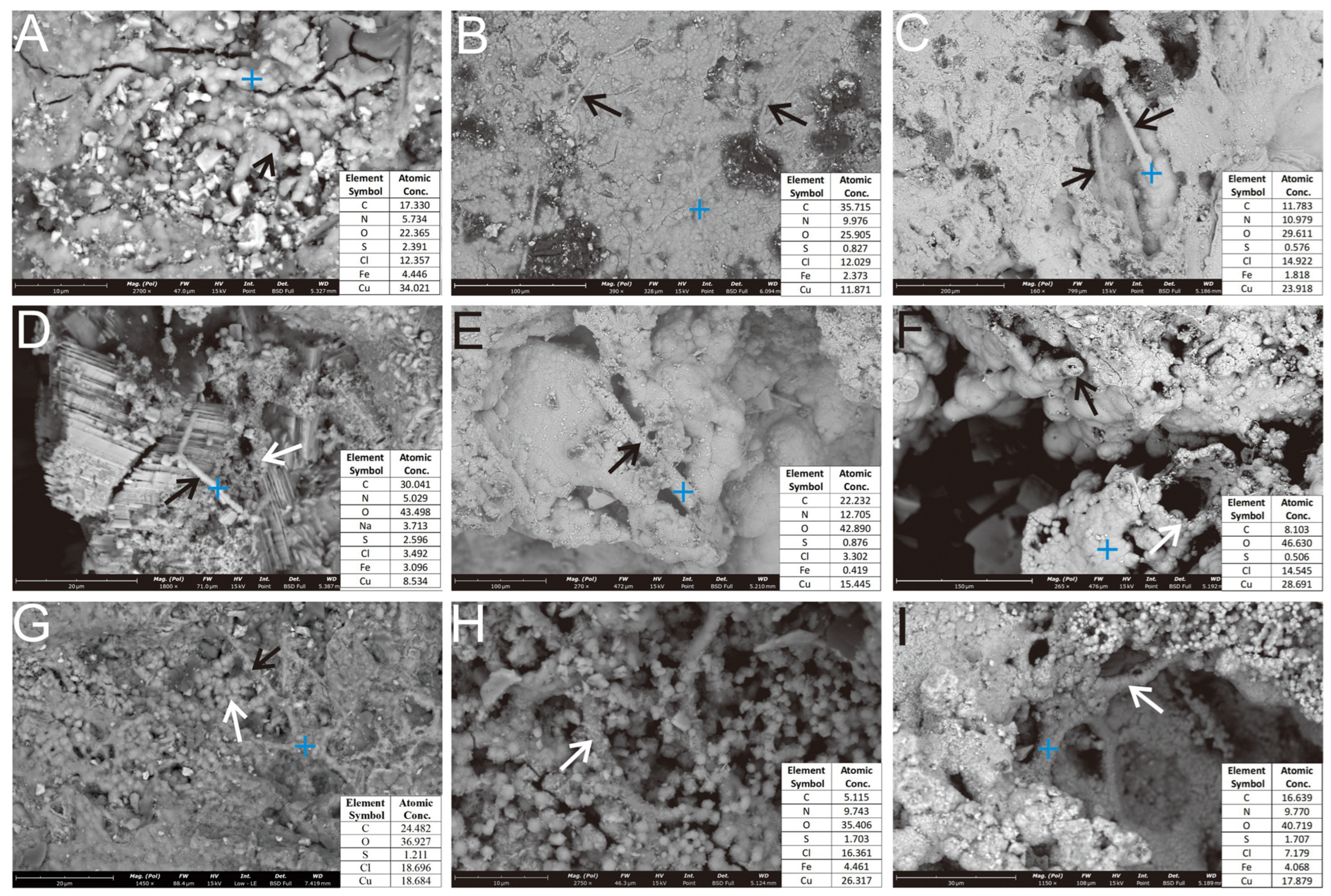
3.2.1. Oxides Formed During the Incubation Experiment
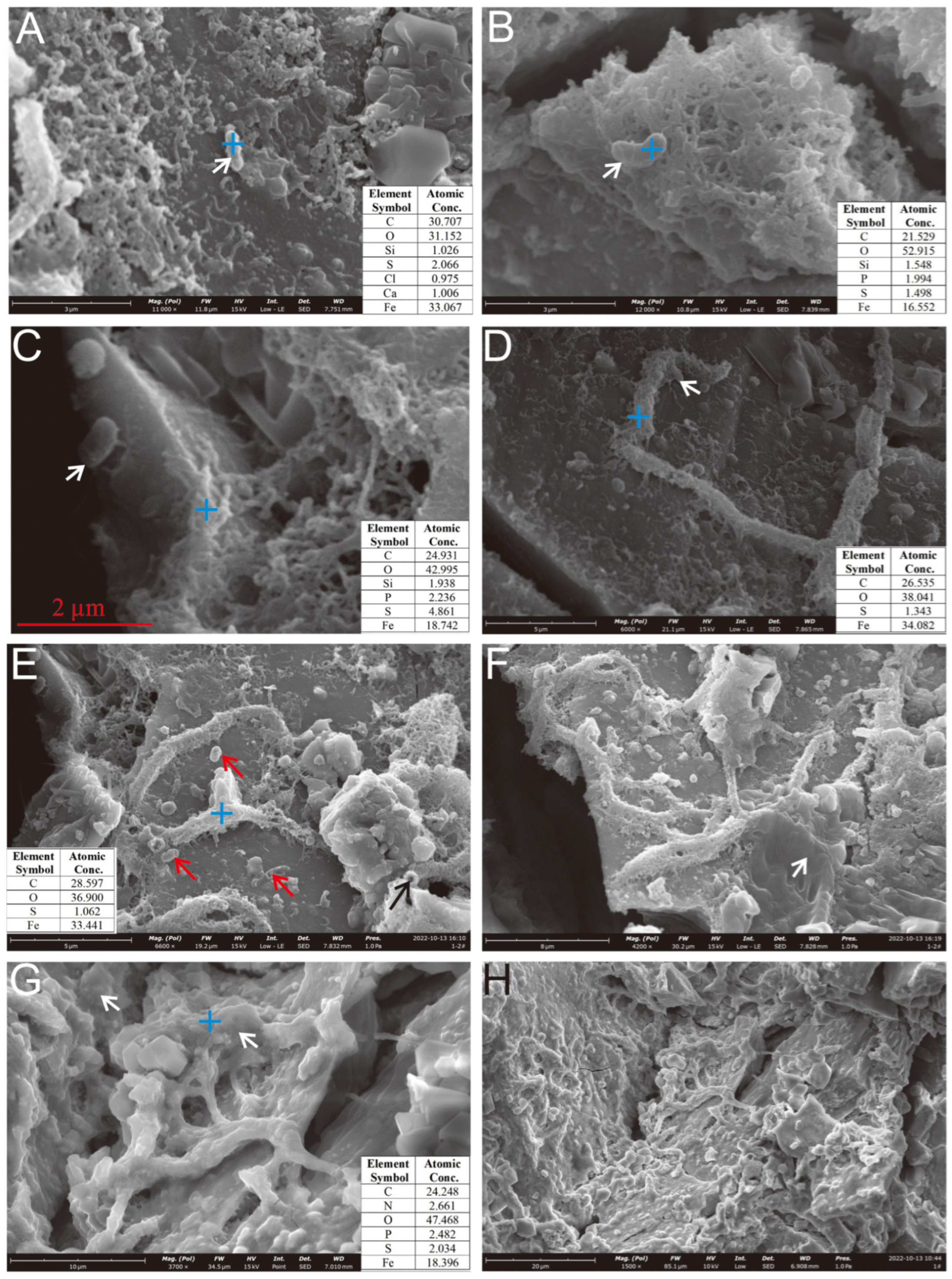
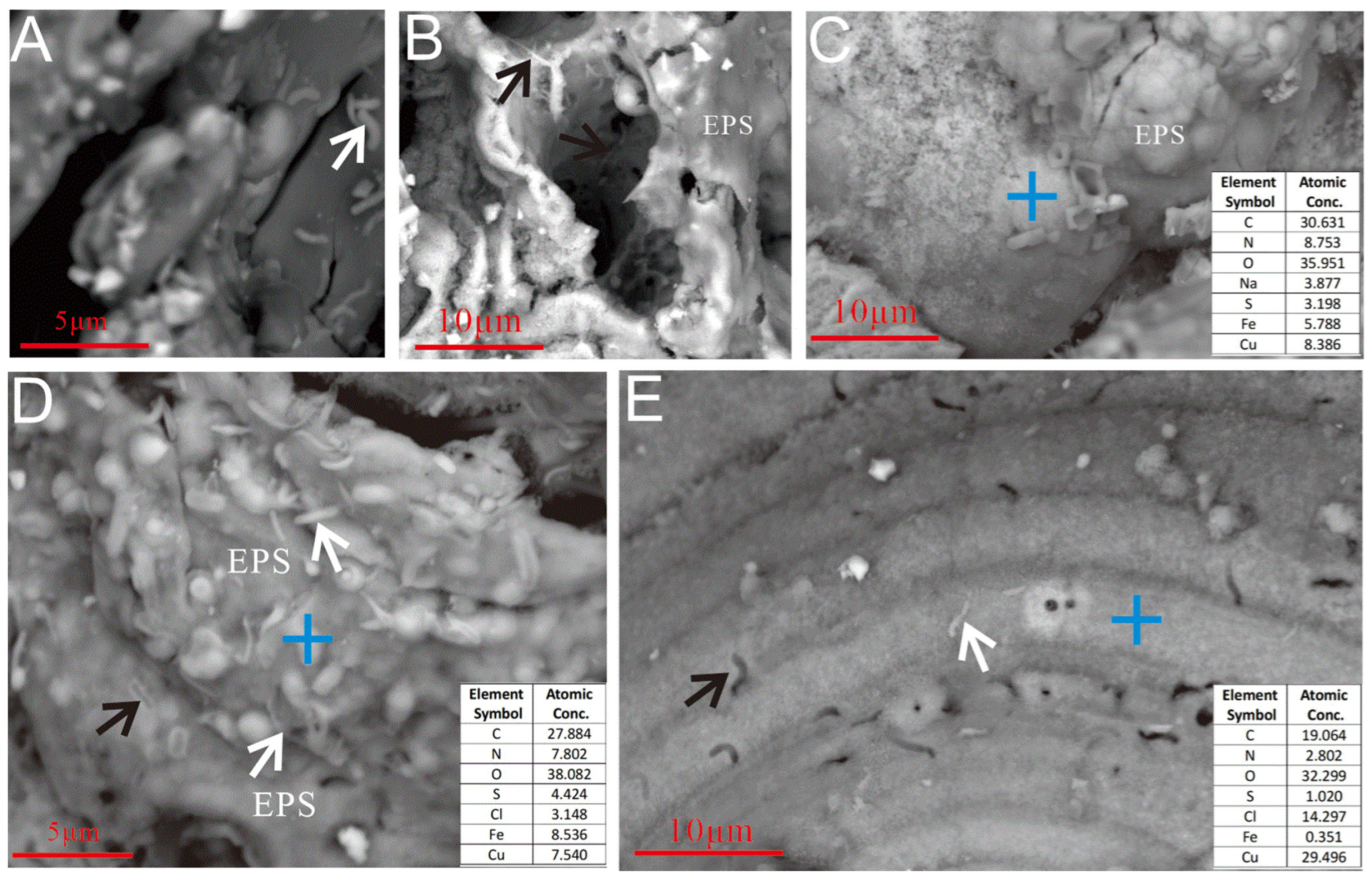
3.2.2. Microbial Mineralization Structures
3.2.3. Microbial Cells and EPSs
3.2.4. Cell Staining and Fluorescence Microscopy Observation
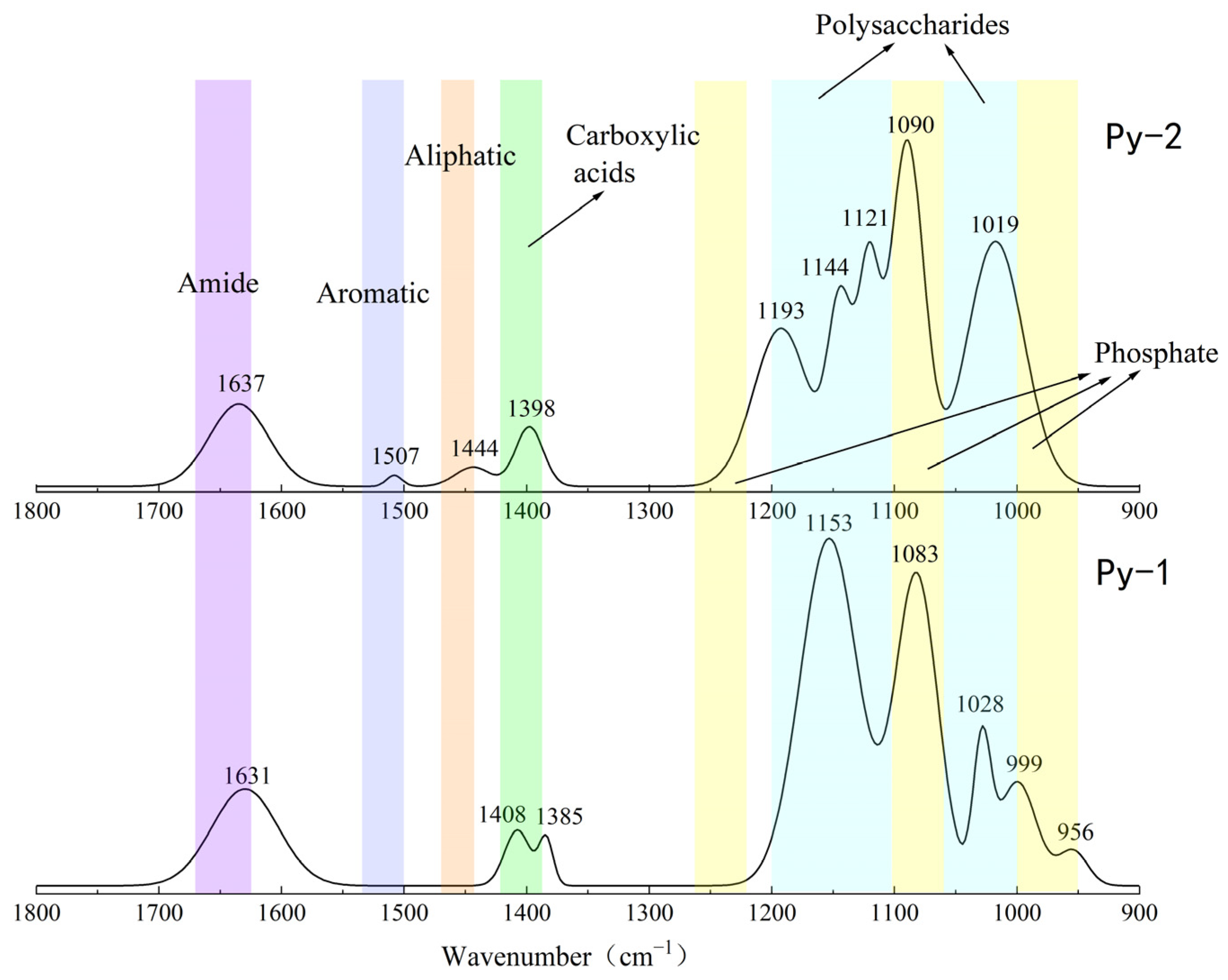
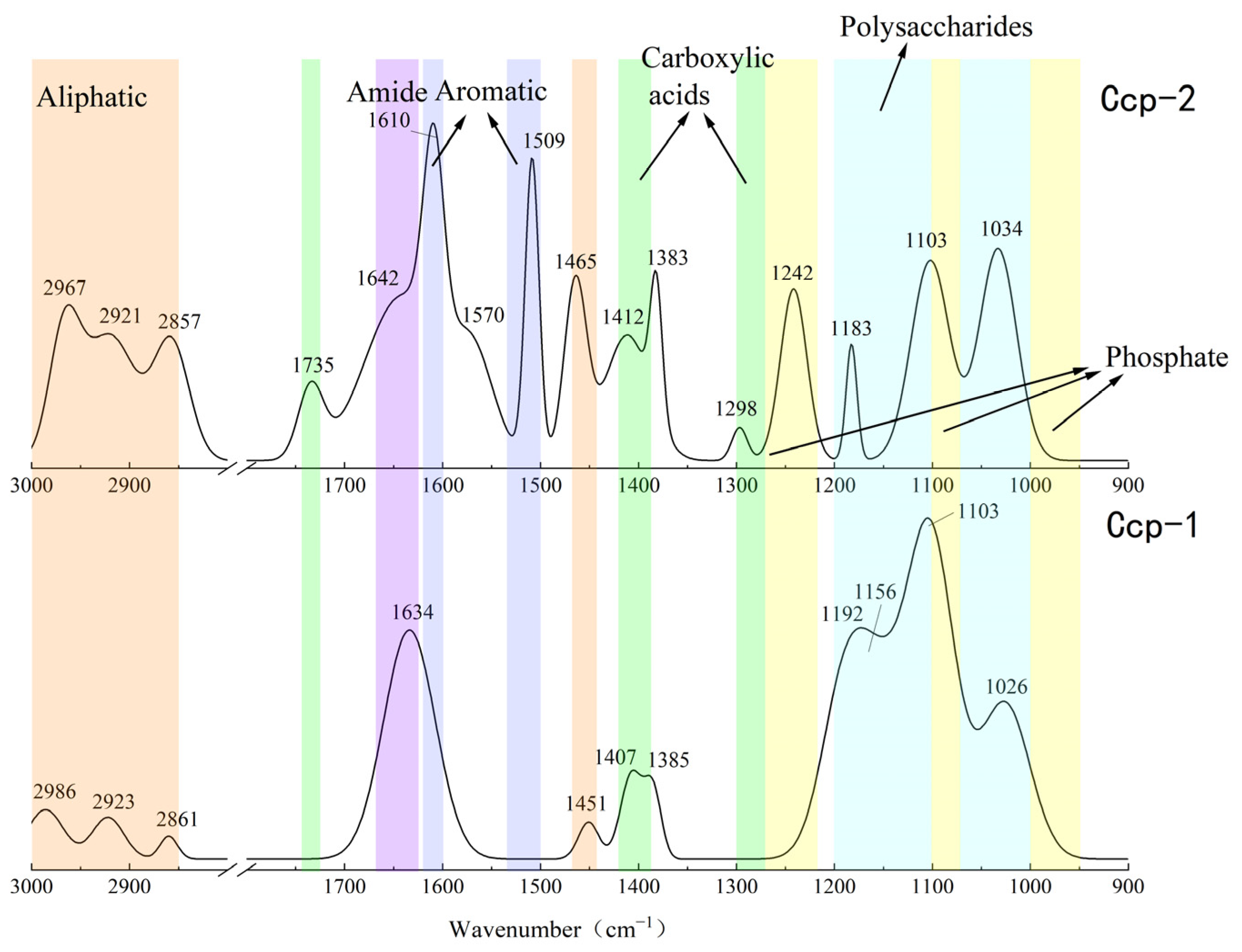
3.3. FT-IR Analysis of Organic Functional Groups
3.3.1. Organic Functional Groups in Sulfides Before and After Experimentation
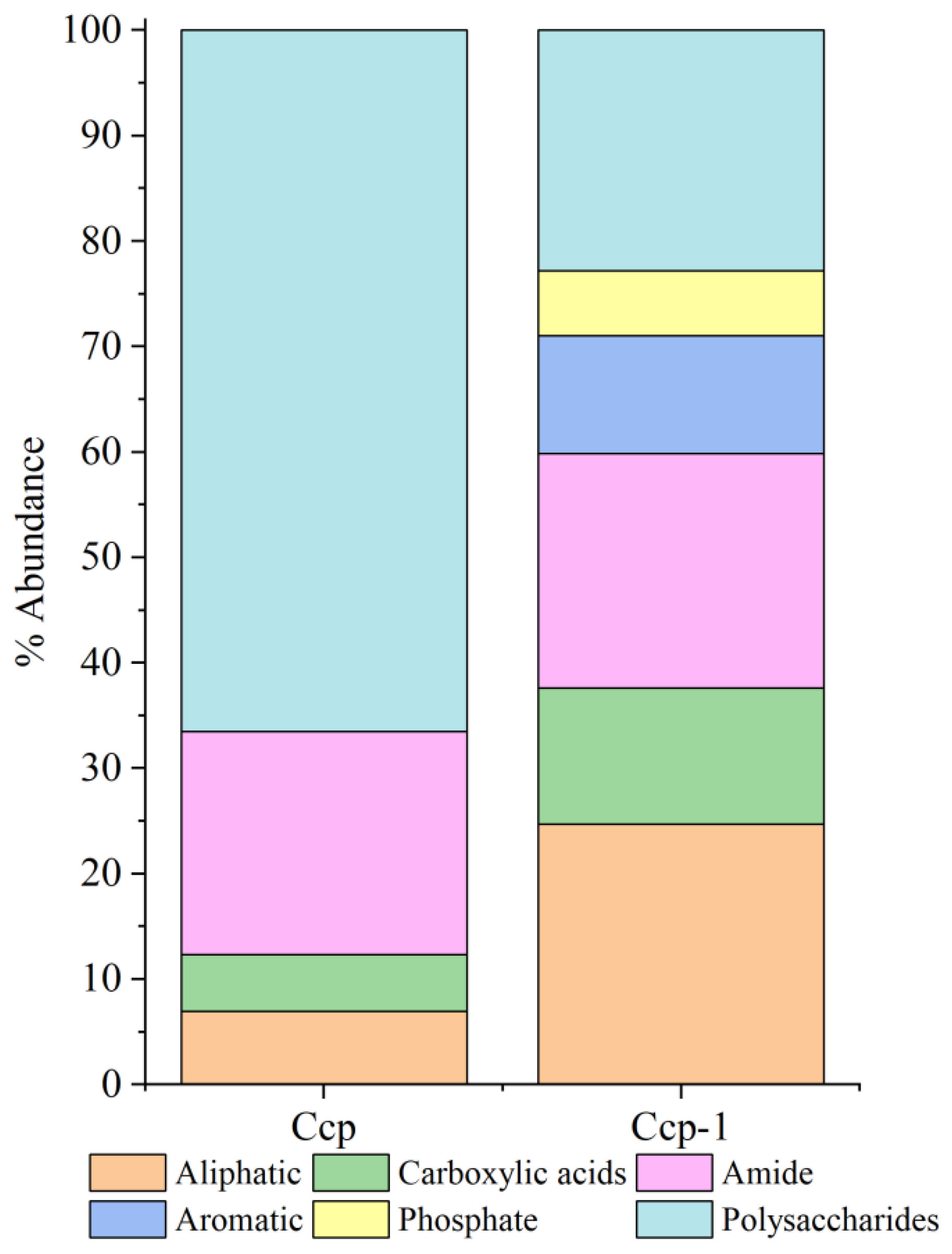
3.3.2. Organic Fractions in Sulfides Before and After In Situ Incubation
4. Discussion
4.1. Characterizing Microbial Interactions with Sulfide Minerals
4.1.1. Surface Evidence of Microbial Weathering
4.1.2. Biogenic Mineralization Structures in Microbial Weathering
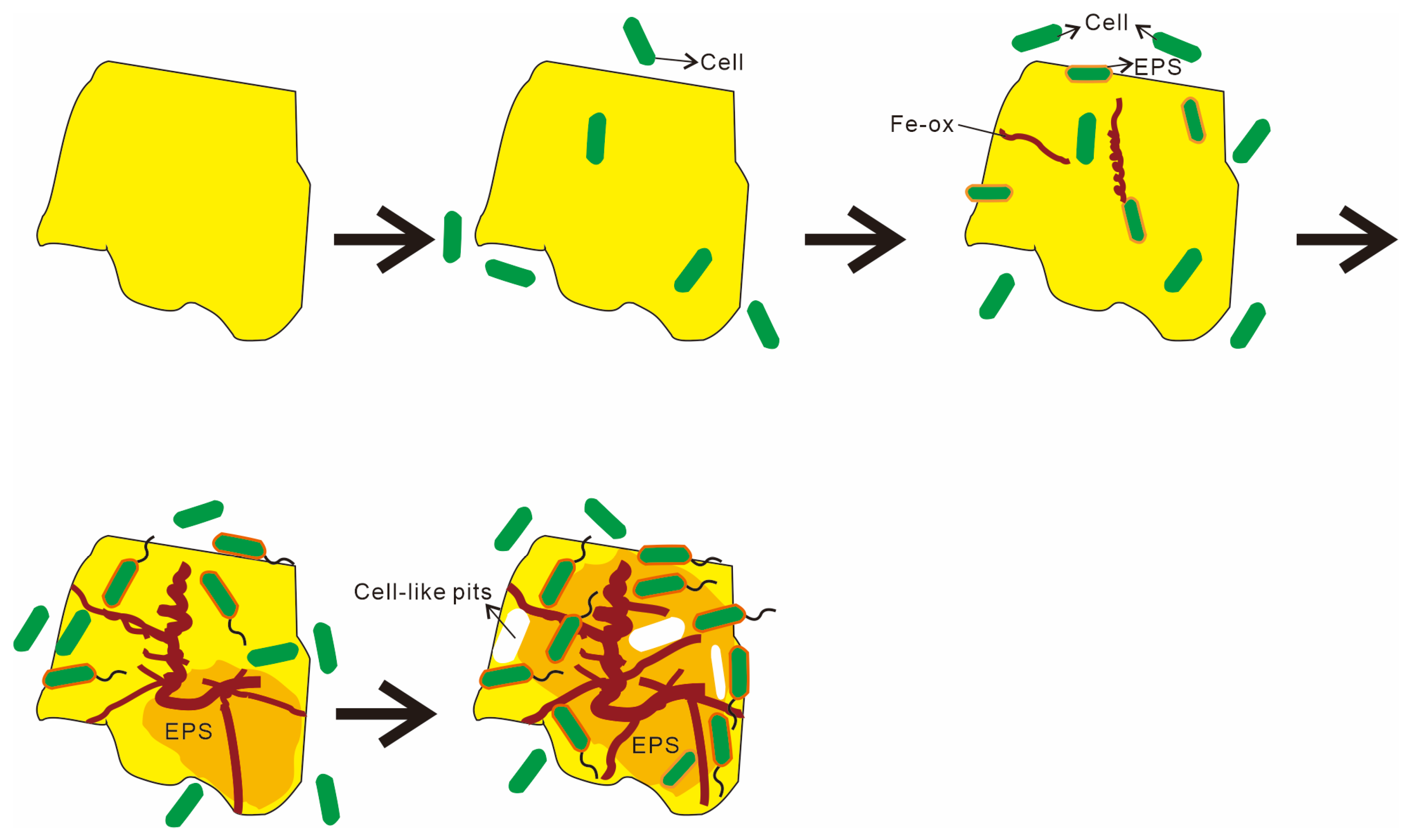
4.2. Stages and Mechanisms of Microbial Weathering in Hydrothermal Sulfides
5. Conclusions
- Microbial influence on seafloor sulfide weathering. Microorganisms play a crucial role in the weathering of seafloor sulfides. Both the Py and Ccp samples were influenced by the combined effects of chemical and microbial weathering.
- Differences in microbial weathering mechanisms. The microbial weathering mechanisms for the two sulfides differ significantly. The microbial weathering of Py is primarily mediated by an EPS-driven indirect contact mechanism, whereas Ccp weathering results from the interplay of both direct and indirect contact mechanisms.
- Distinct stages of microbe–mineral interaction. The process of microbe–sulfide mineral interaction can be summarized in four phases: (1) microbes approach the mineral surface via diffusion, sedimentation, and cell motility; (2) negatively charged bacteria adsorb onto positively charged mineral surfaces through electrostatic attraction; (3) microbial cells firmly adhere to the mineral surface, making detachment difficult; (4) extensive colonization of the mineral surface by microbial cells occurs, leading to biofilm formation and sustained weathering activity.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edmond, J.M.; Measures, C.I.; McDuff, R.E.; Chan, L.H.; Collier, R.; Grant, B.; Gordon, L.I.; Corliss, J.B. Ridge crest hydrothermal activity and the balances of the major and minor elements in the ocean: The Galapagos data. Earth Planet. Sci. Lett. 1979, 46, 1–18. [Google Scholar]
- Herzig, P.M.; Hannington, M.D. Polymetallic massive sulfides at the modern seafloor: A review. Ore Geol. Rev. 1995, 10, 95–115. [Google Scholar]
- Hannington, M.; Jamieson, J.; Monecke, T.; Petersen, S. Estimating the Metal Content of SMS Deposits 2011; IEEE: Piscataway, NJ, USA, 2011; pp. 1–4. [Google Scholar]
- Sylvan, J.B.; Pyenson, B.; Rouxel, O.J.; German, C.R.; Edwards, K.J. Time-series analysis of two hydrothermal plumes at 9°50′ N East Pacific Rise reveals distinct, heterogeneous bacterial populations. Geobiology 2012, 10, 249–267. [Google Scholar] [CrossRef]
- Rohwerder, T.; Gehrke, T.; Kinzler, K.; Sand, W. Bioleaching review part A: Progress in bioleaching: Fundamentals and mechanisms of bacterial metal sulfide oxidation. Appl. Microbiol. Biotechnol. 2003, 63, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Fallon, E.K.; Niehorster, E.; Brooker, R.A.; Scott, T.B. Oxidative dissolution of hydrothermal mixed-sulphide ore: An assessment of current knowledge in relation to seafloor massive sulphide mining. Ore Geol. Rev. 2017, 86, 309–337. [Google Scholar]
- Knight, R.D.; Roberts, S.; Cooper, M.J. Investigating monomineralic and polymineralic reactions during the oxidation of sulphide minerals in seawater: Implications for mining seafloor massive sulphide deposits. Appl. Geochem. 2018, 90, 63–74. [Google Scholar] [CrossRef]
- Edwards, K.J.; McCollom, T.M.; Konishi, H.; Buseck, P.R. Seafloor bioalteration of sulfide minerals: Results from in situ incubation studies. Geochim. Cosmochim. Acta 2003, 67, 2843–2856. [Google Scholar] [CrossRef]
- Krepski, S.T.; Emerson, D.; Hredzak-Showalter, P.L.; Luther III, G.W.; Chan, C.S. Morphology of biogenic iron oxides records microbial physiology and environmental conditions: Toward interpreting iron microfossils. Geobiology 2013, 11, 457–471. [Google Scholar] [CrossRef]
- Fleming, E.J.; McAllister, S.M.; Davis, R.E.; Chan, C.S.; Moyer, C.L.; Tebo, B.M.; Emerson, D. Hidden in plain sight: Discovery of sheath-forming, iron-oxidizing Zetaproteobacteria at Loihi Seamount, Hawaii, USA. FEMS Microbiol. Ecol. 2013, 85, 116–127. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, X.; Wang, W. Microbial Imprints on Sulfide Minerals in Submarine Hydrothermal Deposits of the East Pacific Rise. J. Geophys. Res. Biogeosci. 2020, 125, 8. [Google Scholar] [CrossRef]
- Liu, H.; Lu, X.; Zhang, L.; Xiang, W.; Zhu, X.; Li, J.; Wang, X.; Lu, J.; Wang, R. Collaborative effects of Acidithiobacillus ferrooxidans and ferrous ions on the oxidation of chalcopyrite. Chem. Geol. 2018, 493, 109–120. [Google Scholar] [CrossRef]
- Kocaman, A.T.; Cemek, M.; Edwards, K.J. Kinetics of pyrite, pyrrhotite, and chalcopyrite dissolution by Acidithiobacillus ferrooxidans. Can. J. Microbiol. 2016, 62, 629–642. [Google Scholar] [CrossRef]
- Dong, H.; Huang, L.; Zhao, L.; Zeng, Q.; Liu, X.; Sheng, Y.; Shi, L.; Wu, G.; Jiang, H.; Li, F.; et al. A critical review of mineral-microbe interaction and co-evolution: Mechanisms and applications. Natl. Sci. Rev. 2022, 9, nwac128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, W.; Cai, Y.; Qu, Y.; Pan, Y.; Zhang, W.; Zhao, K. Experimental investigation of the reactions between pyrite and aqueous Cu(I) chloride solution at 100–250 °C. Geochim. Cosmochim. Acta 2021, 298, 1–20. [Google Scholar] [CrossRef]
- Léveillé, R.J.; Juniper, S.K. Microbial colonization and weathering of sulphide minerals at deep-sea hydrothermal vents: In situ exposure experiments. Cah. Biol. Mar. 2002, 43, 285–288. [Google Scholar]
- Barco, R.A.; Hoffman, C.L.; Ramírez, G.A.; Toner, B.M.; Edwards, K.J.; Sylvan, J.B. In situ incubation of iron-sulfur mineral reveals a diverse chemolithoautotrophic community and a new biogeochemical role for Thiomicrospira. Environ. Microbiol. 2017, 19, 1322–1337. [Google Scholar] [CrossRef]
- Emerson, D.; Scott, J.J.; Leavitt, A.; Fleming, E.J.; Moyer, C.L. In situ estimates of iron-oxidation and accretion rates for iron-oxidizing bacterial mats at Loihi Seamount. Deep-Sea Res. I Oceanogr. Res. Pap. 2017, 126, 31–39. [Google Scholar] [CrossRef]
- Wang, C.H.; Gulmann, L.K.; Zhang, T.; Farfán, G.A.; Hansel, C.M.; Sievert, S.M. Microbial colonization of metal sulfide minerals at a diffuse-flow deep-sea hydrothermal vent at 9°50′ N on the East Pacific Rise. Geobiology 2020, 18, 594–605. [Google Scholar] [CrossRef]
- Mitsunobu, S.; Ohashi, Y.; Makita, H.; Suzuki, Y.; Nozaki, T.; Ohigashi, T.; Ina, T.; Takaki, Y. One-year incubation of pyrite at the deep seafloor and its microbiological and biogeochemical characterizations. Appl. Environ. Microbiol. 2021, 87, e00977-21. [Google Scholar] [CrossRef]
- Wang, Y.; Han, X.; Petersen, S.; Frische, M.; Qiu, Z.; Li, H.; Wu, Z.; Cui, R. Mineralogy and trace element geochemistry of sulfide minerals from the Wocan Hydrothermal Field on the slow-spreading Carlsberg Ridge, Indian Ocean. Ore Geol. Rev. 2017, 84, 1–19. [Google Scholar]
- Cai, Y.; Han, X.; Petersen, S.; Wang, Y.; Qiu, Z.; Yang, M. Mineralization of sulfide mound at Wocan-1 hydrothermal field based on geological mapping, mineralogy and sulfur isotope studies, Carlsberg Ridge, Northwest Indian Ocean. Ore Geol. Rev. 2024, 171, 106180. [Google Scholar] [CrossRef]
- Jesús, J.J.; Dittrich, M. Fourier transform infrared spectroscopy for molecular analysis of microbial cells. Methods Mol. Biol. 2012, 881, 187–211. [Google Scholar]
- Movasaghi, Z.; Rehman, S.; ur Rehman, D.I. Fourier transform infrared (FTIR) spectroscopy of biological tissues. Appl. Spectrosc. Rev. 2008, 43, 134–179. [Google Scholar] [CrossRef]
- Chan, C.S.; Fakra, S.C.; Emerson, D.; Fleming, E.J.; Edwards, K.J. Lithotrophic iron-oxidizing bacteria produce organic stalks to control mineral growth: Implications for biosignature formation. ISME J. 2011, 5, 717–727. [Google Scholar] [CrossRef]
- Miot, J.; Benzerara, K.; Obst, M.; Kappler, A.; Hegler, F.; Schädler, S.; Bouchez, C.; Guyot, F.; Morin, G. Extracellular iron biomineralization by photoautotrophic iron-oxidizing bacteria. Appl. Environ. Microbiol. 2009, 75, 5586–5591. [Google Scholar] [CrossRef] [PubMed]
- Emerson, D.; Moyer, C.L. Biogeochemistry and microbiology of microaerobic Fe(II) oxidation. Biochem. Soc. Trans. 2012, 40, 1211–1216. [Google Scholar] [CrossRef]
- Emerson, D.; Moyer, C.L. Neutrophilic Fe-oxidizing bacteria are abundant at the Loihi Seamount hydrothermal vents and play a major role in Fe oxide deposition. Appl. Environ. Microbiol. 2002, 68, 3085–3093. [Google Scholar] [CrossRef]
- Li, J.T.; Zhou, H.; Peng, X.; Wu, Z.; Chen, S.; Fang, J. Microbial diversity and biomineralization in low-temperature hydrothermal iron-silica precipitates of the Lau Basin hydrothermal field. FEMS Microbiol. Ecol. 2012, 81, 205–216. [Google Scholar] [CrossRef]
- Chan, C.S.; McAllister, S.M.; Leavitt, A.H.; Glazer, B.T.; Krepski, S.T.; Emerson, D. The architecture of iron microbial mats reflects the adaptation of chemolithotrophic iron oxidation in freshwater and marine environments. Front. Microbiol. 2016, 7, 796. [Google Scholar] [CrossRef]
- Peng, X.; Zhou, H.; Yao, H.; Li, J.; Tang, S.; Jiang, L.; Wu, Z. Microbe-related precipitation of iron and silica in the Edmond deep-sea hydrothermal vent field on the Central Indian Ridge. Science Bull. 2007, 52, 3233–3238. [Google Scholar] [CrossRef]
- Li, J.; Su, L.; Wang, F.; Yang, J.; Gu, L.; Sun, M.; Li, Q.; Zhou, H.; Fang, J. Elucidating the biomineralization of low-temperature hydrothermal precipitates with varying Fe, Si contents: Indication from ultrastructure and microbiological analyses. Deep Sea Res. Part I Oceanogr. Res. Pap. 2020, 157, 103208. [Google Scholar] [CrossRef]
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, B. Filamentous microfossils in a 3,235-million-year-old volcanogenic massive sulphide deposit. Nature 2000, 405, 676–679. [Google Scholar] [CrossRef]
- Ferris, F.G.; Fyfe, W.S.; Beveridge, T.J. Metallic ion binding by Bacillus subtilis: Implications for the fossilization of microorganisms. Geology 1988, 16, 149–152. [Google Scholar] [CrossRef]
- Flemming, H.C.; Neu, T.R.; Wozniak, D.J. The EPS matrix: The “house of biofilm cells”. J. Bacteriol. 2007, 189, 7945–7947. [Google Scholar] [CrossRef] [PubMed]
- Vrdoljak, G.A.; Spiller, S. Characterization of an acid mine biofilm by cryo scanning electron microscopy. Microsc. Microanal. 2005, 11, 292–293. [Google Scholar] [CrossRef]
- Egli, M.; Mavris, C.; Mirabella, A.; Giaccai, D. Soil organic matter formation along a chronosequence in the Morteratsch proglacial area (Upper Engadine, Switzerland). Catena 2010, 82, 61–69. [Google Scholar]
- Zeng, W.; Li, F.; Wu, C.; Yu, R.; Wu, X.; Shen, L.; Liu, Y.; Qiu, G.; Li, J. Role of extracellular polymeric substance (EPS) in toxicity response of soil bacteria Bacillus sp. S3 to multiple heavy metals. Bioprocess Biosyst. Eng. 2020, 43, 153–167. [Google Scholar] [CrossRef]
- Niemeyer, J.; Chen, Y.; Bollag, J.M. Characterization of humic acids, composts, and peat by diffuse reflectance Fourier-transform infrared spectroscopy. Soil Sci. Soc. Am. J. 1992, 56, 135–140. [Google Scholar] [CrossRef]
- Cocozza, C.; D’Orazio, V.; Miano, T.M.; Shotyk, W. Characterization of solid and aqueous phases of a peat bog profile using molecular fluorescence spectroscopy, ESR, and FT-IR, and comparison with physical properties. Org. Geochem. 2003, 34, 49–60. [Google Scholar] [CrossRef]
- Artz, R.R.E.; Chapman, S.J.; Campbell, C.D. Substrate utilisation profiles of microbial communities in peat are depth dependent and correlate with whole soil FTIR profiles. Soil Biol. Biochem. 2006, 38, 2958–2962. [Google Scholar] [CrossRef]
- Jesús, J.J.; Romero-González, M.E.; Banwart, S.A. Analysis of bacteria on steel surfaces using reflectance micro-Fourier transform infrared spectroscopy. Anal. Chem. 2009, 81, 6467–6473. [Google Scholar]
- Omoike, A.; Chorover, J. Adhesion of bacterial exopolymers to alpha-FeOOH: Inner-sphere complexation of phosphodiester groups. Langmuir 2004, 20, 11108–11114. [Google Scholar] [CrossRef] [PubMed]
- Cagnasso, M.; Boero, V.; Franchini, M.A.; Chorover, J. ATR-FTIR studies of phospholipid vesicle interactions with α-FeOOH and α-Fe2O3 surfaces. Colloids Surf. B Biointerfaces 2010, 76, 456–467. [Google Scholar] [CrossRef]
- Omoike, A.; Chorover, J. Adsorption to goethite of extracellular polymeric substances from Bacillus subtilis. Geochim. Cosmochim. Acta 2006, 70, 827–838. [Google Scholar] [CrossRef]
- Laufer, K.; Nordhoff, M.; Halama, M.; Martinez, R.E.; Obst, M.; Nowak, M.; Stryhanyuk, H.; Richnow, H.H.; Kappler, A. Microaerophilic Fe(II)-oxidizing Zetaproteobacteria isolated from low-Fe marine coastal sediments: Physiology and composition of their twisted stalks. Appl. Environ. Microbiol. 2017, 83, e03118-16. [Google Scholar] [CrossRef]
- Ueshima, M.; Ginn, B.R.; Haack, E.A.; Szymanowski, J.E.; Fein, J.B. Cd adsorption onto Pseudomonas putida in the presence and absence of extracellular polymeric substances. Geochim. Cosmochim. Acta 2008, 72, 5885–5895. [Google Scholar] [CrossRef]
- Demyan, M.S.; Rasche, F.; Schulz, E.; Breulmann, M.; Müller, T.; Cadisch, G. Use of specific peaks obtained by diffuse reflectance Fourier transform mid-infrared spectroscopy to study the composition of organic matter in a Haplic Chernozem. Eur. J. Soil Sci. 2012, 63, 189–199. [Google Scholar] [CrossRef]
- Verati, C.; de Donato, P.; Prieur, D.; Lancelot, J. Evidence of bacterial activity from micrometer-scale layer analyses of black-smoker sulfide structures (Pito Seamount site, Easter microplate). Chem. Geol. 1999, 158, 257–269. [Google Scholar] [CrossRef]
- Asta, M.P.; Cama, J.; Soler, J.M.; Arvidson, R.S.; Lüttge, A. Interferometric study of pyrite surface reactivity in acidic conditions. Am. Miner. 2008, 93, 508–519. [Google Scholar] [CrossRef]
- Lefticariu, L.; Pratt, L.A.; Laverne, J.A.; Schimmelmann, A. Anoxic pyrite oxidation by water radiolysis products: A potential source of biosustaining energy. Earth Planet. Sci. Lett. 2010, 292, 57–67. [Google Scholar] [CrossRef]
- Zhu, T.; Lu, X.; Liu, H.; Li, J.; Zhu, X.; Lu, J.; Wang, R. Quantitative X-ray photoelectron spectroscopy-based depth profiling of bioleached arsenopyrite surface by Acidithiobacillus ferrooxidans. Geochim. Cosmochim. Acta 2014, 127, 120–139. [Google Scholar] [CrossRef]
- Li, J.; Lu, J.; Lu, X.; Tu, B.; Wang, R. Sulfur Transformation in Microbially Mediated Pyrite Oxidation by Acidithiobacillus ferrooxidans: Insights from X-ray Photoelectron Spectroscopy-Based Quantitative Depth Profiling. Geomicrobiol. J. 2016, 33, 118–134. [Google Scholar] [CrossRef]
- Wei, M.; Zeng, X.; Han, X.; Shao, Z.; Xie, Q.; Dong, C.; Wang, Y.; Qiu, Z. Potential autotrophic carbon-fixer and Fe(II)-oxidizer Alcanivorax sp. MM125-6 isolated from Wocan hydrothermal field. Front. Microbiol. 2022, 13, 1–12. [Google Scholar] [CrossRef]
- Edwards, K.J.; Bach, W.; McCollom, T.M.; Rogers, D.R. Neutrophilic iron-oxidizing bacteria in the ocean: Their habitats, diversity, and roles in mineral deposition, rock alteration, and biomass production in the deep sea. Geomicrobiol. J. 2004, 21, 393–404. [Google Scholar] [CrossRef]
- Peng, X.T.; Ta, K.W.; Chen, S.; Zhang, L.J.; Xu, H.C. Coexistence of Fe(II)- and Mn(II)-oxidizing bacteria govern the formation of deep-sea umber deposits. Geochim. Cosmochim. Acta 2015, 169, 200–216. [Google Scholar] [CrossRef]
- Kennedy, C.B.; Scott, S.D.; Ferris, F.G. Ultrastructure and potential sub-seafloor evidence of bacteriogenic iron oxides from Axial Volcano, Juan de Fuca Ridge, northeast Pacific Ocean. FEMS Microbiol. Ecol. 2003, 43, 247–254. [Google Scholar]
- Cornell, R.M.; Schwertmann, U. The Iron Oxides: Structure, Properties, Reactions, Occurrences and Uses, 2nd ed.; Wiley-VCH: Hoboken, NJ, USA, 2004. [Google Scholar]
- Jambor, J.L.; Dutrizac, J.E. Occurrence and constitution of natural and synthetic ferrihydrite, a widespread iron oxyhydroxide. Chem. Rev. 1998, 98, 2549–2586. [Google Scholar] [CrossRef]
- Davis, R.E.; Moyer, C.L. Extreme spatial and temporal variability of hydrothermal microbial mat communities along the Mariana Island Arc and southern Mariana back-arc system. J. Geophys. Res. Solid Earth 2008, 113. [Google Scholar] [CrossRef]
- Davis, R.E.; Stakes, D.S.; Wheat, C.G.; Moyer, C.L. Bacterial variability within an iron-silica-manganese-dominated hydrothermal mound located off-axis at the Cleft Segment, Juan de Fuca Ridge. Geomicrobiol. J. 2009, 26, 570–580. [Google Scholar] [CrossRef]
- Edwards, K.J.; Glazer, B.T.; Rouxel, O.J.; Bach, W.; Emerson, D.; Davis, R.E. Ultra-diffuse hydrothermal venting supports Fe-oxidizing bacteria and massive umber deposition at 5000 m off Hawaii. ISME J. 2011, 5, 1748–1758. [Google Scholar] [CrossRef]
- McAllister, S.M.; Moore, R.M.; Gartman, A.; Luther, G.W.; Emerson, D.; Chan, C.S. The Fe(II)-oxidizing Zetaproteobacteria: Historical, ecological and genomic perspectives. FEMS Microbiol. Ecol. 2019, 95, fiz015. [Google Scholar] [CrossRef] [PubMed]
- Little, C.T.S.; Johannessen, K.C.; Bengtson, S.; Chan, C.S.; Ivarsson, M.; Slack, J.F.; Broman, C.; Thorseth, I.H.; Grenne, T.; Rouxel, O.J.; et al. A late Paleoproterozoic (1.74 Ga) deep-sea, low-temperature, iron-oxidizing microbial hydrothermal vent community from Arizona, USA. Geobiology 2021, 19, 228–249. [Google Scholar] [CrossRef]
- Aguirre, P.; Guerrero, K.; Sánchez-Rodríguez, A.; Gentina, J.C.; Schippers, A. Making sticky cells: Effect of galactose and ferrous iron on the attachment of Leptospirillum ferrooxidans to mineral surfaces. Res. Microbiol. 2018, 169, 569–575. [Google Scholar] [CrossRef]
- Edwards, K.J.; Rogers, D.R.; Wirsen, C.O.; McCollom, T.M. Isolation and characterization of novel iron-oxidizing bacteria that grow at circumneutral pH. Appl. Environ. Microbiol. 2003, 69, 4906–4913. [Google Scholar]
- Johnson, D.B.; Hallberg, K.B. The microbiology of acidic mine waters. Res. Microbiol. 2003, 154, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Fowler, T.A.; Holmes, P.R.; Crundwell, F.K. Mechanism of pyrite dissolution in the presence of Thiobacillus ferrooxidans. Appl. Environ. Microbiol. 1999, 65, 2987–2993. [Google Scholar] [CrossRef]
- Dasgupta, S.; Peng, X.; Tivey, M.K. Interaction between microbes, minerals, and fluids in deep-sea hydrothermal systems. Minerals 2021, 11, 1324. [Google Scholar] [CrossRef]
- Rodriguez-Leiva, M.; Tributsch, H. Morphology of bacterial leaching patterns by Thiobacillus ferrooxidans on synthetic pyrite. Arch. Microbiol. 1988, 149, 401–405. [Google Scholar] [CrossRef]
- Moon, H.S.; Kim, B.A.; Hyun, S.P.; Lee, Y.H.; Shin, D. Effect of the redox dynamics on microbial-mediated As transformation coupled with Fe and S in flow-through sediment columns. J. Hazard. Mater. 2017, 329, 280–289. [Google Scholar] [CrossRef]
- Jones, R.A.; Koval, S.F.; Nesbitt, H.W. Surface alteration of arsenopyrite (FeAsS) by Thiobacillus ferrooxidans. Geochim. Cosmochim. Acta 2003, 67, 955–965. [Google Scholar] [CrossRef]
- Lilova, K.; Karamanev, D. Direct oxidation of copper sulfide by a biofilm of Acidithiobacillus ferrooxidans. Hydrometallurgy 2005, 80, 147–154. [Google Scholar] [CrossRef]
- Crundwell, F.K. How do bacteria interact with minerals? Hydrometallurgy 2003, 71, 75–81. [Google Scholar] [CrossRef]
- Rojas-Chapana, J.A.; Tributsch, H. Bio-leaching of pyrite accelerated by cysteine. Process Biochem. 2000, 35, 815–824. [Google Scholar] [CrossRef]
- Lu, J.; Lu, X.; Wang, R.; Li, J.; Zhu, C. Pyrite surface after Thiobacillus ferrooxidans leaching at 30 °C. Acta Geol. Sin. 2006, 80, 362–367. [Google Scholar]
- Lu, X.; Wang, H. Microbial oxidation of sulfide tailings and the environmental consequences. Elements 2012, 8, 119–124. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, R.; Lu, X.; Lu, J.; Li, C.; Li, J. Bioleaching of chalcopyrite by Acidithiobacillus ferrooxidans. Miner. Eng. 2013, 53, 184–192. [Google Scholar] [CrossRef]
- Wacey, D.; Kilburn, M.R.; Saunders, M.; Cliff, J.; Brasier, M.D. Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nat. Geosci. 2011, 4, 698–702. [Google Scholar]
- Capriel, P. Hydrophobicity of organic matter in arable soils: Influence of management. Eur. J. Soil Sci. 1997, 48, 457–462. [Google Scholar]
- Solari, J.A.; Huerta, G.; Escobar, B.; Valenzuela, T. Interfacial phenomena affecting the adhesion of Thiobacillus ferrooxidans to sulphide mineral surface. Colloids Surf. 1992, 68, 159–166. [Google Scholar] [CrossRef]
- Acuña, J.; Rojas, J.; Amaro, A.M.; Toledo, H.; Jerez, C.A. Chemotaxis of Leptospirillum ferrooxidans and other acidophilic chemolithotrophs: Comparison with the Escherichia coli chemosensory system. FEMS Microbiol. Lett. 1992, 75, 37–42. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Sample Description |
|---|---|
| Py-dominated | Pyrite-dominated, with a porous texture |
| Ccp-dominated | Chalcopyrite-dominated, minor pyrite and sphalerite, well crystallized |
| Wave Numbers (cm)−1 | Vibration Types | Functional Groups | Reference |
|---|---|---|---|
| 2850~3000. 1465, 1451, 1444 | C-H stretching vibrations (νC-H), corresponding to CH3 and CH2; | Aliphatic compounds | [23,38] |
| 1735, 1412, 1408, 1407, 1400, 1398, 1385, 1383, 1298 | Vibrational C=O Stretching (νC=O) | Carboxylic acids | [23,39] |
| 1660~1630 | Stretching C=C; Stretching C=O | Amides | [23,24] |
| 1610, 1509, 1507 | C=C; | Aromatic compounds | [38,40,41,42] |
| 1270–1220, 1100–1070, 1000–950 | ν(P=O); ν(PO)2− | Phosphate | [23,38,43] |
| 1200~950 | Vibrations of C-OH, C-O-C, and C-C | Polysaccharides | [23,43] |
| Sulfide | Treatment | Aliphatic Compound | Carboxylic Acids | Amides | Aromatic Compound | Phosphate | Polysaccharide |
|---|---|---|---|---|---|---|---|
| Py | pre-incubation | 0 | 4.25% | 13.1% | 0 | 36.82% | 45.92% |
| post-incubation | 1.37% | 3.28% | 9.66% | 0.34% | 23.11% | 62.23% | |
| Ccp | pre-incubation | 6.96% | 5.36% | 21.16% | 0 | 0 | 66.52% |
| post-incubation | 24.71% | 12.92% | 22.21% | 11.21% | 6.12% | 22.83% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Han, X.; Wang, Y.; Liu, J.; Wei, M. Microbial Seafloor Weathering of Hydrothermal Sulfides: Insights from an 18-Month In Situ Incubation at the Wocan-1 Hydrothermal Field. Biology 2025, 14, 389. https://doi.org/10.3390/biology14040389
Dong C, Han X, Wang Y, Liu J, Wei M. Microbial Seafloor Weathering of Hydrothermal Sulfides: Insights from an 18-Month In Situ Incubation at the Wocan-1 Hydrothermal Field. Biology. 2025; 14(4):389. https://doi.org/10.3390/biology14040389
Chicago/Turabian StyleDong, Chuanqi, Xiqiu Han, Yejian Wang, Jiqiang Liu, and Mingcong Wei. 2025. "Microbial Seafloor Weathering of Hydrothermal Sulfides: Insights from an 18-Month In Situ Incubation at the Wocan-1 Hydrothermal Field" Biology 14, no. 4: 389. https://doi.org/10.3390/biology14040389
APA StyleDong, C., Han, X., Wang, Y., Liu, J., & Wei, M. (2025). Microbial Seafloor Weathering of Hydrothermal Sulfides: Insights from an 18-Month In Situ Incubation at the Wocan-1 Hydrothermal Field. Biology, 14(4), 389. https://doi.org/10.3390/biology14040389