Enhanced Thermostability and Enzymatic Activity of cel6A Variants from Thermobifida fusca by Empirical Domain Engineering


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. PCR Amplification of cel6A Variants
2.3. Sub Cloning of cel6A Variants in Expression Vector
2.4. Expression Analysis of cel6A Variants
2.5. Enzymatic Activity Assay
2.6. pH Stability Determination
2.7. Thermal Stability Determination
2.8. Molecular Modeling Study
3. Results and Discussion
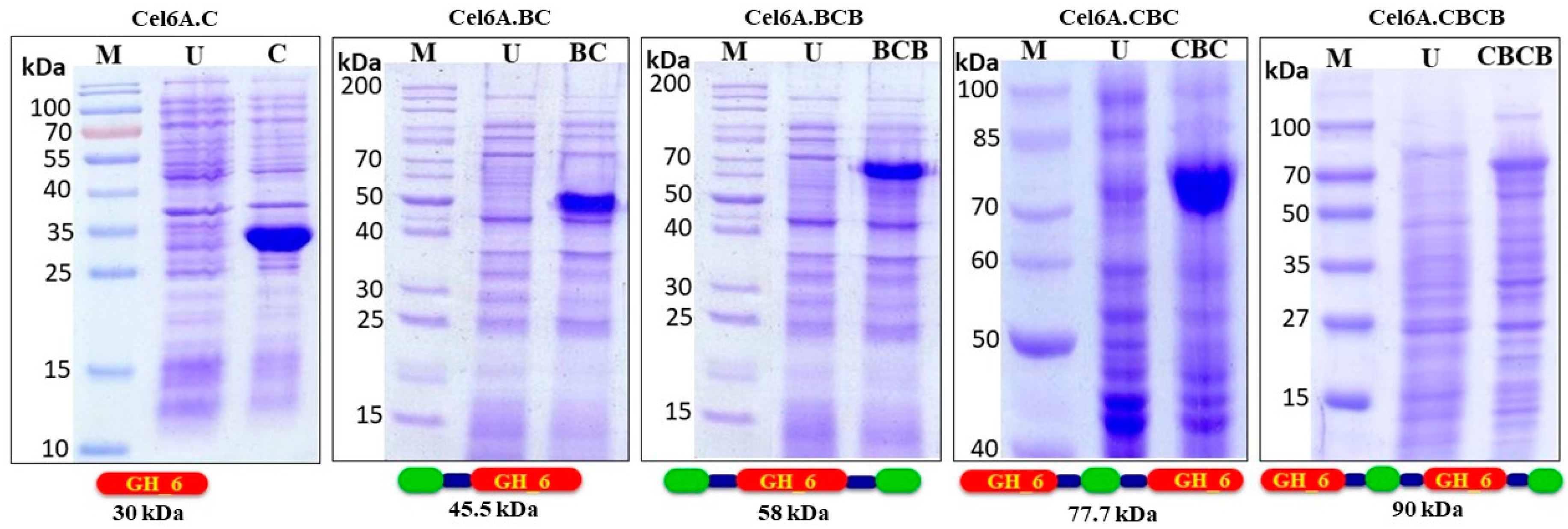
3.1. Construction of Plasmids for Domain Engineered cel6A Variants
3.2. Expression Analysis and Enzymatic Activities of cel6A Variants
3.3. Physiological Characterization of cel6A Variants
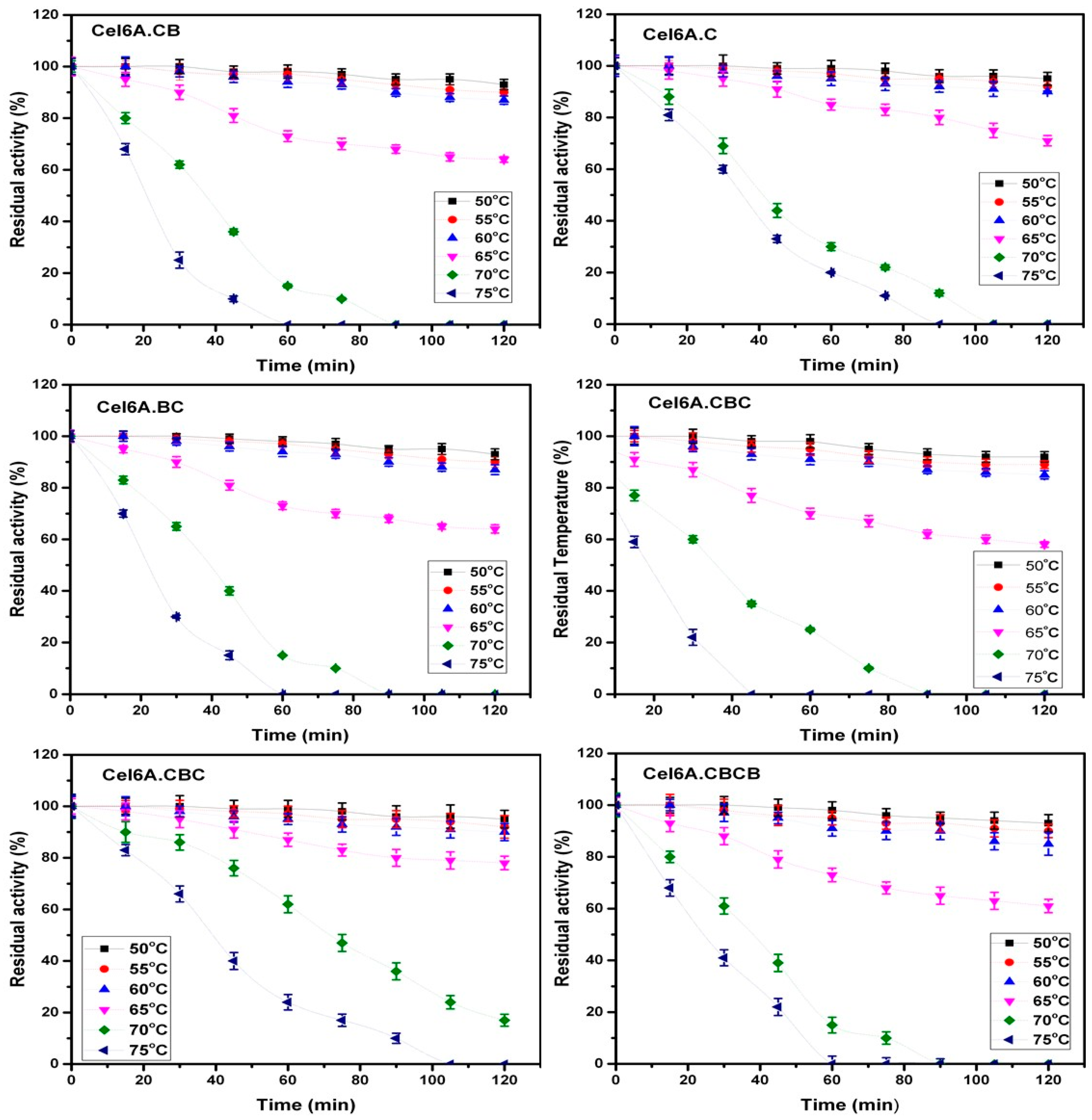
3.3.1. Effect of Temperature on cel6A Variants
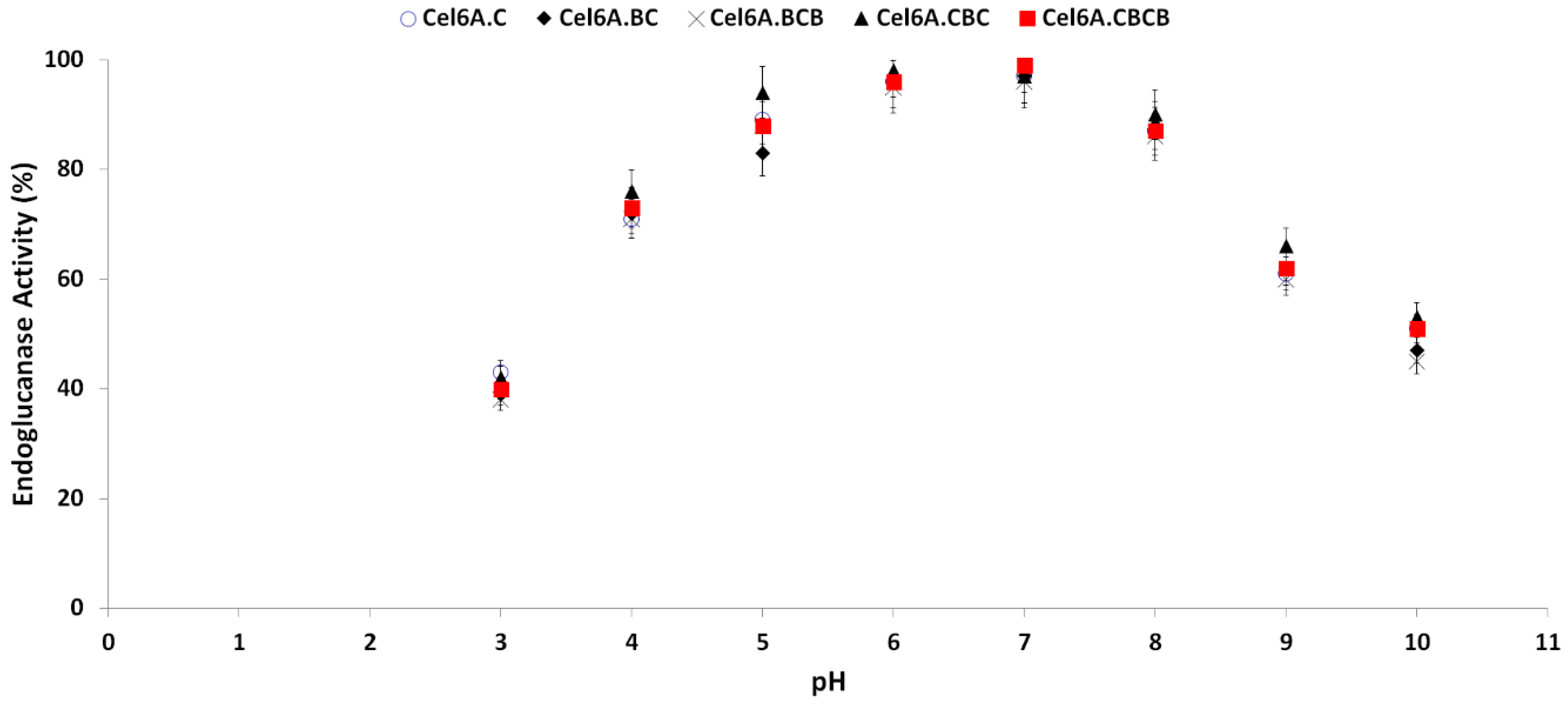
3.3.2. Effect of pH on cel6A Variants
3.4. Molecular Modeling Analysis
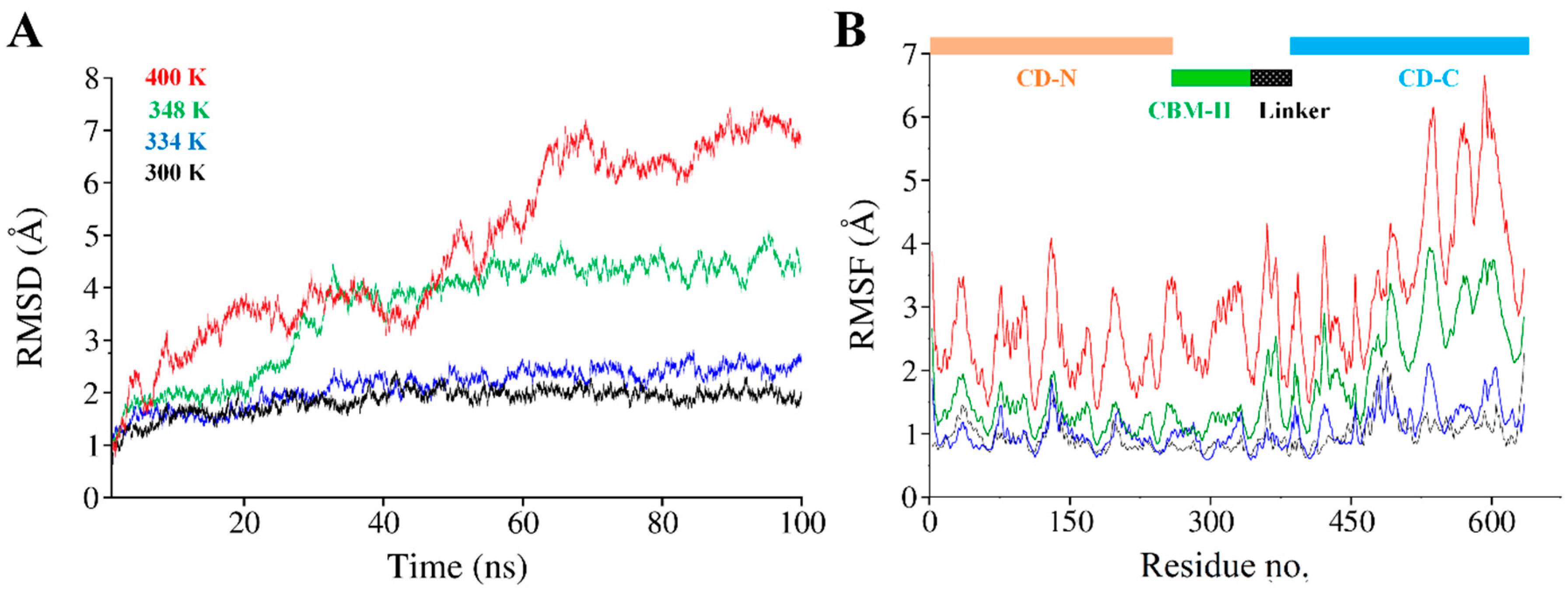
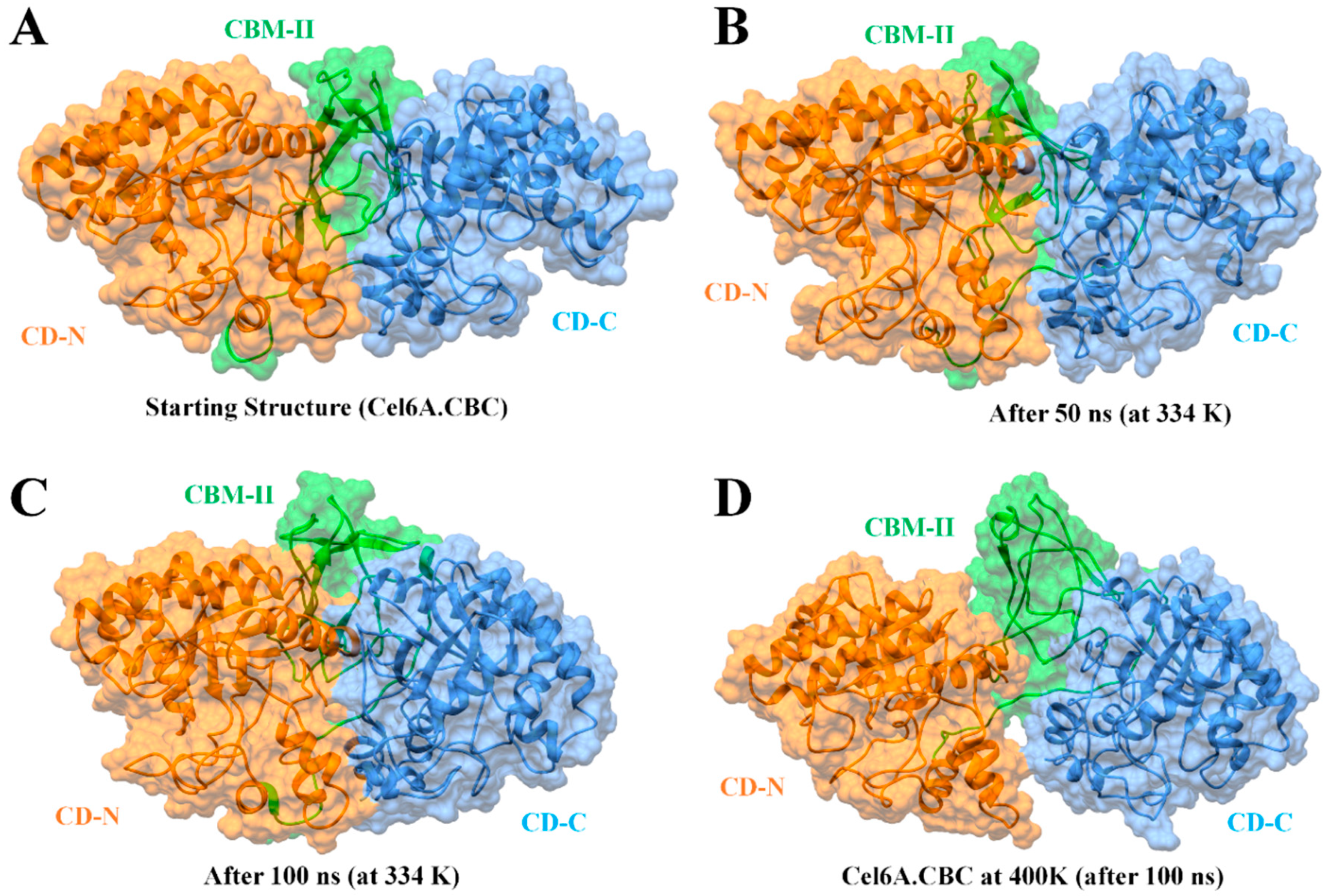
3.5. Global Structural Stability
3.6. Structural Flexibility
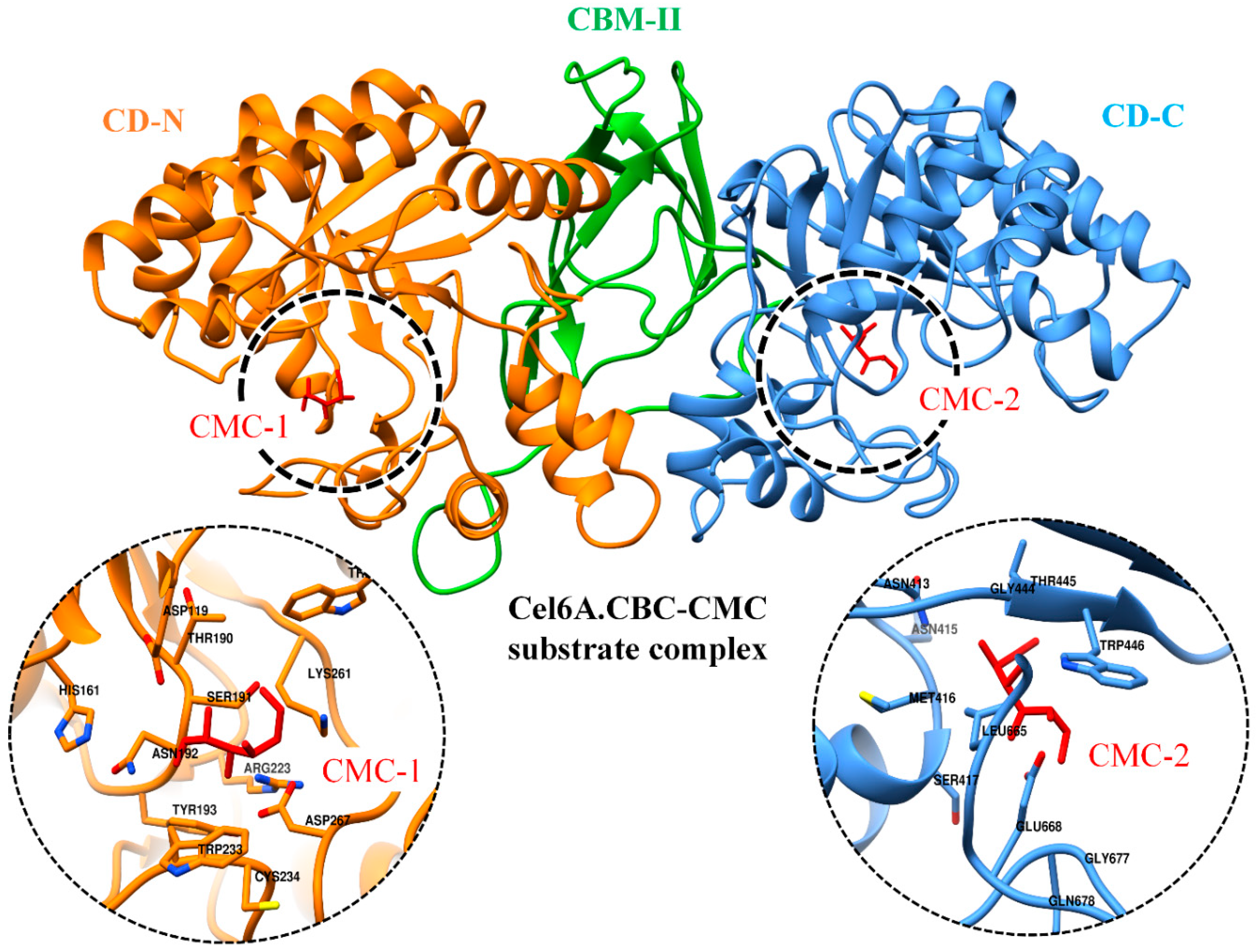
3.7. Binding Free Energy Calculations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heinzelman, P.; Snow, C.D.; Wu, I.; Nguyen, C.; Villalobos, A.; Govindarajan, S.; Minshull, J.; Arnold, F.H. A family of thermostable fungal cellulases created by structure-guided recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 5610–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanch, H.W.; Simmons, B.A.; Klein-Marcuschamer, D. Biomass deconstruction to sugars. Biotechnol. J. 2011, 6, 1086–1102. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.R.; Klein-Marcuschamer, D.; Stephanopoulos, G. Selection and optimization of microbial hosts for biofuels production. Metab. Eng. 2008, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.P.; Himmel, M.E.; Mielenz, J.R. Outlook for cellulase improvement: Screening and selection strategies. Biotechnol. Adv. 2006, 24, 452–481. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, X.-Z.; Zhang, Z.; Zhang, Y.-H.P. Engineering of Clostridium phytofermentans endoglucanase Cel5A for improved thermostability. Appl. Environ. Microbiol. 2010, 76, 4914–4917. [Google Scholar] [CrossRef] [Green Version]
- Paës, G.; O’Donohue, M.J. Engineering increased thermostability in the thermostable GH-11 xylanase from Thermobacillus xylanilyticus. J. Biotechnol. 2006, 125, 338–350. [Google Scholar] [CrossRef]
- Turner, P.; Mamo, G.; Karlsson, E.N. Potential and utilization of thermophiles and thermostable enzymes in biorefining. Microb. Cell Factories 2007, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Caspi, J.; Irwin, D.; Lamed, R.; Shoham, Y.; Fierobe, H.-P.; Wilson, D.B.; Bayer, E.A.J.B. Thermobifida fusca family-6 cellulases as potential designer cellulosome components. Biocatal. Biotransformation 2006, 24, 3–12. [Google Scholar]
- Wilson, D.B. Studies of Thermobifida fusca plant cell wall degrading enzymes. Chem. Rec. 2004, 4, 72–82. [Google Scholar] [CrossRef]
- Bolam, D.N.; Ciruela, A.; McQueen-Mason, S.; Simpson, P.; Williamson, M.P.; Rixon, J.E.; Boraston, A.; Hazlewood, G.P.; Gilbert, H.J. Pseudomonas cellulose-binding domains mediate their effects by increasing enzyme substrate proximity. Biochem. J. 1998, 331, 775–781. [Google Scholar] [CrossRef]
- Gilad, R.; Rabinovich, L.; Yaron, S.; Bayer, E.A.; Lamed, R.; Gilbert, H.J.; Shoham, Y. CelI, a noncellulosomal family 9 enzyme from Clostridium thermocellum, is a processive endoglucanase that degrades crystalline cellulose. J. Bacteriol. 2003, 185, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckham, G.T.; Bomble, Y.J.; Bayer, E.A.; Himmel, M.E.; Crowley, M.F. Applications of computational science for understanding enzymatic deconstruction of cellulose. Curr. Opin. Biotechnol. 2011, 22, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Din, N.; Gilkes, N.R.; Tekant, B.; Miller, R.C.; Warren, R.A.J.; Kilburn, D.G. Non–hydrolytic disruption of cellulose fibres by the binding domain of a bacterial cellulase. Nat. Biotechnol. 1991, 9, 1096–1099. [Google Scholar] [CrossRef]
- Cheng, G.; Liu, Z.; Murton, J.K.; Jablin, M.; Dubey, M.; Majewski, J.; Halbert, C.; Browning, J.; Ankner, J.; Akgun, B. Neutron reflectometry and QCM-D study of the interaction of cellulases with films of amorphous cellulose. Biomacromolecules 2011, 12, 2216–2224. [Google Scholar] [CrossRef] [PubMed]
- Viikari, L.; Alapuranen, M.; Puranen, T.; Vehmaanperä, J.; Siika-Aho, M. Thermostable enzymes in lignocellulose hydrolysis. In Biofuels; Springer: Berlin/Heidelberg, Germany, 2007; pp. 121–145. [Google Scholar]
- Kim, T.W.; Chokhawala, H.A.; Nadler, D.; Blanch, H.W.; Clark, D.S. Binding modules alter the activity of chimeric cellulases: Effects of biomass pretreatment and enzyme source. Biotechnol. Bioeng. 2010, 107, 601–611. [Google Scholar] [CrossRef]
- Anbar, M.; Gul, O.; Lamed, R.; Sezerman, U.O.; Bayer, E.A. Improved thermostability of Clostridium thermocellum endoglucanase Cel8A by using consensus-guided mutagenesis. Appl. Environ. Microbiol. 2012, 78, 3458–3464. [Google Scholar] [CrossRef] [Green Version]
- Anbar, M.; Lamed, R.; Bayer, E.A. Thermostability enhancement of Clostridium thermocellum cellulosomal endoglucanase Cel8A by a single glycine substitution. ChemCatChem 2010, 2, 997–1003. [Google Scholar] [CrossRef]
- Chen, X.; Li, W.; Ji, P.; Zhao, Y.; Hua, C.; Han, C. Engineering the conserved and noncatalytic residues of a thermostable β-1, 4-endoglucanase to improve specific activity and thermostability. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Mitrovic, A.; Flicker, K.; Steinkellner, G.; Gruber, K.; Reisinger, C.; Schirrmacher, G.; Camattari, A.; Glieder, A. Thermostability improvement of endoglucanase Cel7B from Hypocrea pseudokoningii. J. Mol. Catal. B Enzym. 2014, 103, 16–23. [Google Scholar] [CrossRef]
- Voutilainen, S.P.; Murray, P.G.; Tuohy, M.G.; Koivula, A. Expression of Talaromyces emersonii cellobiohydrolase Cel7A in Saccharomyces cerevisiae and rational mutagenesis to improve its thermostability and activity. Protein Eng. Des. Sel. 2010, 23, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.-L.; Pei, X.-Q.; Wu, Z.-L. Introduction of glycine and proline residues onto protein surface increases the thermostability of endoglucanase CelA from Clostridium thermocellum. Bioresour. Technol. 2011, 102, 3636–3638. [Google Scholar] [CrossRef] [PubMed]
- Burstein, T.; Shulman, M.; Jindou, S.; Petkun, S.; Frolow, F.; Shoham, Y.; Bayer, E.A.; Lamed, R. Physical association of the catalytic and helper modules of a family-9 glycoside hydrolase is essential for activity. FEBS Lett. 2009, 583, 879–884. [Google Scholar] [CrossRef] [Green Version]
- Jervis, E.J.; Haynes, C.A.; Kilburn, D.G. Surface diffusion of cellulases and their isolated binding domains on cellulose. J. Biol. Chem. 1997, 272, 24016–24023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleine, J.; Liebl, W. Comparative characterization of deletion derivatives of the modular xylanase XynA of Thermotoga maritima. Extremophiles 2006, 10, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, K.; Sakka, M.; Kimura, T.; Sakka, K. Essential role of a family-32 carbohydrate-binding module in substrate recognition by Clostridium thermocellum mannanase CtMan5A. FEBS Lett. 2014, 588, 1726–1730. [Google Scholar] [CrossRef] [Green Version]
- Pasari, N.; Adlakha, N.; Gupta, M.; Bashir, Z.; Rajacharya, G.H.; Verma, G.; Munde, M.; Bhatnagar, R.; Yazdani, S.S. Impact of Module-X2 and Carbohydrate Binding Module-3 on the catalytic activity of associated glycoside hydrolases towards plant biomass. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Reyes-Ortiz, V.; Heins, R.A.; Cheng, G.; Kim, E.Y.; Vernon, B.C.; Elandt, R.B.; Adams, P.D.; Sale, K.L.; Hadi, M.Z.; Simmons, B.A. Addition of a carbohydrate-binding module enhances cellulase penetration into cellulose substrates. Biotechnol. Biofuels 2013, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Telke, A.A.; Ghatge, S.S.; Kang, S.-H.; Thangapandian, S.; Lee, K.-W.; Shin, H.-D.; Um, Y.; Kim, S.-W. Construction and characterization of chimeric cellulases with enhanced catalytic activity towards insoluble cellulosic substrates. Bioresour. Technol. 2012, 112, 10–17. [Google Scholar] [CrossRef]
- Thongekkaew, J.; Ikeda, H.; Masaki, K.; Iefuji, H. Fusion of cellulose binding domain from Trichoderma reesei CBHI to Cryptococcus sp. S-2 cellulase enhances its binding affinity and its cellulolytic activity to insoluble cellulosic substrates. Enzym. Microb. Technol. 2013, 52, 241–246. [Google Scholar] [CrossRef]
- Durrani, F.G.; Gul, R.; Mirza, M.U.; Kaderbhai, N.N.; Froeyen, M.; Saleem, M. Mutagenesis of DsbAss is Crucial for the Signal Recognition Particle Mechanism in Escherichia coli: Insights from Molecular Dynamics Simulations. Biomolecules 2019, 9, 133. [Google Scholar] [CrossRef] [Green Version]
- Jabbar, B.; Rafique, S.; Salo-Ahen, O.M.H.; Ali, A.; Munir, M.; Idrees, M.; Mirza, M.U.; Vanmeert, M.; Shah, S.Z.; Jabbar, I. Antigenic peptide prediction from E6 and E7 oncoproteins of HPV Types 16 and 18 for therapeutic vaccine design using immunoinformatics and MD simulation analysis. Front. Immunol. 2018, 9, 3000. [Google Scholar] [CrossRef] [Green Version]
- Grottesi, A.; Ceruso, M.A.; Colosimo, A.; Di Nola, A. Molecular dynamics study of a hyperthermophilic and a mesophilic rubredoxin. Proteins Struct. Funct. Bioinform. 2002, 46, 287–294. [Google Scholar] [CrossRef]
- Gruia, A.D.; Fischer, S.; Smith, J.C. Molecular dynamics simulation reveals a surface salt bridge forming a kinetic trap in unfolding of truncated Staphylococcal nuclease. Proteins Struct. Funct. Bioinform. 2003, 50, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Bowen, D.; Southerland, W.M. Barnase thermal titration via molecular dynamics simulations: Detection of early denaturation sites. J. Mol. Graph. Model. 2006, 24, 233–243. [Google Scholar] [CrossRef]
- Liu, J.; Yu, H.; Shen, Z. Insights into thermal stability of thermophilic nitrile hydratases by molecular dynamics simulation. J. Mol. Graph. Model. 2008, 27, 529–535. [Google Scholar] [CrossRef]
- Bae, E.; Phillips, G.N. Identifying and Engineering Ion Pairs in Adenylate Kinases Insights from molecular dynamics simulations of thermophilic and mesophilic homologues. J. Biol. Chem. 2005, 280, 30943–30948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Cruz, R.; Jäger, C.M.; Lau, P.L.; Gomes, R.L.; Pordea, A. Rational Design of Thermostable Carbonic Anhydrase Mutants Using Molecular Dynamics Simulations. J. Phys. Chem. B 2018, 122, 8526–8536. [Google Scholar] [CrossRef]
- Kundu, S.; Roy, D. Structural study of carboxylesterase from hyperthermophilic bacteria Geobacillus stearothermophilus by molecular dynamics simulation. J. Mol. Graph. Model. 2010, 28, 820–827. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, S.; Aurilia, V.; Marabotti, A.; Gonnelli, M.; Strambini, G. Structure and dynamics of cold-adapted enzymes as investigated by phosphorescence spectroscopy and molecular dynamics studies. 2. The case of an esterase from Pseudoalteromonas haloplanktis. J. Phys. Chem. B 2009, 113, 13171–13178. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, S.; Herman, P.; Lakowicz, J.R.; Bertoli, E.; Tanfani, F.; Rossi, M.; Manco, G. The thermophilic esterase from Archaeoglobus fulgidus: Structure and conformational dynamics at high temperature. Proteins Struct. Funct. Bioinform. 2000, 38, 351–360. [Google Scholar] [CrossRef]
- Wintrode, P.L.; Zhang, D.; Vaidehi, N.; Arnold, F.H.; Goddard, W.A., III. Protein dynamics in a family of laboratory evolved thermophilic enzymes. J. Mol. Biol. 2003, 327, 745–757. [Google Scholar] [CrossRef]
- Cao, R.; Jin, Y.; Xu, D. Recognition of cello-oligosaccharides by CBM17 from Clostridium cellulovorans: Molecular dynamics simulation. J. Phys. Chem. B 2012, 116, 6087–6096. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, C.; Xu, D. Molecular dynamics investigations of cello-oligosaccharide recognition by Cel9G–CBM3c from Clostridium cellulovorans. Phys. Chem. Chem. Phys. 2018, 20, 5235–5245. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Ahmad, S.; Asghar, R.; Sajjad, M.; Awan, M.T.; Akhtar, M.W. mRNA secondary structure engineering of Thermobifida fusca endoglucanase (cel6A) for enhanced expression in E. coli. World J. Microbiol. Biotechnol. 2015, 331, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Ghose, T.K. Measurements of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Khan, M.I.M.; Sajjad, M.; Ali, I.; Ahmad, S.; Akhtar, M.W. Influence of transposition and insertion of additional binding domain on expression and characteristics of xylanase C of Clostridium thermocellum. J. Biotechnol. 2010, 150, 1–5. [Google Scholar] [CrossRef]
- Sajjad, M.; Khan, M.I.M.; Akbar, N.S.; Ahmad, S.; Ali, I.; Akhtar, M.W. Enhanced expression and activity yields of Clostridium thermocellum xylanases without non-catalytic domains. J. Biotechnol. 2010, 145, 38–42. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Protein Structure Modeling with MODELLER; Protein Structure Prediction; Methods in Molecular Biology, Humana Press: New York, NY, USA, 2014; pp. 1–15. [Google Scholar]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Case, D.; Ben-Shalom, I.; Brozell, S.; Cerutti, D.; Cheatham, T., III; Cruzeiro, V.; Darden, T.; Duke, R.; Ghoreishi, D.; Gilson, M. AMBER 2018; University of California: San Francisco, CA, USA, 2018. [Google Scholar]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, D.J.; Brooks, C.L., III. A modified TIP3P water potential for simulation with Ewald summation. J. Chem. Phys. 2004, 121, 10096–10103. [Google Scholar] [CrossRef] [PubMed]
- Mirza, M.U.; Vanmeert, M.; Froeyen, M.; Ali, A.; Rafique, S.; Idrees, M. In silico structural elucidation of RNA-dependent RNA polymerase towards the identification of potential Crimean-Congo Hemorrhagic Fever Virus inhibitors. Sci. Rep. 2019, 9, 6809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, S.; Bonnefond, A.; Tamanini, F.; Mirza, M.U.; Manzoor, J.; Janjua, Q.M.; Din, S.M.; Gaitan, J.; Milochau, A.; Durand, E. Loss-of-function mutations in ADCY3 cause monogenic severe obesity. Nat. Genet. 2018, 50, 175. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A.M.; Bergfors, T.; Dultz, E.; Irwin, D.C.; Roos, A.; Driguez, H.; Wilson, D.B.; Jones, T.A. Crystal structure of Thermobifida fusca endoglucanase cel6A in complex with substrate and inhibitor: The role of tyrosine Y73 in substrate ring distortion. Biochemistry 2005, 44, 12915–12922. [Google Scholar] [CrossRef]
- Iman, K.; Mirza, M.U.; Mazhar, N.; Vanmeert, M.; Irshad, I.; Kamal, M.A. In silico Structure-based Identification of Novel Acetylcholinesterase Inhibitors Against Alzheimer’s Disease. CNS Neurol. Disord. Drug Targets Former. Curr. Drug Targets CNS Neurol. Disord. 2018, 17, 54–68. [Google Scholar] [CrossRef]
- Hou, T.; Wang, J.; Li, Y.; Wang, W. Assessing the performance of the molecular mechanics/Poisson Boltzmann surface area and molecular mechanics/generalized Born surface area methods. II. The accuracy of ranking poses generated from docking. J. Comput. Chem. 2011, 32, 866–877. [Google Scholar] [CrossRef] [Green Version]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W. Calculating structures and free energies of complex molecules: Combining molecular mechanics and continuum models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef]
- Sajjad, M.; Khan, M.I.M.; Zafar, R.; Ahmad, S.; Niazi, U.H.K.; Akhtar, M.W. Influence of positioning of carbohydrate binding module on the activity of endoglucanase CelA of Clostridium thermocellum. J. Biotechnol. 2012, 161, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.C.; Spezio, M.; Walker, L.P.; Wilson, D.B. Activity studies of eight purified cellulases: Specificity, synergism, and binding domain effects. Biotechnol. Bioeng. 1993, 42, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Kollman, P.A. Pathways to a protein folding intermediate observed in a 1-microsecond simulation in aqueous solution. Science 1998, 282, 740–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Wang, L.; Kollman, P.A. The early stage of folding of villin headpiece subdomain observed in a 200-nanosecond fully solvated molecular dynamics simulation. Proc. Natl. Acad. Sci. USA 1998, 95, 9897–9902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostylev, M. Studies of Thermobifida Fusca Cellulases Cel9A and Cel48A Using a Novel Kinetic Modeling Approach for the Enzymatic Digestion of Cellulose. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 2013. [Google Scholar]
- Purmonen, M.; Valjakka, J.; Takkinen, K.; Laitinen, T.; Rouvinen, J. Molecular dynamics studies on the thermostability of family 11 xylanases. Protein Eng. Des. Sel. 2007, 20, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liwo, A.; Czaplewski, C.; Ołdziej, S.; Scheraga, H.A. Computational techniques for efficient conformational sampling of proteins. Curr. Opin. Struct. Biol. 2008, 18, 134–139. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endoglucanase (cel6A) Variants | Cell Soluble Protein (mg l−1 OD600−1) | Expression Level of Variant in Soluble Cell Protein (%) | cel6A Yield (mg l−1 OD600−1) | Activity on 1% Carboxymethyl Cellulose (CMC) | Activity on Insoluble Substrates a (μmol Reducing Sugar μM−1 Enzyme) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Endoglu-Canase Activity a (U l−1 OD600−1) | Specific Activity (U mg−1 Enzyme) | Filter Paper (FP) | Bagasse | Regenerated Amorphous Cellulose (RAC) | Bacterial Microcrystalline Cellulose (BMCC) | ||||
| cel6A.C | 170 ± 8.7 | 45 ± 2.9 | 76.5 ± 3.9 | 320 ± 12.5 | 4.19 ± 0.05 | 0.7 ± 0.02 | 0.9 ± 0.06 | 1.3 ± 0.06 | 0.24 ± 0.03 |
| cel6A.BC | 168 ± 9.2 | 40 ± 3.5 | 67.2 ± 1.4 | 290 ± 10.2 | 4.32 ± 0.05 | 1.0 ± 0.05 | 1.4 ± 0.03 | 2.6 ± 0.12 | 0.28 ± 0.01 |
| cel6A.BCB | 190 ± 8.4 | 30 ± 2.1 | 57.0 ± 0.6 | 170 ± 9.9 | 2.98 ± 0.03 | 1.7 ± 0.09 | 2.2 ± 0.14 | 3.2 ± 0.18 | 0.60 ± 0.03 |
| cel6A.CBC | 185 ± 8.0 | 45 ± 3.0 | 83.3 ± 1.3 | 600 ± 14.8 | 7.20 ± 0.13 | 2.0 ± 0.11 | 3.4 ± 0.11 | 5.4 ± 0.25 | 0.90 ± 0.02 |
| cel6A.CBCB | 180 ± 10.3 | 20 ± 1.4 | 36.0 ± 0.3 | 250 ± 10.7 | 6.94 ± 0.31 | 1.5 ± 0.13 | 2.0 ± 0.10 | 1.7 ± 0.14 | 0.56 ± 0.09 |
| All-Atom Contacts | Summary Statistics | cel6A.CB (20 ns) | cel6A.CBC (20 ns) | ||
|---|---|---|---|---|---|
| Clash Score, All Atoms: | 0.94 | 1.13 | |||
| Protein Geometry | Poor rotamers | 8/407 | 1.96% | 16/689 | 2.32% |
| Favored rotamers | 372/407 | 91.4% | 632/689 | 91.72% | |
| Ramachandran outliers | 7/407 | 1.71% | 11/689 | 1.59% | |
| Ramachandran favored | 370/407 | 90.9% | 622/689 | 90.27% | |
| Ramachandran allowed | 394/407 | 96.8% | 674/689 | 97.9% | |
| MolProbity score | 1.70 | 1.68 | |||
| Cβ deviations >0.25Å | 23 | 5.65% | 51 | 7.4% | |
| Bad bonds: | 30/2194 | 1.37% | 62/5038 | 1.23% | |
| Bad angles: | 74/3014 | 2.45% | 105/6939 | 1.51% | |
| Peptide Omegas | Cis Prolines: | 0/20 | 0.00% | 0/52 | 0.00% |
| Cis nonProlines: | 1 | 0.24% | 3 | 0.42% | |
| Contributions | cel6A.CB (kcal/mol) | cel6A.CBC (kcal/mol) | |||
|---|---|---|---|---|---|
| 26.85 °C | 60.85 °C | 74.85 °C | 126.85 °C | ||
| ΔEele | −5.55 | −8.94 | −7.85 | −8.24 | −2.15 |
| ΔEvdw | −14.57 | −23.87 | −22.84 | −20.26 | −13.2 |
| ΔEMM | −20.12 | −32.81 | −30.69 | −28.5 | −15.35 |
| ΔGp | 10.2 | 15.63 | 13.96 | 13.45 | 7.98 |
| ΔGnp | −2.09 | −3.45 | −4.82 | −4.21 | −1.62 |
| ΔGsol | 8.11 | 12.18 | 9.14 | 9.24 | 6.36 |
| ΔGtol | −12.01 | −20.63 | −21.55 | −19.26 | −8.99 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, I.; Rehman, H.M.; Mirza, M.U.; Akhtar, M.W.; Asghar, R.; Tariq, M.; Ahmed, R.; Tanveer, F.; Khalid, H.; Alghamdi, H.A.; et al. Enhanced Thermostability and Enzymatic Activity of cel6A Variants from Thermobifida fusca by Empirical Domain Engineering. Biology 2020, 9, 214. https://doi.org/10.3390/biology9080214
Ali I, Rehman HM, Mirza MU, Akhtar MW, Asghar R, Tariq M, Ahmed R, Tanveer F, Khalid H, Alghamdi HA, et al. Enhanced Thermostability and Enzymatic Activity of cel6A Variants from Thermobifida fusca by Empirical Domain Engineering. Biology. 2020; 9(8):214. https://doi.org/10.3390/biology9080214
Chicago/Turabian StyleAli, Imran, Hafiz Muzzammel Rehman, Muhammad Usman Mirza, Muhammad Waheed Akhtar, Rehana Asghar, Muhammad Tariq, Rashid Ahmed, Fatima Tanveer, Hina Khalid, Huda Ahmed Alghamdi, and et al. 2020. "Enhanced Thermostability and Enzymatic Activity of cel6A Variants from Thermobifida fusca by Empirical Domain Engineering" Biology 9, no. 8: 214. https://doi.org/10.3390/biology9080214
APA StyleAli, I., Rehman, H. M., Mirza, M. U., Akhtar, M. W., Asghar, R., Tariq, M., Ahmed, R., Tanveer, F., Khalid, H., Alghamdi, H. A., & Froeyen, M. (2020). Enhanced Thermostability and Enzymatic Activity of cel6A Variants from Thermobifida fusca by Empirical Domain Engineering. Biology, 9(8), 214. https://doi.org/10.3390/biology9080214