

Evaluation of Inflammatory Cellular Model by Advanced Bioanalytic and Artificial Intelligence Analyses of Lipids: Lipidomic Landscape of Inflammaging






Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture and Treatment
2.3. Sample Preparation
2.4. Lipid Extraction by MTBE
2.5. Mass Spectrometry for Untargeted Analysis of Lipidome
2.6. Statistical and Networking Data Analysis
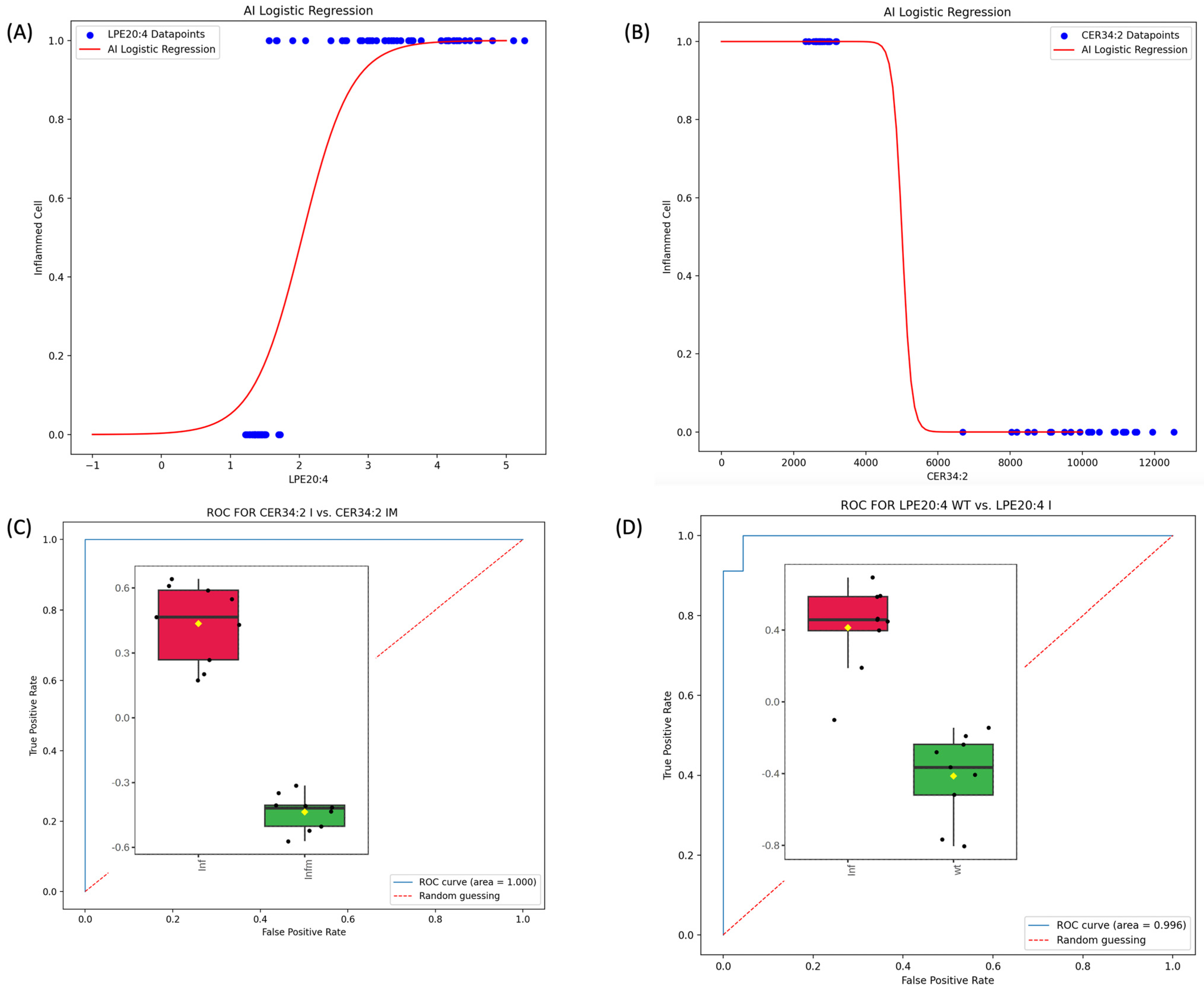
2.7. AI, ML, Monte Carlo, and ROC Analyses
3. Results and Discussion
3.1. Setup of Analytical Method of Lipid Analysis
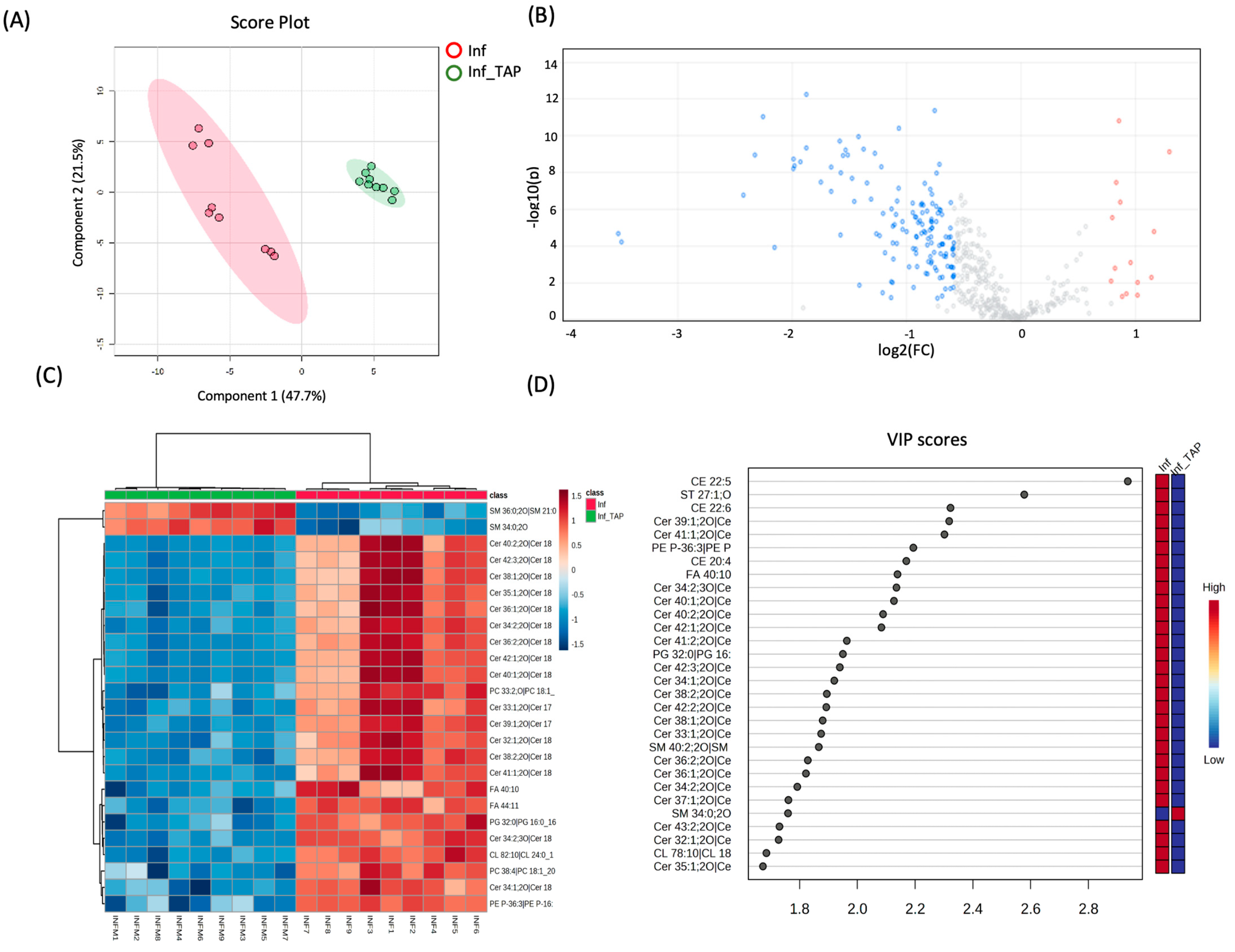
3.2. Effect of Induced Inflammation on R3/1 NF-κB Reporter Cells: Study of Lipidome of Cellular Model
3.3. Effect of Thinned Apple Polyphenols (TAPs) on Inflamed R3/1 NF-κB Reporter Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ceramide Species | Inf_TAP vs. Inf | |
|---|---|---|
| Log2Ratio | References | |
| Cer 32:1;2O| Cer(d18:1/14:0) | −1.28 | [27] |
| Cer 33:1;2O| Cer(d17:1/16:0) | −1.52 | |
| Cer 34:1;2O| Cer(d18:1/16:0) | −1.56 | [28,29] |
| Cer 34:2;2O| Cer(d18:2/16:0) | −1.38 | |
| Cer 34:2;3O| Cer(d18:2/16:0(2OH)) | −1.88 | |
| Cer 35:1;2O| Cer(d18:1/17:0) | −1.22 | |
| Cer 36:1;2O| Cer(d18:1/18:0) | −1.48 | [28,29] |
| Cer 36:2;2O| Cer(d18:2/18:0) | −1.42 | |
| Cer 37:1;2O| Cer(d18:1/19:0) | −1.46 | |
| Cer 38:0;2O| Cer(d18:0/20:0) | −1.09 | |
| Cer 38:1;2O| Cer(d18:1/20:0) | −1.58 | |
| Cer 38:2;2O| Cer(d18:2/20:0) | −1.53 | |
| Cer 39:1;2O| Cer(d17:1/22:0) | −2.26 | |
| Cer 40:0;2O| Cer(d18:0/22:0) | −1.08 | |
| Cer 40:1;2O| Cer(d18:1/22:0) | −1.99 | |
| Cer 40:2;2O| Cer(d18:2/22:0) | −1.93 | |
| Cer 41:1;2O| Cer(d18:1/23:0) | −2.33 | |
| Cer 41:2;2O| Cer(d18:1/23:1) | −1.75 | |
| Cer 42:1;2O| Cer(d18:0/24:1) | −1.11 | |
| Cer 42:1;2O| Cer(d18:1/24:0) | −1.88 | [28,29] |
| Cer 42:1;3O| Cer(d18:0/24:1(2OH)) | −0.80 | [28,29] |
| Cer 42:2;2O| Cer(d18:1/24:1) | −1.66 | |
| Cer 42:3;2O| Cer(d18:2/24:1) | −1.66 | |
| Cer 43:2;2O| Cer(d18:1/25:1) | −1.35 |
3.4. Construction of ML Diagnostic Model for Inf vs. Wt and Inf_TAP vs. Inf
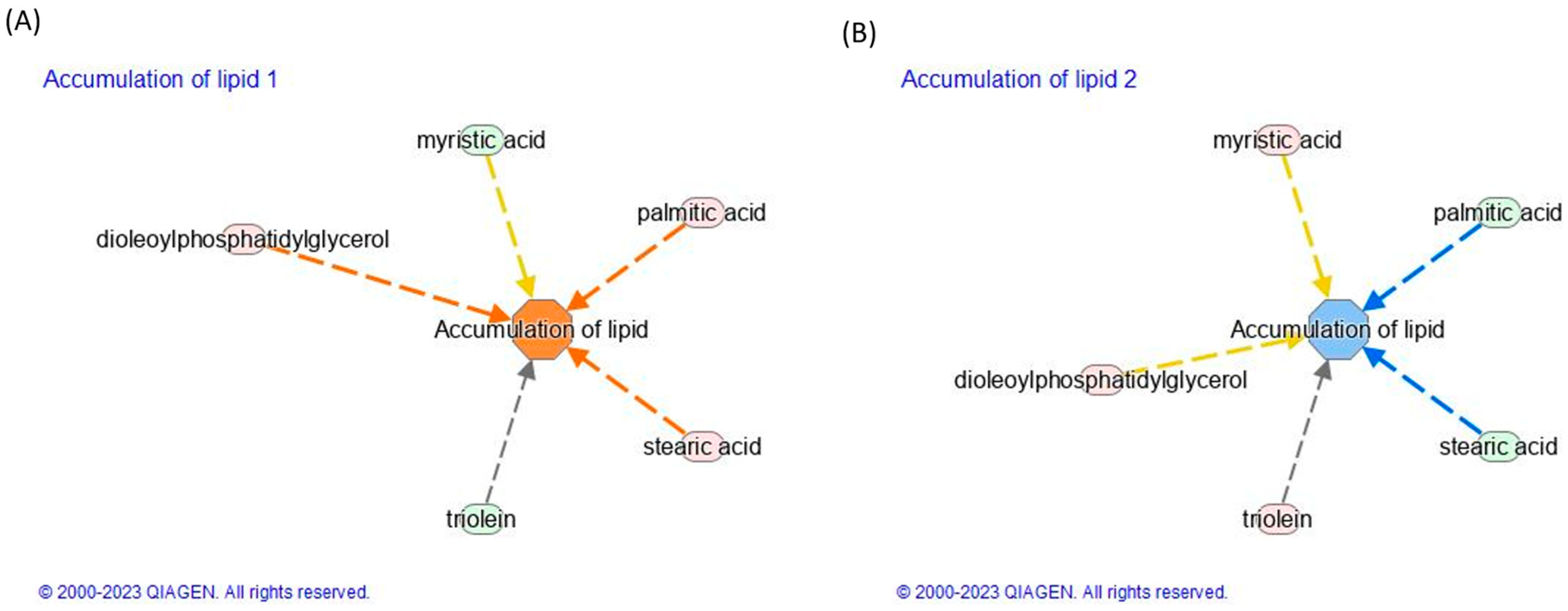
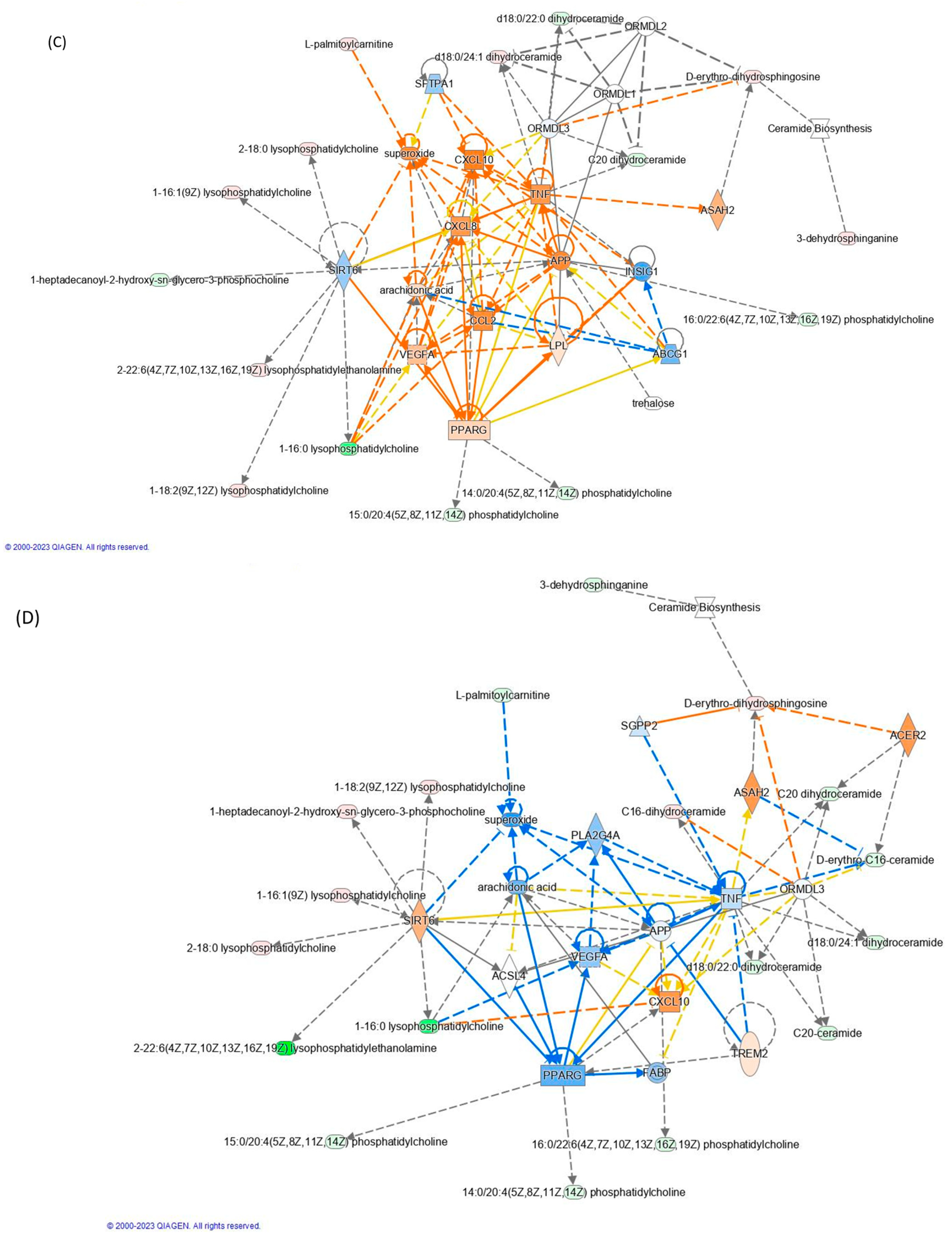
3.5. Effect of TNF-α and TAPs on Signaling Pathway Modulation by Network Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barcia, A.M.; Harris, H.W. Triglyceride-rich lipoproteins as agents of innate immunity. Clin. Infect. Dis. 2005, 41, S498–S503. [Google Scholar] [CrossRef]
- Khovidhunkit, W.; Kim, M.S.; Memon, R.A.; Shigenaga, J.K.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Effects of infection and inflammation on lipid and lipoprotein metabolism: Mechanisms and consequences to the host. J. Lipid Res. 2004, 45, 1169–1196. [Google Scholar] [CrossRef]
- Doyle, R.; Sadlier, D.M.; Godson, C. Pro-resolving lipid mediators: Agents of anti-ageing? Semin. Immunol. 2018, 40, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Sen, P.; Lamichhane, S.; Mathema, V.B.; McGlinchey, A.; Dickens, A.M.; Khoomrung, S.; Oresic, M. Deep learning meets metabolomics: A methodological perspective. Brief. Bioinform. 2021, 22, 1531–1542. [Google Scholar] [CrossRef]
- Tosi, D.; Campi, A. How Data Analytics and Big Data Can Help Scientists in Managing COVID-19 Diffusion: Modeling Study to Predict the COVID-19 Diffusion in Italy and the Lombardy Region. J. Med. Internet Res. 2020, 22, e21081. [Google Scholar] [CrossRef]
- Jin, Y.N.; Arroo, R. The protective effects of flavonoids and carotenoids against diabetic complications-A review of in vivo evidence. Front. Nutr. 2023, 10, 1020950. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, G.; Baron, G.; Gado, F.; Della Vedova, L.; Bombardelli, E.; Carini, M.; D’Amato, A.; Aldini, G.; Altomare, A. Polyphenols from Thinned Young Apples: HPLC-HRMS Profile and Evaluation of Their Anti-Oxidant and Anti-Inflammatory Activities by Proteomic Studies. Antioxidants 2022, 11, 1577. [Google Scholar] [CrossRef] [PubMed]
- Dei Cas, M.; Ghidoni, R. Cancer Prevention and Therapy with Polyphenols: Sphingolipid-Mediated Mechanisms. Nutrients 2018, 10, 940. [Google Scholar] [CrossRef]
- Ma, J.; Zheng, Y.M.; Tang, W.J.; Yan, W.X.; Nie, H.F.; Fang, J.; Liu, G. Dietary polyphenols in lipid metabolism: A role of gut microbiome. Anim. Nutr. 2020, 6, 404–409. [Google Scholar] [CrossRef]
- Rahn, C.; Bakuradze, T.; Stegmuller, S.; Galan, J.; Niesen, S.; Winterhalter, P.; Richling, E. Polyphenol-Rich Beverage Consumption Affecting Parameters of the Lipid Metabolism in Healthy Subjects. Int. J. Mol. Sci. 2023, 24, 841. [Google Scholar] [CrossRef]
- Baron, G.; Altomare, A.; Mol, M.; Garcia, J.L.; Correa, C.; Raucci, A.; Mancinelli, L.; Mazzotta, S.; Fumagalli, L.; Trunfio, G.; et al. Analytical Profile and Antioxidant and Anti-Inflammatory Activities of the Enriched Polyphenol Fractions Isolated from Bergamot Fruit and Leave. Antioxidants 2021, 10, 141. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Xia, J.G.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef]
- Zweig, M.H.; Campbell, G. Receiver-Operating Characteristic (Roc) Plots—A Fundamental Evaluation Tool in Clinical Medicine. Clin. Chem. 1993, 39, 561–577. [Google Scholar] [CrossRef]
- Dominguez-Almendros, S.; Benitez-Parejo, N.; Gonzalez-Ramirez, A.R. Logistic Regression Models. Allergol. Immunopathol. 2011, 39, 295–305. [Google Scholar] [CrossRef]
- Shimizu, T. Lipid Mediators in Health and Disease: Enzymes and Receptors as Therapeutic Targets for the Regulation of Immunity and Inflammation. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 123–150. [Google Scholar] [CrossRef]
- Chang, M.C.; Lee, J.J.; Chen, Y.J.; Lin, S.I.; Lin, L.D.; Liou, E.J.W.; Huang, W.L.; Chan, C.P.; Huang, C.C.; Jeng, J.H. Lysophosphatidylcholine induces cytotoxicity/apoptosis and IL-8 production of human endothelial cells: Related mechanisms. Oncotarget 2017, 8, 106177–106189. [Google Scholar] [CrossRef] [PubMed]
- Brkic, L.; Riederer, M.; Graier, W.F.; Malli, R.; Frank, S. Acyl chain-dependent effect of lysophosphatidylcholine on cyclooxygenase (COX)-2 expression in endothelial cells. Atherosclerosis 2012, 224, 348–354. [Google Scholar] [CrossRef]
- Liu, P.P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef]
- Joffre, C.; Rey, C.; Laye, S. N-3 Polyunsaturated Fatty Acids and the Resolution of Neuroinflammation. Front. Pharmacol. 2019, 10, 1022. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.S.; McMahon, E.J.; Pop, S.M.; Reap, E.A.; Caricchio, R.; Cohen, P.L.; Earp, H.S.; Matsushima, G.K. Phagocytosis and clearance of apoptotic cells is mediated by MER. Nature 2001, 411, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Munoz, A.; Presa, N.; Gomez-Larrauri, A.; Rivera, I.G.; Trueba, M.; Ordonez, M. Control of inflammatory responses by ceramide, sphingosine 1-phosphate and ceramide 1-phosphate. Prog. Lipid Res. 2016, 61, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Serna, I.M.R.; Sitina, M.; Stokin, G.B.; Medina-Inojosa, J.R.; Lopez-Jimenez, F.; Gonzalez-Rivas, J.P.; Vinciguerra, M. Lipidomic Profiling Identifies Signatures of Poor Cardiovascular Health. Metabolites 2021, 11, 747. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Pang, J.; Zhang, Y.; Zhang, H.Y.; Xu, Z.L.; Chen, Q.; Ling, W.H. Associations between plasma ceramides and mortality in patients with coronary artery disease. Atherosclerosis 2020, 314, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, R.; Ekroos, K.; Sysi-Aho, M.; Hilvo, M.; Vihervaara, T.; Kauhanen, D.; Suoniemi, M.; Hurme, R.; März, W.; Scharnagl, H.; et al. Plasma ceramides predict cardiovascular death in patients with stable coronary artery disease and acute coronary syndromes beyond LDL-cholesterol. Eur. Heart J. 2016, 37, 1967–1976. [Google Scholar] [CrossRef]
- Scheiblich, H.; Schlutter, A.; Golenbock, D.T.; Latz, E.; Martinez-Martinez, P.; Heneka, M.T. Activation of the NLRP3 inflammasome in microglia: The role of ceramide. J. Neurochem. 2017, 143, 534–550. [Google Scholar] [CrossRef]
- Field, B.C.; Gordillo, R.; Scherer, P.E. The Role of Ceramides in Diabetes and Cardiovascular Disease Regulation of Ceramides by Adipokines. Front. Endocrinol. 2020, 11, 569250. [Google Scholar] [CrossRef] [PubMed]
- Havulinna, A.S.; Sysi-Aho, M.; Hilvo, M.; Kauhanen, D.; Hurme, R.; Ekroos, K.; Salomaa, V.; Laaksonen, R. Circulating Ceramides Predict Cardiovascular Outcomes in the Population-Based FINRISK 2002 Cohort. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2424–2430. [Google Scholar] [CrossRef]
- Gaggini, M.; Ndreu, R.; Michelucci, E.; Rocchiccioli, S.; Vassalle, C. Ceramides as Mediators of Oxidative Stress and Inflammation in Cardiometabolic Disease. Int. J. Mol. Sci. 2022, 23, 2719. [Google Scholar] [CrossRef] [PubMed]
- Fretts, A.M.; Jensen, P.N.; Hoofnagle, A.N.; McKnight, B.; Howard, B.V.; Umans, J.; Sitlani, C.M.; Siscovick, D.S.; King, I.B.; Djousse, L.; et al. Plasma ceramides containing saturated fatty acids are associated with risk of type 2 diabetes. J. Lipid Res. 2021, 62, 100119. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Tveter, K.M.; Villa-Rodriguez, J.A.; Cabales, A.J.; Zhang, L.; Bawagan, F.G.; Duran, R.M.; Roopchand, D.E. Polyphenol-induced improvements in glucose metabolism are associated with bile acid signaling to intestinal farnesoid X receptor. BMJ Open Diabetes Res. Care 2020, 8, e001386. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morato, J.; Boronat, A.; Serreli, G.; Enriquez, L.; Gomez-Gomez, A.; Pozo, O.J.; Fito, M.; de la Torre, R. Effects of Wine and Tyrosol on the Lipid Metabolic Profile of Subjects at Risk of Cardiovascular Disease: Potential Cardioprotective Role of Ceramides. Antioxidants 2021, 10, 1679. [Google Scholar] [CrossRef]
- Karin, N.; Razon, H. Chemokines beyond chemo-attraction: CXCL10 and its significant role in cancer and autoimmunity. Cytokine 2018, 109, 24–28. [Google Scholar] [CrossRef]

| Number of Lipids | % | Number of Lipids | % | ||
|---|---|---|---|---|---|
| Identified Features in MS-DIAL | 469 | Identified Features in MS-DIAL | |||
| PC | 94 | 20.2 | |||
| PE | 96 | 20.2 | LPE | 10 | 2.0 |
| SM | 47 | 10.1 | HexCer | 9 | 1.8 |
| PI | 28 | 6.9 | CAR | 3 | 0.6 |
| Cer | 3 | 6.3 | CE | 3 | 0.6 |
| PG | 26 | 5.3 | NAE | 5 | 1.0 |
| PS | 24 | 5.1 | MG | 2 | 0.4 |
| DG | 18 | 4.5 | ST | 2 | 0.4 |
| FA | 21 | 4.3 | SPB | 2 | 0.4 |
| TG | 19 | 3.8 | LPG | 1 | 0.2 |
| CL | 16 | 3.2 | LPI | 1 | 0.2 |
| LPC | 10 | 2.2 | LPS | 1 | 0.2 |
| Inf vs. Wt | Inf_TAP vs. Inf | |||
| Canonical Pathway | p-Value | z-Score | p-Value | z-Score |
| Accumulation of lipids | 4.23 × 10−3 | 1.23 | 4.23 × 10−3 | −0.54 |
| Inf vs. Wt | Inf_TAP vs. Inf | |||
| Lipid nomenclature | Log2Ratio | Log2Ratio | ||
| FA 14:0 (myristic acid) | −5.9 × 10−1 | 4.2 × 10−1 | ||
| TG 54:3|TG 18:1_18:1_18:1 (triolein) | −6.0 × 10−3 | 7.0 × 10−2 | ||
| FA 16:0 (palmitic acid) | 9.0 × 10−3 | −2.7 × 10−1 | ||
| FA 18:0 (stearic acid) | 1.5 × 10−2 | −2.3 × 10−1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, G.; Tosi, D.; Aldini, G.; Carini, M.; D’Amato, A. Evaluation of Inflammatory Cellular Model by Advanced Bioanalytic and Artificial Intelligence Analyses of Lipids: Lipidomic Landscape of Inflammaging. Cosmetics 2024, 11, 140. https://doi.org/10.3390/cosmetics11040140
Aiello G, Tosi D, Aldini G, Carini M, D’Amato A. Evaluation of Inflammatory Cellular Model by Advanced Bioanalytic and Artificial Intelligence Analyses of Lipids: Lipidomic Landscape of Inflammaging. Cosmetics. 2024; 11(4):140. https://doi.org/10.3390/cosmetics11040140
Chicago/Turabian StyleAiello, Gilda, Davide Tosi, Giancarlo Aldini, Marina Carini, and Alfonsina D’Amato. 2024. "Evaluation of Inflammatory Cellular Model by Advanced Bioanalytic and Artificial Intelligence Analyses of Lipids: Lipidomic Landscape of Inflammaging" Cosmetics 11, no. 4: 140. https://doi.org/10.3390/cosmetics11040140