Nematicidal Effect and Histological Modifications Induced by Hydrolysable Tannin Extract on the Third-Stage Infective Larvae of Haemonchus contortus

 , , and
, , and
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethic Statements
2.2. Parasites
2.3. Hydrolysable Tannin Extract (HTE)
2.4. Bioassays
2.5. Scanning Electronic Microscopy (SEM)
2.6. Statistical Analyses
3. Results
3.1. HTE Induced Mortality on L3 Larvae of H. Contortus
3.2. Lethal Concentration of HTE on L3 of H. Contortus
3.3. Damage of L3 Induced by HTE Observed by Optical Microscopy
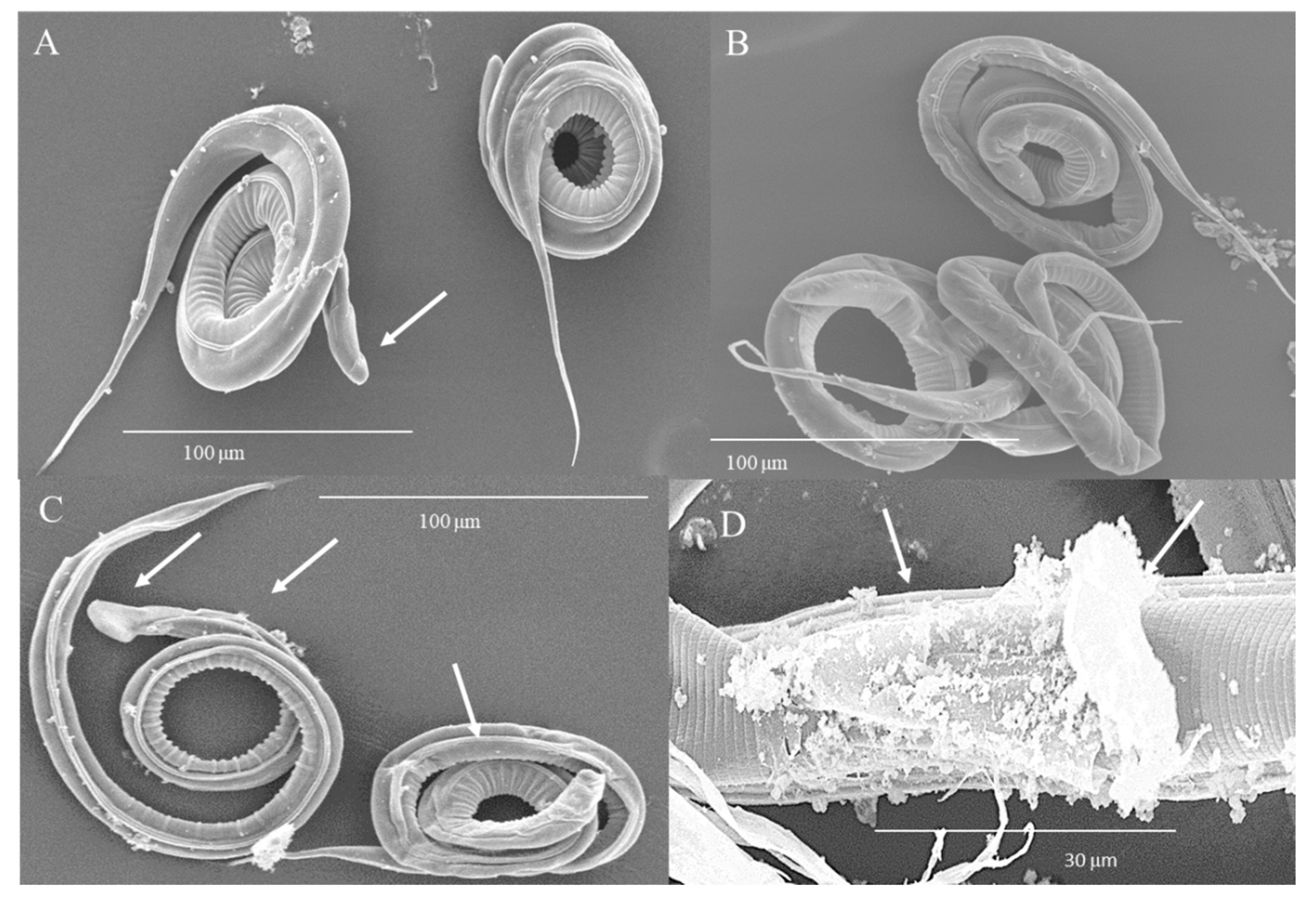
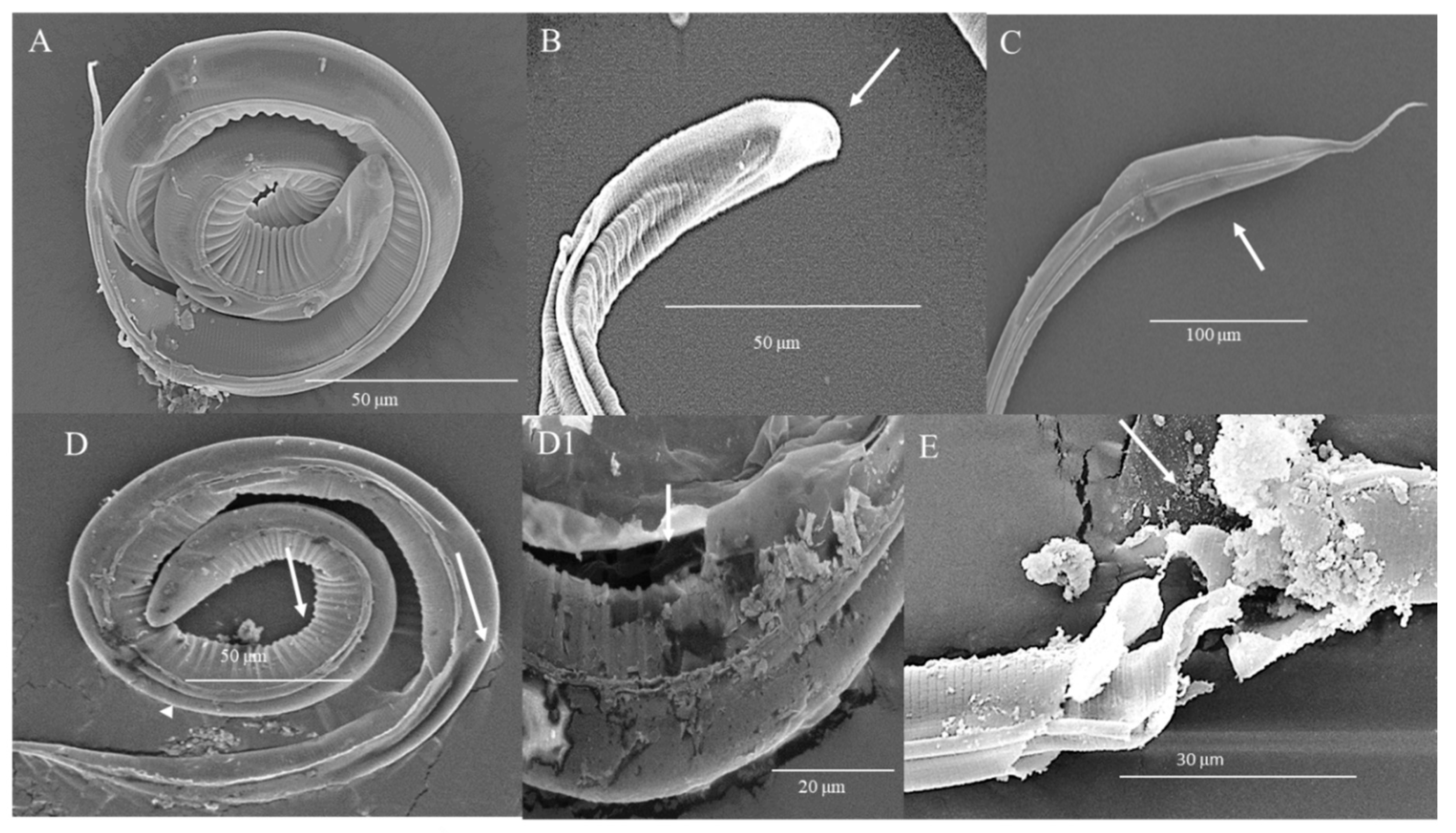
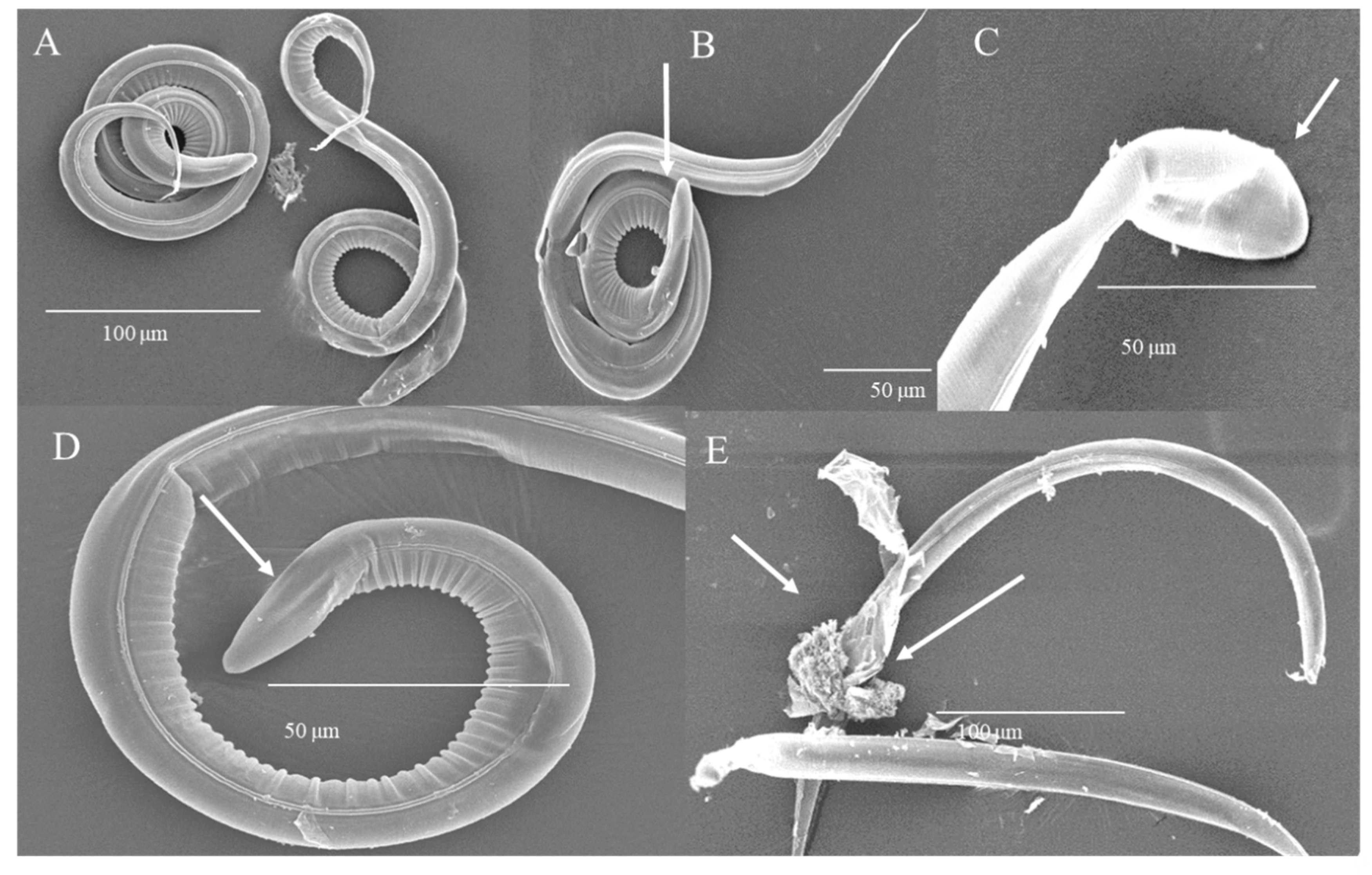
3.4. Damage of L3 Induced by HTE Observed by Scanning Electron Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Assefa, A.; Kechero, Y.; Tolemariam, T.; Kebede, A.; Shumi, E. Anthelmintic effects of indigenous multipurpose fodder tree extracts against Haemonchus contortus. Trop. Anim. Health Prod. 2017, 50, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.W.; Chandrawathani, P. Haemonchus contortus: Parasite problem No. 1 from tropics—Polar Circle. Problems and prospects for control based on epidemiology. Trop. Biomed. 2005, 22, 131–137. [Google Scholar] [PubMed]
- De Cezaro, M.C.; Tvarijonaviciute, A.; Tecles, F.; Cerón, J.J.; Eckersall, D.P.; Ferreira, J.C.; Schmidt, E.M.S. Changes in biochemical analytes in calves infected by nematode parasites in field conditions. Vet. Parasitol. 2016, 219, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabien, M. Livestock Nematode Infection in a Changing World: Investigating the European Situation; University of Zurich: Zürich, Switzerland, 2016; p. 32. [Google Scholar] [CrossRef]
- Rochfort, S.; Parker, A.J.; Dunshea, F.R. Plant bioactives for ruminant health and productivity. Phytochemistry 2008, 69, 299–322. [Google Scholar] [CrossRef] [Green Version]
- Hoste, H.; Martinez-Ortiz-De-Montellano, C.; Manolaraki, F.; Brunet, S.; Ojeda-Robertos, N.; Fourquaux, I.; Torres-Acosta, J.; Sandoval-Castro, C. Direct and indirect effects of bioactive tannin-rich tropical and temperate legumes against nematode infections. Vet. Parasitol. 2012, 186, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadou, S.; Kyriazakis, I.; Jackson, F.; Coop, R. Direct anthelmintic effects of condensed tannins towards different gastrointestinal nematodes of sheep: In vitro and in vivo studies. Vet. Parasitol. 2001, 99, 205–219. [Google Scholar] [CrossRef]
- Min, B.R.; Hart, S.P. Tannins for suppression of internal parasites. J. Anim. Sci. 2003, 81, 102–109. [Google Scholar]
- Engström, M.T.; Karonen, M.; Ahern, J.R.; Baert, N.; Payre, B.; Hoste, H.; Salminen, J. Chemical Structures of Plant Hydrolyzable Tannins Reveal Their in Vitro Activity against Egg Hatching and Motility of Haemonchus contortus Nematodes. J. Agric. Food Chem. 2016, 64, 840–851. [Google Scholar] [CrossRef]
- Corona-Palazuelos, M.; Murillo-Ayala, E.; Castro-del Campo, N.; Romo-Rubio, J.; Cervantes-Pacheco, B.; Gaxiola-Camacho, S.; Barajas-Cruz, R. Influence of tannin extract addition on the amount of nematodes found in feedlot calves at the beginning of the fattening process. Agrociencia 2016, 50, 1013–1025. [Google Scholar]
- Acevedo-Ramírez, P.M.D.C.; Hallal-Calleros, C.; Flores-Pérez, F.I.; Alba-Hurtado, F.; Mendoza-Garfías, M.B.; Del Campo, N.C.; Barajas, R. Anthelmintic effect and tissue alterations induced in vitro by hydrolysable tannins on the adult stage of the gastrointestinal nematode Haemonchus contortus. Vet. Parasitol. 2019, 266, 1–6. [Google Scholar] [CrossRef]
- Krisper, P.; Tisler, V.; Skubic, I.; Rupnik, Q.; Kobal, S. The Use of Tannin from Chestnut (Castanea vesca). In Plant Poly Phenols; Hemingway, R.W., Laks, P.E., Eds.; Plenum Press: New York, NY, USA, 1992. [Google Scholar]
- Campo, M.; Pinelli, P.; Romani, A. Hydrolyzable Tannins from Sweet Chestnut Fractions Obtained by a Sustainable and Eco-friendly Industrial Process. Nat. Prod. Commun. 2016, 11, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcão, L.; Araújo, M. Vegetable Tannins Used in the Manufacture of Historic Leathers. Molecules 2018, 23, 1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kardel, M.; Taube, F.; Schulz, H.; Schutze, W.; Gierus, M. Different approaches to evaluate tannin content and structure of selected plant extracts—Review and new aspects. J. Appl. Bot. Food Qual. 2013, 86, 154–166. [Google Scholar] [CrossRef]
- Pizzi, A. Tannins: Prospectives and Actual Industrial Applications. Biomolecules 2019, 9, 344. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Amézquita, R.A.; Morales-Montor, J.; Muñoz-Guzmán, M.A.; Nava-Castro, K.E.; Ramírez-Álvarez, H.; Cuenca-Verde, C.; Moreno-Mendoza, N.A.; Cuéllar-Ordaz, J.A.; Alba-Hurtado, F. Progesterone inhibits the in vitro L3/L4 molting process in Haemonchus contortus. Vet. Parasitol. 2017, 248, 48–53. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle; National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- Perez-Ponce de León, G.; Pinacho-Pinacho, C.; Mendoza-Garfias, B.; Choudhury, A.; García-Varela, M.J. Phylogenetic analysis using the 28S rRNA gene reveals that the genus Paracreptoma (Digenea: Allocreadiidae) is not monophyletic; description of two genera and one new species. J. Parasitol. 2016, 102, 131–142. [Google Scholar] [CrossRef]
- Jamous, R.M.; Shtayeh, M.S.A.; Abu-Zaitoun, S.Y.; Markovics, A.; Azaizeh, H. Effects of selected Palestinian plants on the in vitro exsheathment of the third stage larvae of gastrointestinal nematodes. BMC Vet. Res. 2017, 13, 308. [Google Scholar] [CrossRef] [Green Version]
- Zabré, G.; Kaboré, A.; Bayala, B.; Katiki, L.M.; Costa-Júnior, L.M.; Tamboura, H.H.; Belem, A.M.; Abdalla, A.L.; Niderkorn, V.; Hoste, H.; et al. Comparison of thein vitroanthelmintic effects ofAcacia niloticaandAcacia raddiana. Parasite 2017, 24, 44. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Ramírez, G.; Mathieu, C.; Vilarem, G.; Hoste, H.; Mendoza-De-Gives, P.; González-Pech, P.G.; Torres-Acosta, J.F.J.; Sandoval-Castro, C. Age of Haemonchus contortus third stage infective larvae is a factor influencing the in vitro assessment of anthelmintic properties of tannin containing plant extracts. Vet. Parasitol. 2017, 243, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Bahuaud, D.; Martinez-Ortiz de Montellano, C.; Chauveau, S.; Prevot, F.; Torres-Acosta, F.; Fouraste, I.; Hoste, H. Effects of four tanniferous plant extracts on the in vitro exsheathment of third-stage larvae of parasitic nematodes. Parasitology 2006, 132, 545–554. [Google Scholar] [CrossRef]
- Brunet, S.; De Montellano, C.M.-O.; Torres-Acosta, J.; Sandoval-Castro, C.; Aguilar-Caballero, A.; Capetillo-Leal, C.; Hoste, H. Effect of the consumption of Lysiloma latisiliquum on the larval establishment of gastrointestinal nematodes in goats. Vet. Parasitol. 2008, 157, 81–88. [Google Scholar] [CrossRef] [PubMed]
- De Hoyos-Martínez, P.L.; Merle, J.; Labidi, J.; Bouhtoury, F.C.-E. Tannins extraction: A key point for their valorization and cleaner production. J. Clean. Prod. 2019, 206, 1138–1155. [Google Scholar] [CrossRef] [Green Version]
- Féboli, A.; Laurentiz, A.C.; Soares, S.C.; Augusto, J.G.; Anjos, L.A.; Magalhães, L.G.; Filardi, R.S.; Laurentiz, R.D.S.D. Ovicidal and larvicidal activity of extracts of Opuntia ficus-indica against gastrointestinal nematodes of naturally infected sheep. Vet. Parasitol. 2016, 226, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Amiji, M.M. Complexation and Protein Binding. In Applied Physical Pharmacy; Amiji, M.M., Cook, T.J., Mobley, C., Eds.; McGraw-Hill Education: New York, NY, USA, 2014. [Google Scholar]
- Hagerman, A.E. Chemistry of Tannin-Protein Complexation. In Chemistry and Significance of Condensed Tannins; Hemingway, R.W., Karchesy, J.J., Branham, S.J., Eds.; Springer: Boston, MA, USA, 1989. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acevedo-Ramírez, P.M.d.C.; Hallal-Calleros, C.; Flores-Pérez, I.; Alba-Hurtado, F.; Mendoza-Garfias, M.B.; Barajas, R. Nematicidal Effect and Histological Modifications Induced by Hydrolysable Tannin Extract on the Third-Stage Infective Larvae of Haemonchus contortus. Biology 2020, 9, 442. https://doi.org/10.3390/biology9120442
Acevedo-Ramírez PMdC, Hallal-Calleros C, Flores-Pérez I, Alba-Hurtado F, Mendoza-Garfias MB, Barajas R. Nematicidal Effect and Histological Modifications Induced by Hydrolysable Tannin Extract on the Third-Stage Infective Larvae of Haemonchus contortus. Biology. 2020; 9(12):442. https://doi.org/10.3390/biology9120442
Chicago/Turabian StyleAcevedo-Ramírez, Perla María del Carmen, Claudia Hallal-Calleros, Iván Flores-Pérez, Fernando Alba-Hurtado, María Berenit Mendoza-Garfias, and Rubén Barajas. 2020. "Nematicidal Effect and Histological Modifications Induced by Hydrolysable Tannin Extract on the Third-Stage Infective Larvae of Haemonchus contortus" Biology 9, no. 12: 442. https://doi.org/10.3390/biology9120442