Characterization of Perionyx excavatus Development and Its Head Regeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
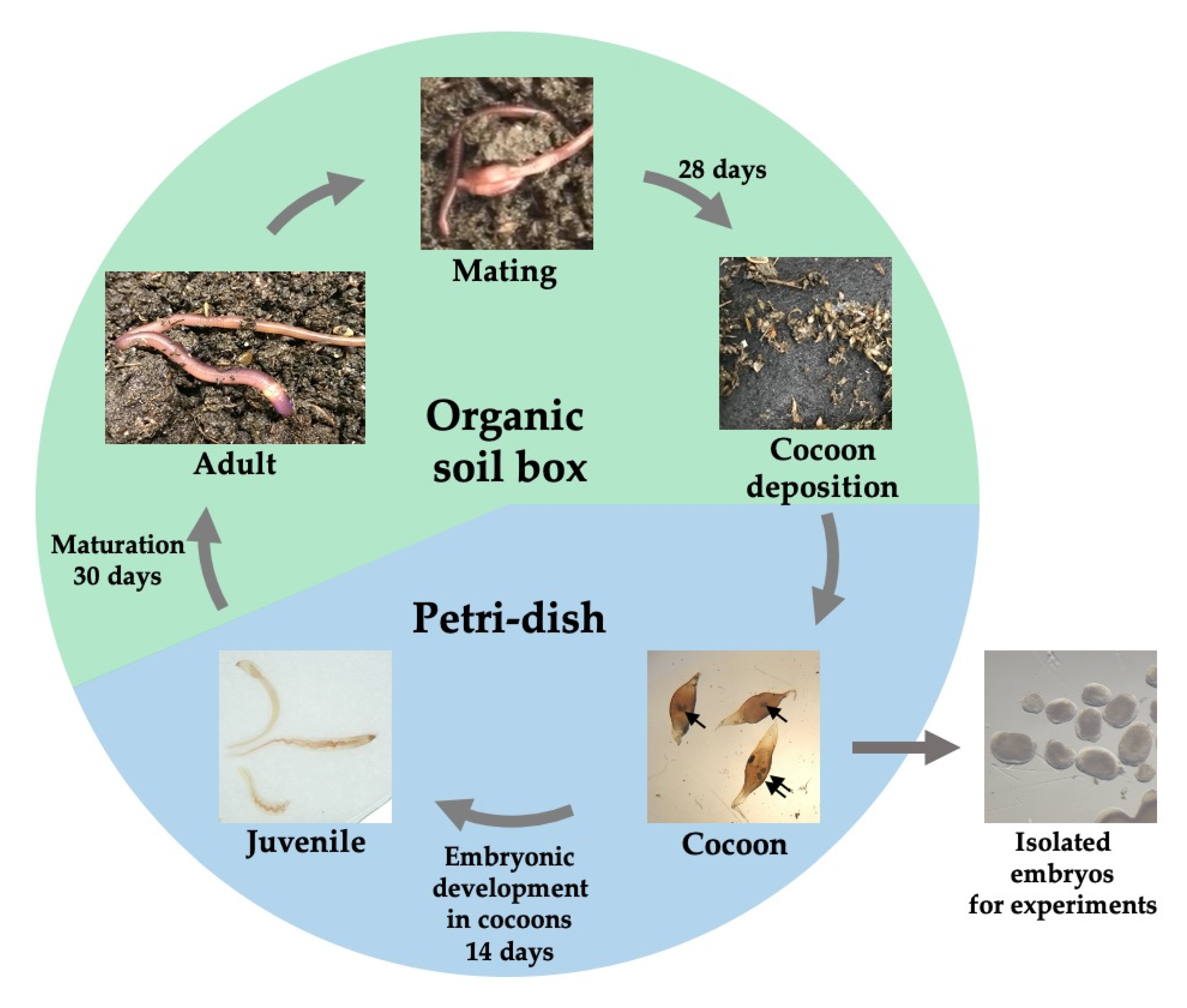
2.1. Cultivation of P. excavatus
2.2. Embryo and Juvenile Preparation
2.3. Regeneration of Juvenile Head
2.4. Sample Preparation for Imaging
2.5. EdU Assay for Cell Proliferation
2.6. Phalloidin Staining and Immunohistochemistry
2.7. Microscopy
3. Results and Discussions
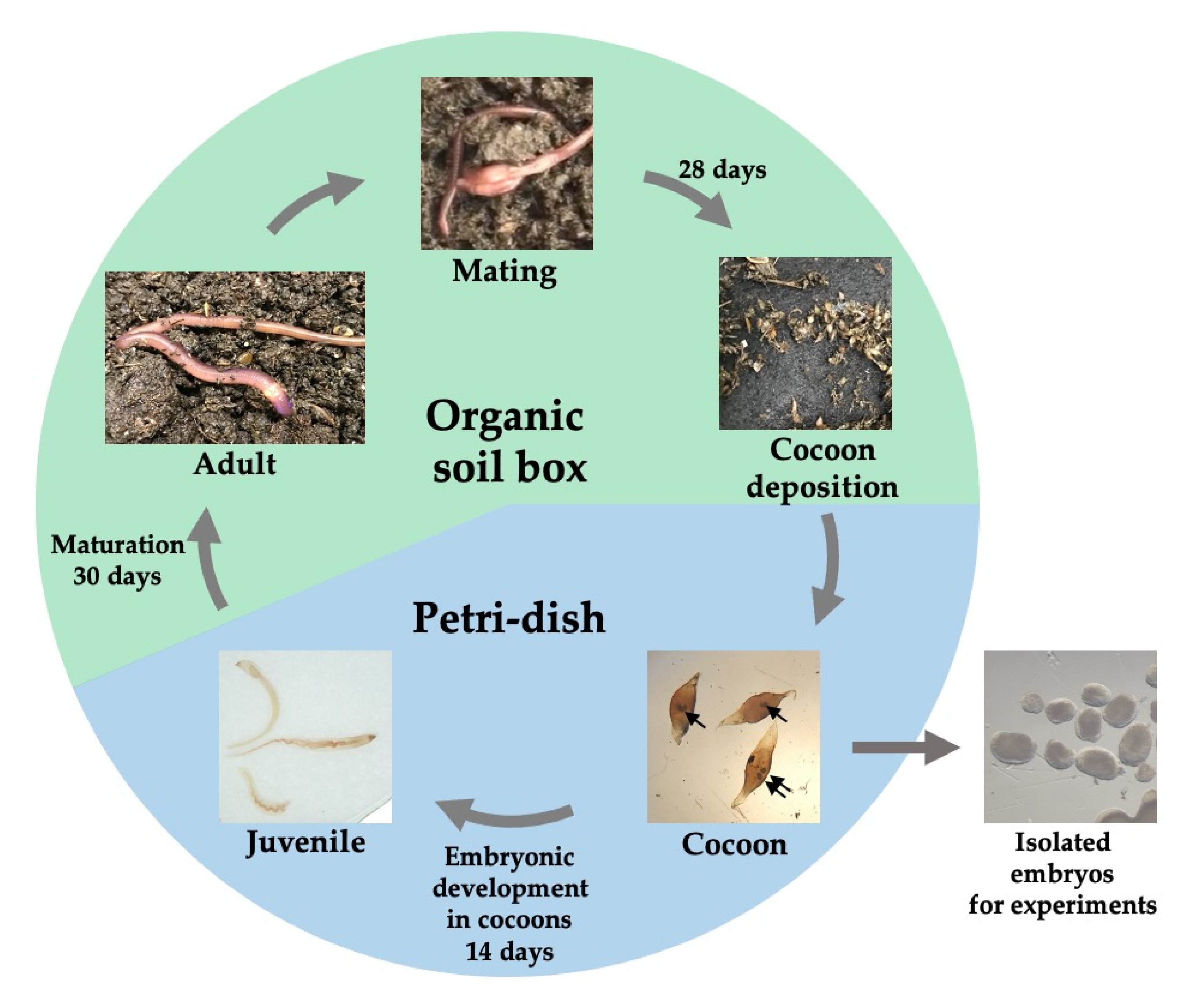
3.1. The Life Cycle of Perionyx excavatus
3.2. Characterization of Embryonic Development of Perionyx excavatus
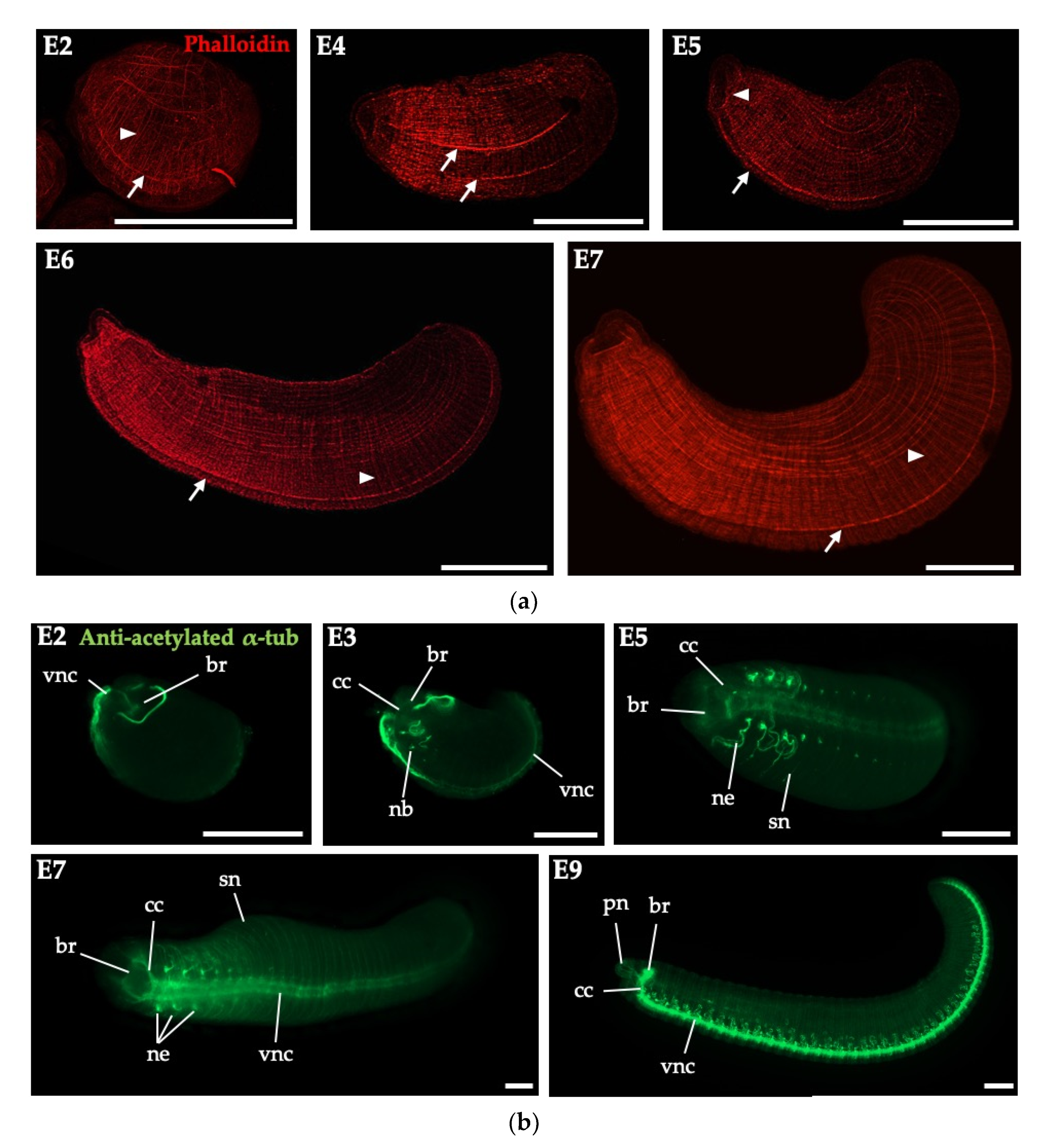
3.3. Embryonic Myogenesis and Neurogenesis of Perionyx excavatus
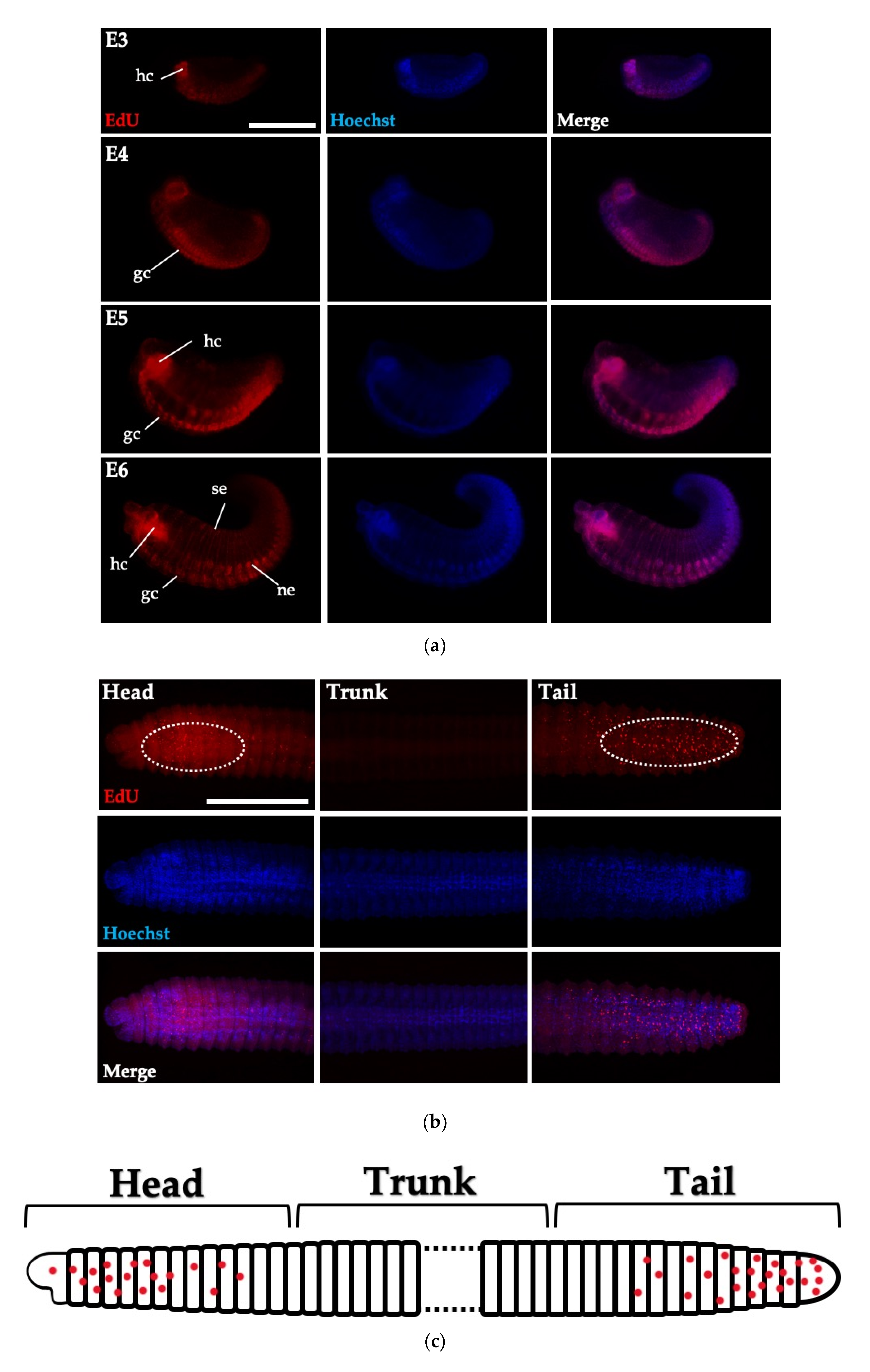
3.4. Cell Proliferation in Embryos and Juveniles of Perionyx excavatus
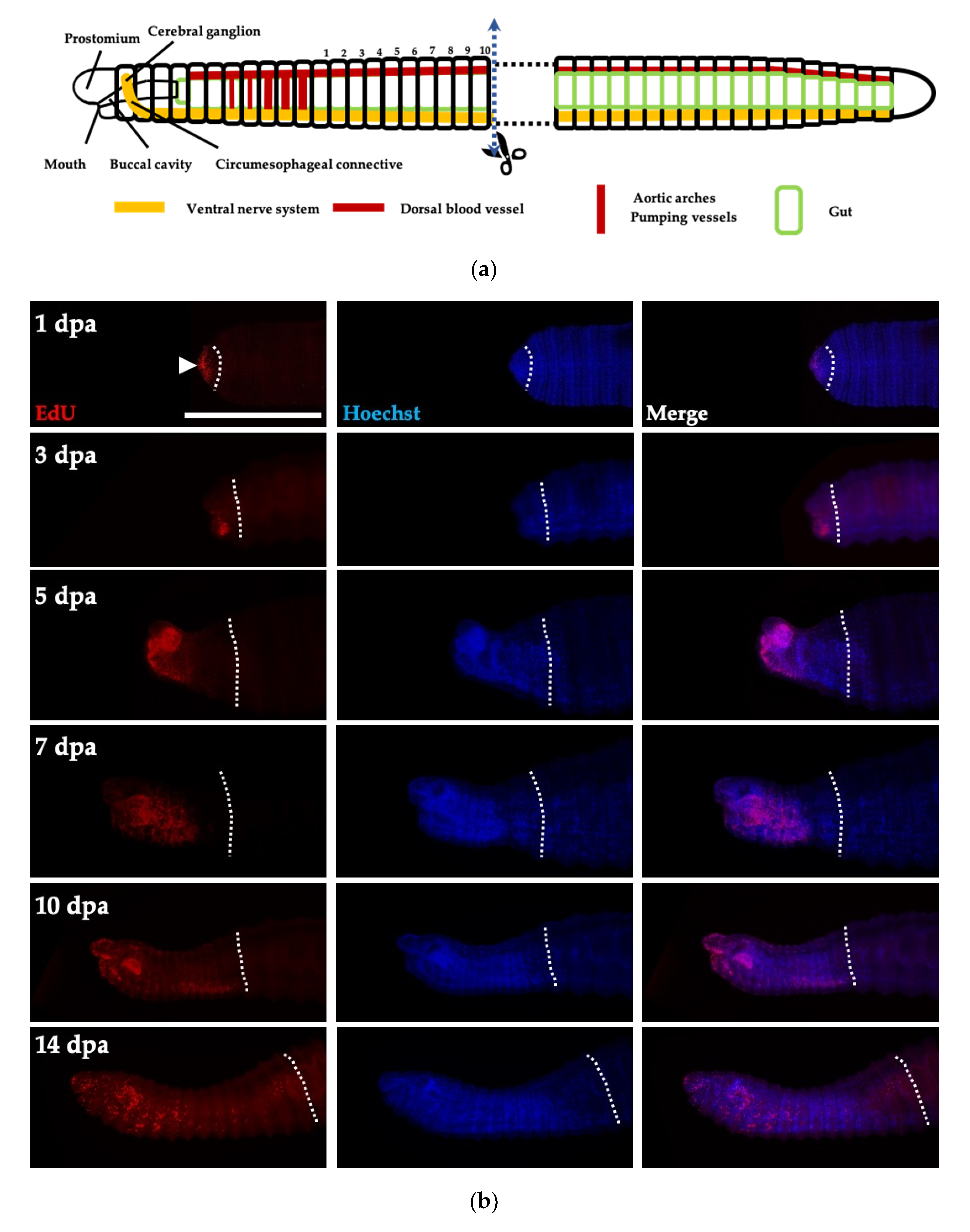
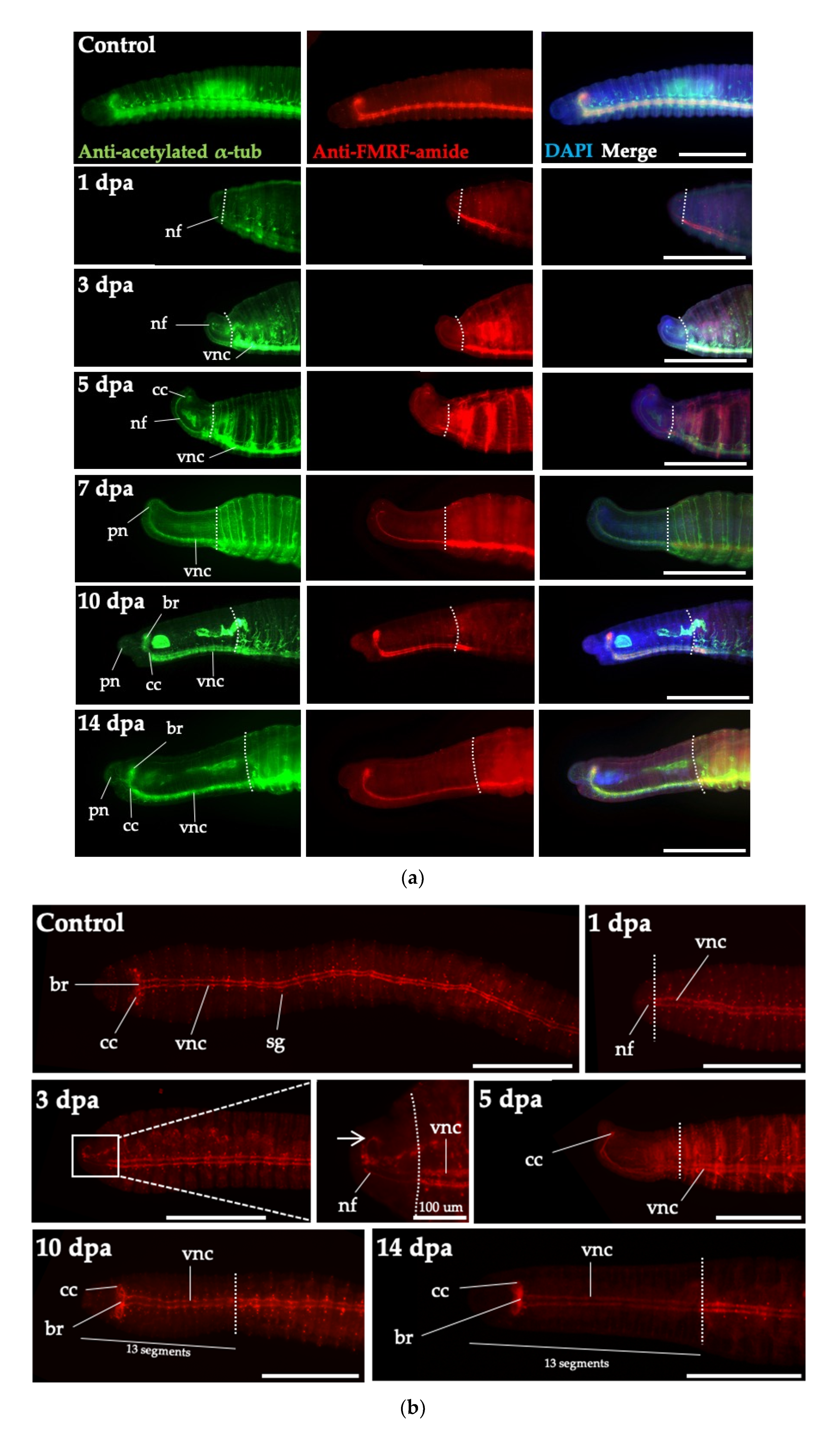
3.5. Cell Proliferation in Regenerative Tissues of Perionyx excavatus
3.6. Head Regeneration of Juveniles of Perionyx excavatus
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, J.M.; Ross, K.G.; Zayas, R.M. Regeneration in Invertebrates: Model Systems. In eLS; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 1–9. [Google Scholar] [CrossRef]
- Agata, K.; Saito, Y.; Nakajima, E. Unifying principles of regeneration I: Epimorphosis versus morphallaxis. Dev. Growth Differ. 2007, 49, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Özpolat, B.D.; Bely, A.E. Developmental and molecular biology of annelid regeneration: A comparative review of recent studies. Curr. Opin. Genet. Dev. 2016, 40, 144–153. [Google Scholar] [CrossRef]
- Ribeiro, R.P.; Bleidorn, C.; Aguado, M.T. Regeneration mechanisms in Syllidae (Annelida). Regeneration 2018, 5, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, M.E. Regeneration: Thomas Hunt Morgan’s Window into Development. J. Hist. Biol. 2019, 43, 325–361. [Google Scholar] [CrossRef] [PubMed]
- Zoran, M.J. Regeneration in Annelids. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 1–7. [Google Scholar] [CrossRef]
- Holstein, T.W.; Hobmayer, E.; David, C.N. Pattern of epithelial cell cycling in Hydra. Dev. Biol. 1991, 148, 602–611. [Google Scholar] [CrossRef] [Green Version]
- Newmark, P.A.; Sánchez Alvarado, A. Bromodeoxyuridine Specifically Labels the Regenerative Stem Cells of Planarians. Dev. Biol. 2000, 220, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleo, G.; Brown, C.W.; Laforest, L.; Akimenko, M.-A. Cell proliferation and movement during early fin regeneration in zebrafish. Dev. Dyn. 2001, 221, 380–390. [Google Scholar] [CrossRef]
- Roy, S.; Lévesque, M. Limb Regeneration in Axolotl: Is it superhealing? Sci. World J. 2006, 6, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Joven, A.; Elewa, A.; Simon, A. Model systems for regeneration: Salamanders. Development 2019, 146, dev167700. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-J.; Lee, M.S.; Tak, E.S.; Lee, E.; Koh, K.S.; Ahn, C.H.; Park, S.C. Gene Expression Profile in the Anterior Regeneration of the Earthworm Using Expressed Sequence Tags. Biosci. Biotechnol. Biochem. 2019, 73, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-J.; Koh, K.S.; Lee, E.; Park, S.C. Differential Expression of Three labial Genes during Earthworm Head Regeneration. Biosci. Biotechnol. Biochem. 2009, 73, 2609–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallatt, L.; Reinecke, A.J.; Viljoen, S.A. Life Cycle of the Oriental Compost Worm Perionyx excavatus(Oligochaeta). S. Afr. J. Zool. 1990, 25, 41–45. [Google Scholar] [CrossRef]
- Hunnekuhl, V.S.; Bergter, A.; Purschke, G.; Paululat, A. Development and embryonic pattern of body wall musculature in the crassiclitellate Eisenia andrei(Annelida, Clitellata). J. Morphol. 2009, 270, 1122–1136. [Google Scholar] [CrossRef] [PubMed]
- Koza, A.; Wilhelm, M.; Hiripi, L.; Elekes, K.; Csoknya, M. Embryogenesis of the serotonergic system in the earthworm Eisenia fetida (Annelida, Oligochaeta): Immunohistochemical and biochemical studies. J. Comp. Neurol. 2006, 497, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Myohara, M. Differential tissue development during embryogenesis and regeneration in an annelid. Dev. Dyn. 2004, 231, 349–358. [Google Scholar] [CrossRef]
- Bergter, A.; Paululat, A. Pattern of body-wall muscle differentiation during embryonic development of Enchytraeus coronatus (Annelida: Oligochaeta; Enchytraeidae). J. Morphol. 2007, 268, 537–549. [Google Scholar] [CrossRef]
- Boros, A.; Reglodi, D.; Herbert, Z.; Kiszler, G.; Nemeth, J.; Lubics, A.; Molnar, L. Changes in the Expression of PACAP-like Compounds During the Embryonic Development of the Earthworm Eisenia fetida. J. Mol. Neurosci. 2008, 36, 157–165. [Google Scholar] [CrossRef]
- Boros, Á.; Somogyi, I.; Engelmann, P.; Lubics, A.; Reglodi, D.; Pollák, E.; Molnár, L. Pituitary adenylate cyclase-activating polypeptide type 1 (PAC1) receptor is expressed during embryonic development of the earthworm. Cell Tissue Res. 2010, 339, 649–653. [Google Scholar] [CrossRef]
- Prosser, C.L. Correlation between development of behavior and neuromuscular differentiation in embryos of Eisenia foetida, Sav. J. Comp. Neurol. 1933, 58, 603–641. [Google Scholar] [CrossRef]
- Seaver, E.C.; Thamm, K.; Hill, S.D. Growth patterns during segmentation in the two polychaete annelids, Capitella sp. I and Hydroides elegans: Comparisons at distinct life history stages. Evol. Dev. 2005, 7, 312–326. [Google Scholar] [CrossRef]
- Myohara, M. What Role Do Annelid Neoblasts Play? A Comparison of the Regeneration Patterns in a Neoblast-Bearing and a Neoblast-Lacking Enchytraeid Oligochaete. PLoS ONE 2012, 7, e37319. [Google Scholar] [CrossRef] [PubMed]
- De Jong, D.M.; Seaver, E.C. A Stable Thoracic Hox Code and Epimorphosis Characterize Posterior Regeneration in Capitella teleta. PLoS ONE 2016, 11, e0149724. [Google Scholar] [CrossRef] [Green Version]
- Myohara, M.; Yoshida-Noro, C.; Kobari, F.; Tochinai, S. Fragmenting oligochaete Enchytraeus japonensis: A new material for regeneration study. Dev. Growth Differ. 1999, 41, 549–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zattara, E.E.; Bely, A.E. Evolution of a novel developmental trajectory: Fission is distinct from regeneration in the annelid Pristina leidyi. Evol. Dev. 2011, 13, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Zattara, E.E.; Bely, A.E. Investment choices in post-embryonic development: Quantifying interactions among growth, regeneration, and asexual reproduction in the annelid Pristina leidyi. J. Exp. Zool. Part B Mol. Dev. Evol. 2013, 320, 471–488. [Google Scholar] [CrossRef]
- Muller, M.C.M. Nerve development, growth and differentiation during regeneration in Enchytraeus fragmentosus and Stylaria lacustris (Oligochaeta). Dev. Growth Differ. 2004, 46, 471–478. [Google Scholar] [CrossRef]
- Meyer, N.P.; Seaver, E.C. Neurogenesis in an annelid: Characterization of brain neural precursors in the polychaete Capitella sp. I. Dev. Biol. 2009, 335, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Weidhase, M.; Helm, C.; Bleidorn, C. Morphological investigations of posttraumatic regeneration in Timarete cf. punctata (Annelida: Cirratulidae). Zool. Lett. 2015, 1, 20. [Google Scholar] [CrossRef] [Green Version]
- Kozin, V.V.; Filippova, N.A.; Kostyuchenko, R.P. Regeneration of the nervous and muscular system after caudal amputation in the polychaete Alitta virens (Annelida: Nereididae). Russ. J. Dev. Biol. 2017, 48, 198–210. [Google Scholar] [CrossRef]
- Weidhase, M.; Beckers, P.; Bleidorn, C.; Aguado, M.T. Nervous system regeneration in Typosyllis antoni (Annelida: Syllidae). Zool. Anz. 2017, 269, 57–67. [Google Scholar] [CrossRef]
- Starunov, V.V.; Voronezhskaya, E.E.; Nezlin, L.P. Development of the nervous system in Platynereis dumerilii (Nereididae, Annelida). Front. Zool. 2017, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.G.; Menger, G.J., III; Zoran, M.J. Regeneration and asexual reproduction share common molecular changes: Upregulation of a neural glycoepitope during morphallaxis in Lumbriculus. Mech. Dev. 2005, 122, 721–732. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, Y.S.; Kim, J.; Yi, J.; Park, S.C.; Lee, H.-Y.; Cho, S.-J. Characterization of Perionyx excavatus Development and Its Head Regeneration. Biology 2020, 9, 273. https://doi.org/10.3390/biology9090273
Bae YS, Kim J, Yi J, Park SC, Lee H-Y, Cho S-J. Characterization of Perionyx excavatus Development and Its Head Regeneration. Biology. 2020; 9(9):273. https://doi.org/10.3390/biology9090273
Chicago/Turabian StyleBae, Yun Seon, Jung Kim, Jeesoo Yi, Soon Cheol Park, Hae-Youn Lee, and Sung-Jin Cho. 2020. "Characterization of Perionyx excavatus Development and Its Head Regeneration" Biology 9, no. 9: 273. https://doi.org/10.3390/biology9090273
APA StyleBae, Y. S., Kim, J., Yi, J., Park, S. C., Lee, H. -Y., & Cho, S. -J. (2020). Characterization of Perionyx excavatus Development and Its Head Regeneration. Biology, 9(9), 273. https://doi.org/10.3390/biology9090273