A Novel Antimicrobial Peptide (Kassinatuerin-3) Isolated from the Skin Secretion of the African Frog, Kassina senegalensis

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Biodata and Secretion Acquisition
2.2. Molecular Cloning of Kassinatuerin-3 Precursor-Encoding cDNA
2.3. Isolation of Kassinatuerin-3 from the Skin Secretion
2.4. Solid-Phase Peptide Synthesis
2.5. Circular Dichorism
2.6. Antimicrobial Assays
2.7. Hemolysis Assay
2.8. Anti-Biofilm Assay
2.9. Cell Cytotoxicity
3. Results
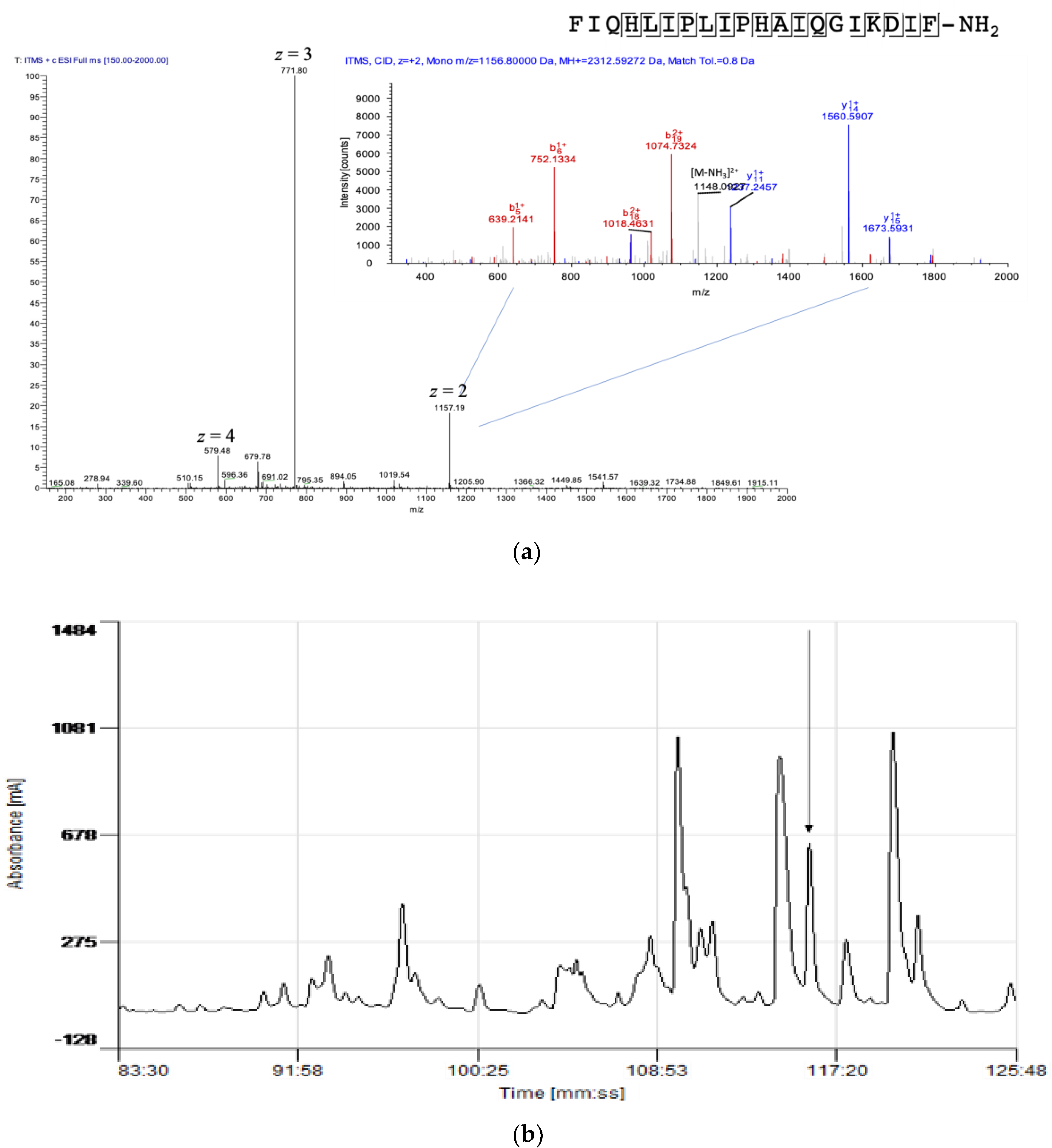
3.1. Molecular Cloning of Kassinatuerin-3 Peptide Precursor Transcript
3.2. Identification and Structural Analysis of Kassinatuerin-3
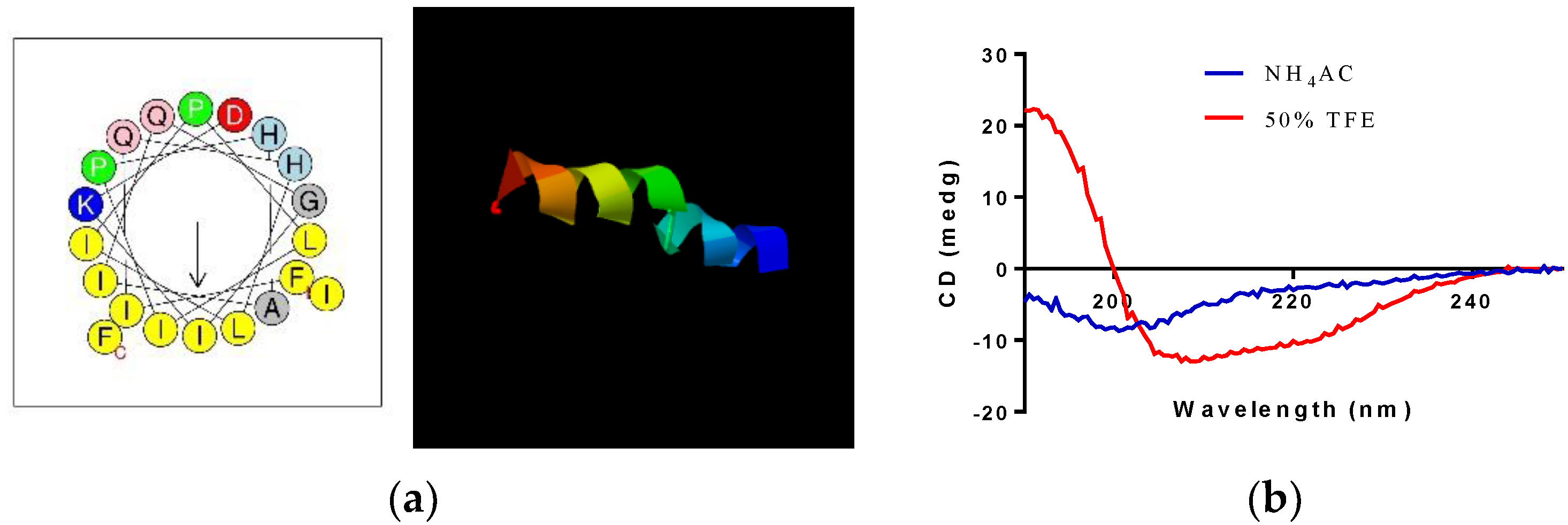
3.3. Prediction of Secondary Structure and Physiochemical Properties
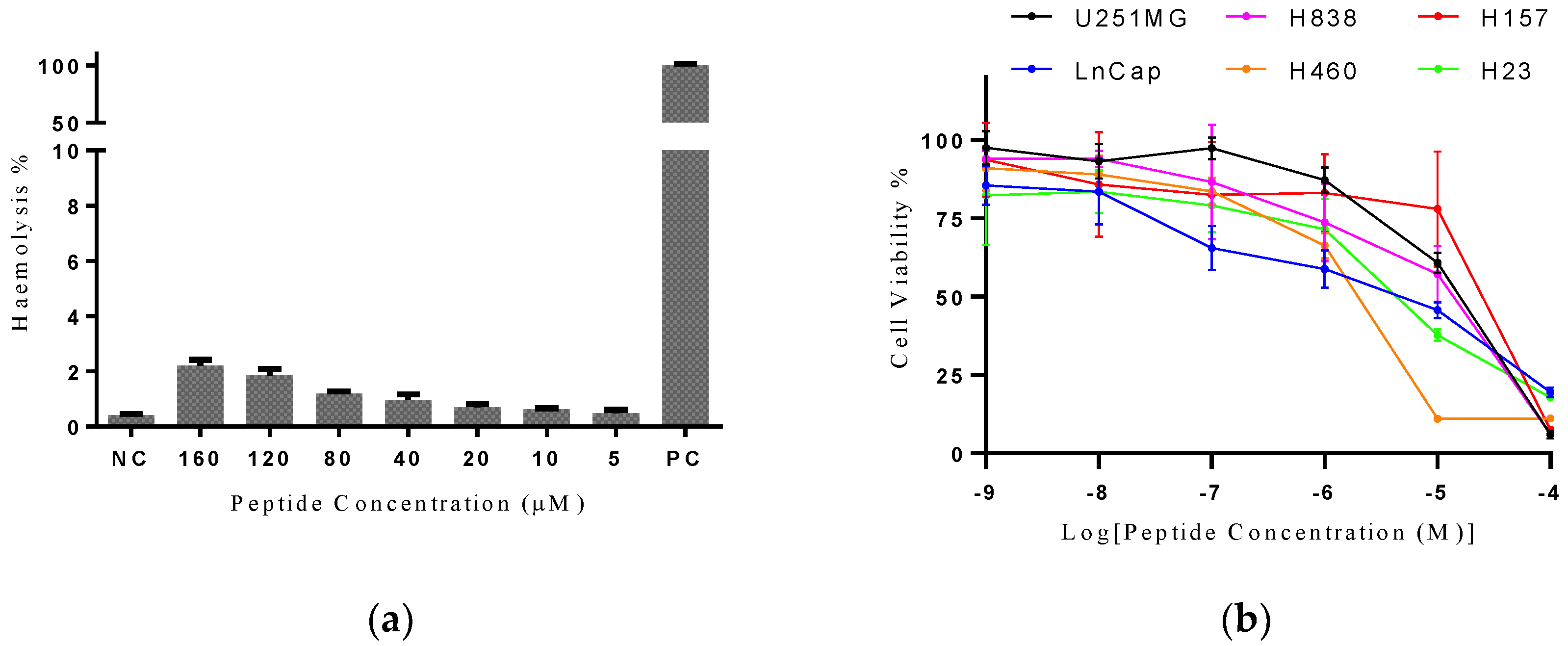
3.4. Antimicrobial and Hemolysis Assays
3.5. Antibiofilm Assay
3.6. Hemolytic and Antiproliferative Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, J.M. The contribution of skin antimicrobial peptides to the system of innate immunity in anurans. Cell Tissue Res. 2011, 343, 201–212. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Renner, L.D.; Zan, J.; Hu, L.I.; Martinez, M.; Resto, P.J.; Siegel, A.C.; Torres, C.; Hall, S.B.; Slezak, T.R.; Nguyen, T.H. Detection of ESKAPE bacterial pathogens at the point of care using isothermal DNA-based assays in a portable degas-actuated microfluidic diagnostic assay platform. Appl. Environ. Microbiol. 2017, 83, e02449-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Pozo, J.; Serrera, A.; Martinez-Cuesta, A.; Leiva, J.; Penades, J.; Lasa, I. Biofilm related infections: Is there a place for conservative treatment of port-related bloodstream infections? Int. J. Artif. Organs 2006, 29, 379–386. [Google Scholar] [CrossRef]
- Martin, E.; Ganz, T.; Lehrer, R.I. Defensins and other endogenous peptide antibiotics of vertebrates. J. Leukoc. Biol. 1995, 58, 128–136. [Google Scholar] [CrossRef]
- Hancock, R.E.; Rozek, A. Role of membranes in the activities of antimicrobial cationic peptides. FEMS Microbiol. Lett. 2002, 206, 143–149. [Google Scholar] [CrossRef]
- Pletzer, D.; Hancock, R.E. Antibiofilm peptides: Potential as broad-spectrum agents. J. Bacteriol. 2016, 198, 2572–2578. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, G. APD: The antimicrobial peptide database. Nucleic Acids Res. 2004, 32, D590–D592. [Google Scholar] [CrossRef] [Green Version]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial peptides: Amphibian host defense peptides. Curr. Med. Chem. 2019, 26, 5924–5946. [Google Scholar] [CrossRef]
- Conlon, J.M. Structural diversity and species distribution of host-defense peptides in frog skin secretions. Cell. Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Amiche, M.; Ladram, A.; Nicolas, P. A consistent nomenclature of antimicrobial peptides isolated from frogs of the subfamily Phyllomedusinae. Peptides 2008, 29, 2074–2082. [Google Scholar] [CrossRef]
- Simmaco, M.; Kreil, G.; Barra, D. Bombinins, antimicrobial peptides from Bombina species. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 1551–1555. [Google Scholar] [CrossRef] [Green Version]
- Conlon, J.M.; Mechkarska, M.; King, J.D. Host-defense peptides in skin secretions of African clawed frogs (Xenopodinae, Pipidae). Gen. Comp. Endocrinol. 2012, 176, 513–518. [Google Scholar] [CrossRef]
- Roudi, R.; Syn, N.L.; Roudbary, M. Antimicrobial peptides as biologic and immunotherapeutic agents against cancer: A comprehensive overview. Front. Immunol. 2017, 8, 1320. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Hooper, L.V. Antimicrobial defense of the intestine. Immunity 2015, 42, 28–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipsky, B.A.; Holroyd, K.J.; Zasloff, M. Topical versus systemic antimicrobial therapy for treating mildly infected diabetic foot ulcers: A randomized, controlled, double-blinded, multicenter trial of pexiganan cream. Clin. Infect. Dis. 2008, 47, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lashua, L.P.; Melvin, J.A.; Deslouches, B.; Pilewski, J.M.; Montelaro, R.C.; Bomberger, J.M. Engineered cationic antimicrobial peptide (eCAP) prevents Pseudomonas aeruginosa biofilm growth on airway epithelial cells. J. Antimicrob. Chemother. 2016, 71, 2200–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, H.M.; Wiseman, L.R. Pexiganan acetate. Drugs 1998, 56, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- König, E.; Bininda-Emonds, O.R.; Shaw, C. The diversity and evolution of anuran skin peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef] [PubMed]
- Resnick, N.M.; Maloy, W.L.; Guy, H.R.; Zasloff, M. A novel endopeptidase from Xenopus that recognizes α-helical secondary structure. Cell 1991, 66, 541–554. [Google Scholar] [CrossRef]
- DARBY, N.J.; LACKEY, D.B.; SMYTH, D.G. Purification of a cysteine endopeptidase which is secreted with bioactive peptides from the epidermal glands of Xenopus laevis. Eur. J. Biochem. 1991, 195, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Shimoi, H.; Iwasaki, Y.; Kawahara, T.; Matsuura, Y.; Nishikawa, Y. Elucidation of amidating reaction mechanism by frog amidating enzyme, peptidylglycine alpha-hydroxylating monooxygenase, expressed in insect cell culture. EMBO J. 1990, 9, 4259–4265. [Google Scholar] [CrossRef] [PubMed]
- Savelyeva, A.; Ghavami, S.; Davoodpour, P.; Asoodeh, A.; Łos, M.J. An overview of Brevinin superfamily: Structure, function and clinical perspectives. In Anticancer Genes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 197–212. [Google Scholar]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial peptides from the skins of North American frogs. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 1556–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasi, A.; Montecucchi, P.; Erspamer, V.; Visser, J. Amino acid composition and sequence of kassinin, a tachykinin dodecapeptide from the skin of the African frog Kassina senegalensis. Experientia 1977, 33, 857–858. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Feng, W.; Zhou, M.; Ma, C.; Chen, T.; Zeller, M.; Hornshaw, M.; Wang, L.; Shaw, C. Kasstasin: A novel potent vasoconstrictor peptide from the skin secretion of the African red-legged running frog, Kassina maculata. Biochimie 2011, 93, 1537–1542. [Google Scholar] [CrossRef]
- Chen, T.; Reid, C.N.; Walker, B.; Zhou, M.; Shaw, C. Kassinakinin S: A novel histamine-releasing heptadecapeptide from frog (Kassina senegalensis) skin secretion. Biochem. Biophys. Res. Commun. 2005, 337, 474–480. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, M.; McGrath, S.; Chen, T.; Gorman, S.P.; Walker, B.; Shaw, C. A family of kassinatuerin-2 related peptides from the skin secretion of the African hyperoliid frog, Kassina maculata. Peptides 2009, 30, 1428–1433. [Google Scholar] [CrossRef]
- Mattute, B.; Knoop, F.C.; Conlon, J.M. Kassinatuerin-1: A peptide with broad-spectrum antimicrobial activity isolated from the skin of the hyperoliid frog, Kassina senegalensis. Biochem. Biophys. Res. Commun. 2000, 268, 433–436. [Google Scholar] [CrossRef]
- Wang, H.; Li, R.; Xi, X.; Meng, T.; Zhou, M.; Wang, L.; Zhang, Y.; Chen, T.; Shaw, C. Senegalin: A novel antimicrobial/myotropic hexadecapeptide from the skin secretion of the African running frog, Kassina senegalensis. Amino Acids 2013, 44, 1347–1355. [Google Scholar] [CrossRef]
- Chen, H.; Wang, L.; Zeller, M.; Hornshaw, M.; Wu, Y.; Zhou, M.; Li, J.; Hang, X.; Cai, J.; Chen, T. Kassorins: Novel innate immune system peptides from skin secretions of the African hyperoliid frogs, Kassina maculata and Kassina senegalensis. Mol. Immunol. 2011, 48, 442–451. [Google Scholar] [CrossRef]
- Conlon, J.M.; Abraham, B.; Galadari, S.; Knoop, F.C.; Sonnevend, A.; Pál, T. Antimicrobial and cytolytic properties of the frog skin peptide, kassinatuerin-1 and its L-and D-lysine-substituted derivatives. Peptides 2005, 26, 2104–2110. [Google Scholar] [CrossRef] [PubMed]
- Tyler, M.J.; Stone, D.J.; Bowie, J.H. A novel method for the release and collection of dermal, glandular secretions from the skin of frogs. J. Pharmacol. Toxicol. Methods 1992, 28, 199–200. [Google Scholar] [CrossRef]
- Du, Q.; Wang, H.; Ma, C.; Wu, Y.; Xi, X.; Zhou, M.; Chen, T.; Shaw, C.; Wang, L. Identification of a novel vasodilatory octapeptide from the skin secretion of the African hyperoliid frog, Kassina senegalensis. Molecules 2017, 22, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Gao, Y.; Wang, L.; Xi, X.; Wu, Y.; Zhou, M.; Zhang, Y.; Ma, C.; Chen, T.; Shaw, C. A combined molecular cloning and mass spectrometric method to identify, characterize, and design frenatin peptides from the skin secretion of Litoria infrafrenata. Molecules 2016, 21, 1429. [Google Scholar] [CrossRef]
- Wu, D.; Gao, Y.; Tan, Y.; Liu, Y.; Wang, L.; Zhou, M.; Xi, X.; Ma, C.; Bininda-Emonds, O.R.; Chen, T. Discovery of distinctin-like-peptide-ph (dlp-ph) from the skin secretion of phyllomedusa hypochondrialis, a prototype of a novel family of antimicrobial peptide. Front. Microbiol. 2018, 9, 541. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Zhou, M.; Duan, J.; Bininda-Emonds, O.R.; Chen, T. Targeted modification of a novel amphibian antimicrobial peptide from Phyllomedusa tarsius to enhance its activity against MRSA and microbial biofilm. Front. Microbiol. 2017, 8, 628. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-ph: A novel peptide with antimicrobial and anticancer activities from the skin secretion of the south american orange-legged leaf frog, pithecopus (phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef]
- Barceloux, D.G. Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Ong, J.M.; da Cruz, L. The bionic eye: A review. Clin. Exp. Ophthalmol. 2012, 40, 6–17. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, C.; Lai, R. Antimicrobial peptides from amphibians. Biomol. Concepts 2011, 2, 27–38. [Google Scholar] [CrossRef]
- Barra, D.; Simmaco, M. Amphibian skin: A promising resource for antimicrobial peptides. Trends Biotechnol. 1995, 13, 205–209. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Barra, D. Antimicrobial peptides from amphibian skin: What do they tell us? Pept. Sci. 1998, 47, 435–450. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug resistance in bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, L.B. Antimicrobial resistance in gram-positive bacteria. Am. J. Infect. Control 2006, 34, S11–S19. [Google Scholar] [CrossRef] [PubMed]
- Brand, G.D.; Leite, J.R.S.; de Sá Mandel, S.M.; Mesquita, D.A.; Silva, L.P.; Prates, M.V.; Barbosa, E.A.; Vinecky, F.; Martins, G.R.; Galasso, J.H. Novel dermaseptins from Phyllomedusa hypochondrialis (Amphibia). Biochem. Bioph. Res. Commun. 2006, 347, 739–746. [Google Scholar] [CrossRef]
- Thompson, A.H.; Bjourson, A.J.; Orr, D.F.; Shaw, C.; McClean, S. A combined mass spectrometric and cDNA sequencing approach to the isolation and characterization of novel antimicrobial peptides from the skin secretions of Phyllomedusa hypochondrialis azurea. Peptides 2007, 28, 1331–1343. [Google Scholar] [CrossRef]
- Kückelhaus, S.; Leite, J.R.S.; Neves, M.P.; Frota, K.S.; Abdala, L.F.; Muniz-Junqueira, M.I.; Bloch, C.; Tosta, C.E. Toxicity evaluation to mice of phylloseptin-1, an antimicrobial peptide from the skin secretion of Phyllomedusa hypochondrialis (Amphibia). Int. J. Pept. Res. Ther. 2007, 13, 423–429. [Google Scholar] [CrossRef]
- Duda, T.F., Jr.; Vanhoye, D.; Nicolas, P. Roles of diversifying selection and coordinated evolution in the evolution of amphibian antimicrobial peptides. Mol. Biol. Evol. 2002, 19, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.-C.; Hui, L.; Chen, H.M. Enhancement of the cytolytic effect of anti-bacterial cecropin by the microvilli of cancer cells. Anticancer Res. 1998, 18, 4467–4474. [Google Scholar]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moosazadeh Moghaddam, M.; Mirnejad, R. Antimicrobial peptides: Features, action, and their resistance mechanisms in bacteria. Microb. Drug Resist. 2018, 24, 747–767. [Google Scholar] [CrossRef]
- Beveridge, T.J. Structures of gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [CrossRef] [Green Version]
- Sani, M.-A.; Henriques, S.T.; Weber, D.; Separovic, F. Bacteria may cope differently from similar membrane damage caused by the Australian tree frog antimicrobial peptide maculatin 1.1. J. Biol. Chem. 2015, 290, 19853–19862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, R.; Joshi, M.; Mohanram, H.; Bhunia, A.; Mangoni, M.L.; Bhattacharjya, S. NMR structure of temporin-1 ta in lipopolysaccharide micelles: Mechanistic insight into inactivation by outer membrane. PLoS ONE 2013, 8, e72718. [Google Scholar] [CrossRef] [PubMed]
- Kunda, N.K. Antimicrobial peptides as novel therapeutics for nonsmall cell lung cancer. Drug Discov. Today 2019, 25, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta (BBA) Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, R.M.; Srivastava, S.; Singh, M.; Bajpai, V.K.; Ghosh, J.K. Consequences of alteration in leucine zipper sequence of melittin in its neutralization of lipopolysaccharide-induced proinflammatory response in macrophage cells and interaction with lipopolysaccharide. J. Biol. Chem. 2012, 287, 1980–1995. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, C.; Hamadat, S.; Le Saux, K.; Newton, C.; Mazouni, M.; Zargarian, L.; Miro-Padovani, M.; Zadigue, P.; Delbé, J.; Hamma-Kourbali, Y. Studies of the antitumor mechanism of action of dermaseptin B2, a multifunctional cationic antimicrobial peptide, reveal a partial implication of cell surface glycosaminoglycans. PLoS ONE 2017, 12, e0182926. [Google Scholar] [CrossRef] [Green Version]
- Makovitzki, A.; Fink, A.; Shai, Y. Suppression of human solid tumor growth in mice by intratumor and systemic inoculation of histidine-rich and pH-dependent host defense–like lytic peptides. Cancer Res. 2009, 69, 3458–3463. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrophobicity <H> | Hydrophobic Moment <µH> | Net Charge z |
|---|---|---|
| 0.880 | 0.570 | +1 |
| Kassinatuerin-3 | Melittin | |||
|---|---|---|---|---|
| MIC a (µM) | MBC b (µM) | MIC (µM) | MBC (µM) | |
| S. aureus | 16–32 | 32–64 | 1–2 | 2–4 |
| E. coli | >512 | >512 | 1–4 | 8–16 |
| C. albicans | 64-128 | 128 | 2–8 | 16–32 |
| MRSA | 32–64 | 64–128 | 1–2 | 2–4 |
| E. faecalis | 128 | 128 | 1–2 | 4 |
| P. aeruginosa | >512 | >512 | 16–32 | 32–64 |
| Kassinatuerin-3 | Melittin | |||
|---|---|---|---|---|
| MBIC a (µM) | MBEC b (µM) | MBIC (µM) | MBEC (µM) | |
| S. aureus | 16–32 | 64–128 | 1–4 | 16–32 |
| E. coli | >512 | >512 | 4–8 | 16–32 |
| C. albicans | 128 | >512 | 4–16 | 32–64 |
| MRSA | 32–64 | 128 | 1–4 | 16–32 |
| E. faecalis | 128 | 256-512 | 4–8 | 16–32 |
| P. aeruginosa | >512 | >512 | 32–64 | 128–256 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; He, H.; Chen, X.; Zhou, M.; Wei, M.; Xi, X.; Ma, C.; Du, Q.; Chen, T.; Shaw, C.; et al. A Novel Antimicrobial Peptide (Kassinatuerin-3) Isolated from the Skin Secretion of the African Frog, Kassina senegalensis. Biology 2020, 9, 148. https://doi.org/10.3390/biology9070148
Wang H, He H, Chen X, Zhou M, Wei M, Xi X, Ma C, Du Q, Chen T, Shaw C, et al. A Novel Antimicrobial Peptide (Kassinatuerin-3) Isolated from the Skin Secretion of the African Frog, Kassina senegalensis. Biology. 2020; 9(7):148. https://doi.org/10.3390/biology9070148
Chicago/Turabian StyleWang, Hui, Haoyang He, Xiaoling Chen, Mei Zhou, Minjie Wei, Xinping Xi, Chengbang Ma, Qiang Du, Tianbao Chen, Chris Shaw, and et al. 2020. "A Novel Antimicrobial Peptide (Kassinatuerin-3) Isolated from the Skin Secretion of the African Frog, Kassina senegalensis" Biology 9, no. 7: 148. https://doi.org/10.3390/biology9070148
APA StyleWang, H., He, H., Chen, X., Zhou, M., Wei, M., Xi, X., Ma, C., Du, Q., Chen, T., Shaw, C., & Wang, L. (2020). A Novel Antimicrobial Peptide (Kassinatuerin-3) Isolated from the Skin Secretion of the African Frog, Kassina senegalensis. Biology, 9(7), 148. https://doi.org/10.3390/biology9070148