Targeted Analysis of 46 Bile Acids to Study the Effect of Acetaminophen in Rat by LC-MS/MS



Abstract
:1. Introduction
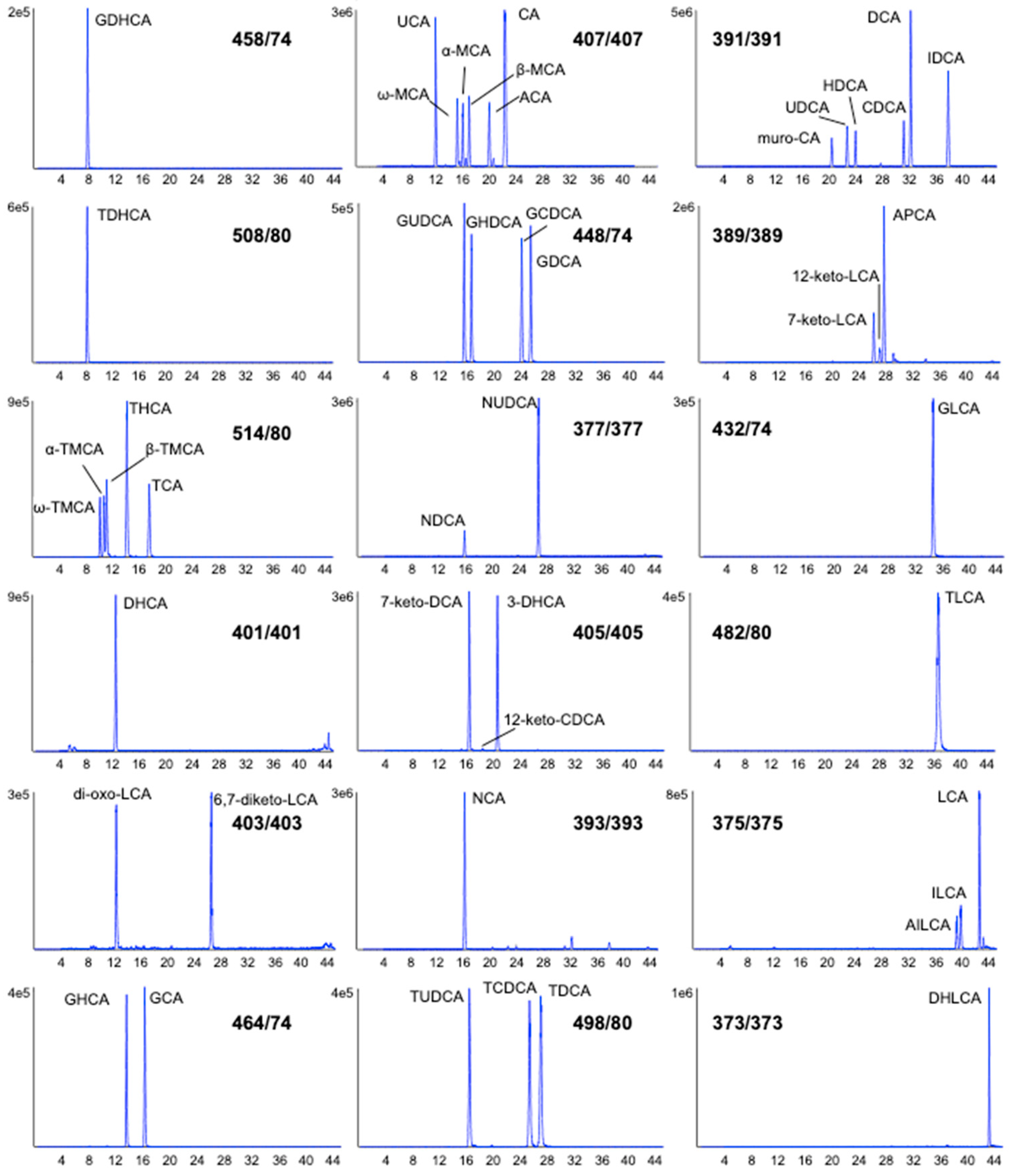
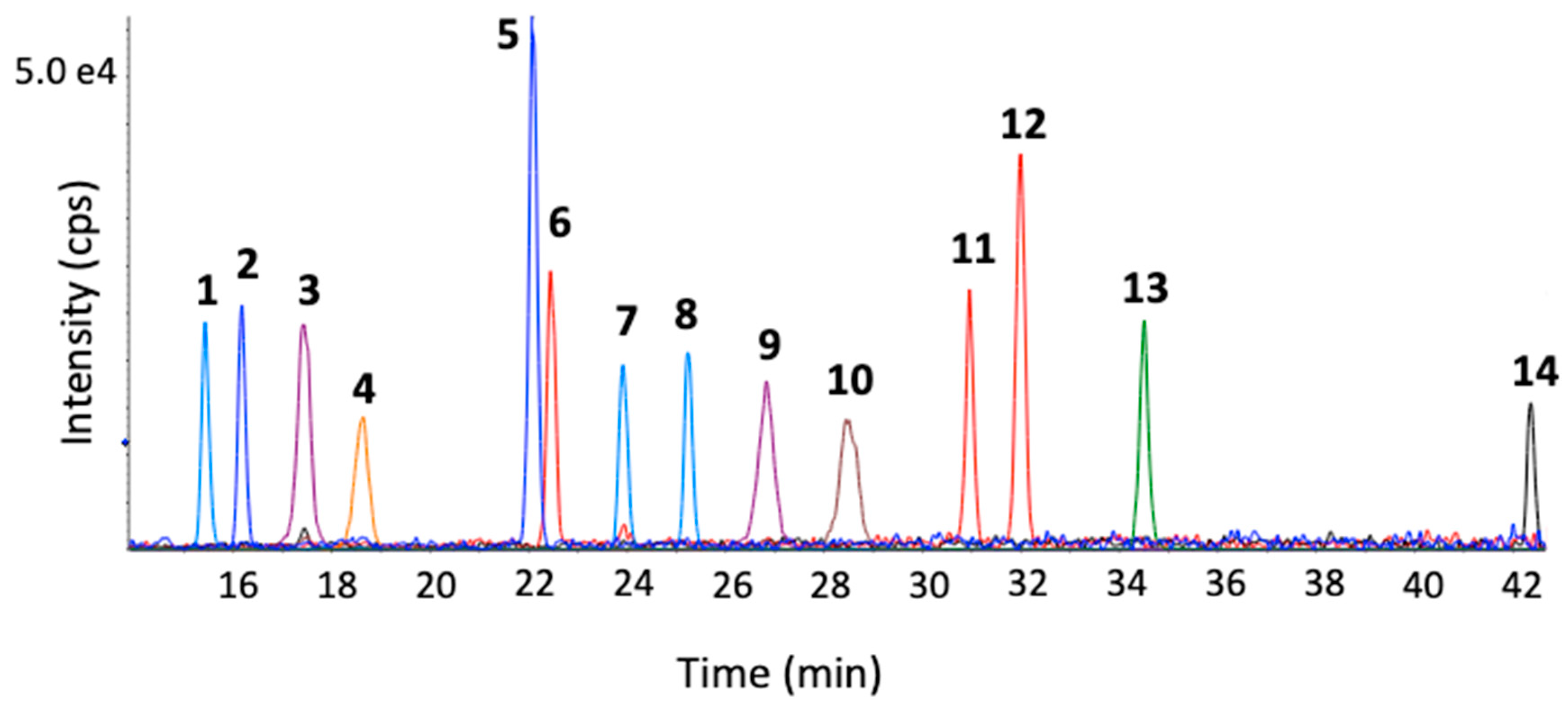
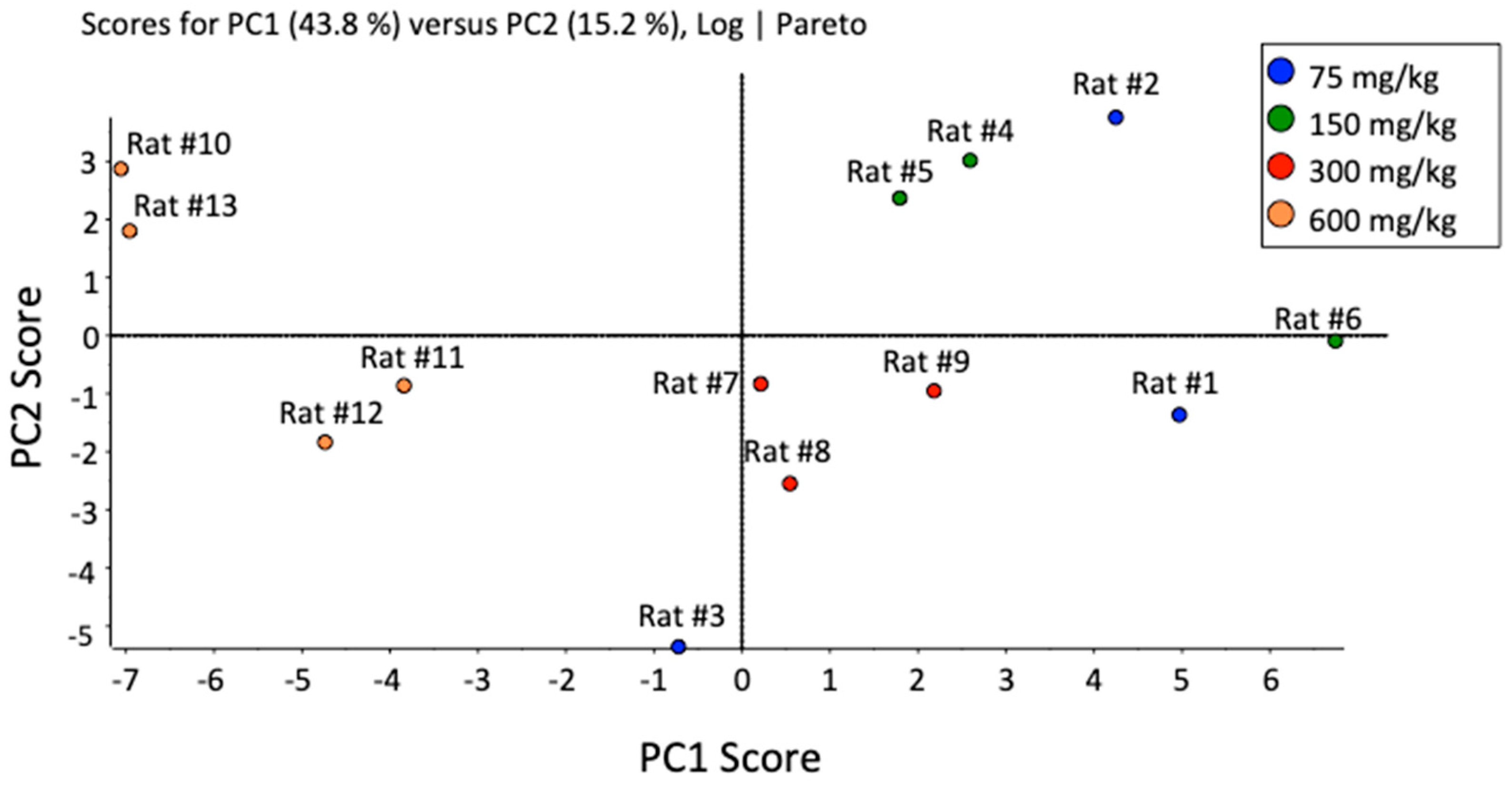
2. Results and Discussion
3. Materials and Methods
3.1. Materials
- The standard mix containing 46 bile acids (each at 2.5 nmol), except for deoxycholic acid (5 nmol) and taurohyocholic acid (6.5 nmol) was provided as a dried sample (Tube A). The bile acids in the standard mix were as follows: glycodehydrocholic acid (GDHCA), taurodehydrocholic acid (TDHCA), tauro-ω-muricholic acid (ω-TMCA), tauro-α-muricholic acid (α-TMCA), tauro-β-muricholic acid (β-TMCA), taurohyocholic acid (THCA), taurocholic acid (TCA), dehydrocholic acid (DHCA), dioxolithocholic acid (di-oxo-LCA), 6,7-diketolithocholic acid (6,7-diketo-LCA), glycohyocholic acid (GHCA), glycocholic acid (GCA), ursocholic acid (UCA), ω-muricholic acid (ω-MCA), α-muricholic acid (α-MCA), β-muricholic acid (β-MCA), allocholic acid (ACA), cholic acid (CA), glycoursodeoxycholic acid (GUDCA), glycohyodeoxycholic acid (GHDCA), glycochenodeoxycholic acid (GCDCA), glycodeoxycholic acid (GDCA), nordeoxycholic acid (NDCA), norursodeoxycholic acid (NUDCA), 7-ketodeoxycholic acid (7-keto-DCA), 12-ketodeoxycholic acid (12-keto-DCA), 3-dehydrocholic acid (3-DHCA), norcholic acid (NCA), tauroursodeoxycholic acid (TUDCA), taurochenodeoxycholic acid (TCDCA), taurodeoxycholic acid (TDCA), murocholic acid (muro-CA), ursodeoxycholic acid (UDCA), hyodeoxycholic acid (HDCA), chenodeoxycholic acid (CDCA), deoxycholic acid (DCA), isodeoxycholic acid (IDCA), 7-ketolithocholic acid (7-keto-LCA), 12-ketolithocholic acid (12-keto-LCA), apocholic acid (APCA), glycolithocholic acid (GLCA), taurolithocholic acid (TLCA), alloisolithocholic acid (AILCA), isolithocholic acid (ILCA), lithocholic acid (LCA) and dehydrolithocholic acid (DHLCA).
- Isotopically labeled bile acids were provided as an IS mix for normalization purposes. The labeled bile acids were present at between 0.1–0.75 nmol, as a dried sample (Tube B). The labeled bile acids in the IS mix were as follows: glycoursodeoxycholic acid-d4 (d4-GUDCA), glycocholic acid-d4 (d4-GCA), tauroursodeoxycholic acid-d4 (d4-TUDCA), taurocholic acid-d4 (d4-TCA), cholic acid-d4 (d4-CA), ursodeoxycholic acid-d4 (d4-UDCA), glycochenodeoxycholic acid-d4 (d4-GCDCA), glycodeoxycholic acid-d4 (d4-5GDCA), taurochenodeoxycholic acid-d4 (d4-TCDCA), taurodeoxycholic acid-d6 (d6-TDCA), chenodeoxycholic acid-d4 (d4-CDCA), deoxycholic acid-d4 (d4-DCA), glycolithocholic acid-d4 (d4-GLCA) and lithocholic acid-d4 (d4-LCA).
3.2. Sample Preparation
3.3. LC-MS/MS
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GDHCA | glycodehydrocholic acid |
| TDHCA | taurodehydrocholic acid |
| ω-TMCA | tauro-ω-muricholic acid |
| α-TMCA | tauro-α-muricholic acid |
| β-TMCA | tauro-β-muricholic acid |
| THCA | taurohyocholic acid |
| TCA | taurocholic acid |
| DHCA | dehydrocholic acid |
| di-oxo-LCA | dioxolithocholic acid |
| 6,7-diketo-LCA | 6,7-diketolithocholic acid |
| GHCA | glycohyocholic acid |
| GCA | glycocholic acid |
| UCA | ursocholic acid |
| ω-MCA | ω-muricholic acid |
| α-MCA | α-muricholic acid |
| β-MCA | β-muricholic acid |
| ACA | allocholic acid |
| CA | cholic acid |
| GUDCA | glycoursodeoxycholic acid |
| GHDCA | glycohyodeoxycholic acid |
| GCDCA | glycochenodeoxycholic acid |
| GDCA | glycodeoxycholic acid |
| NDCA | nordeoxycholic acid |
| NUDCA | norursodeoxycholic acid |
| 7-keto-DCA | 7-ketodeoxycholic acid |
| 12-keto-DCA | 12-ketodeoxycholic acid |
| 3-DHCA | 3-dehydrocholic acid |
| NCA | norcholic acid |
| TUDCA | tauroursodeoxycholic acid |
| TCDCA | taurochenodeoxycholic acid |
| TDCA | taurodeoxycholic acid |
| muro-CA | murocholic acid |
| UDCA | ursodeoxycholic acid |
| HDCA | hyodeoxycholic acid |
| CDCA | chenodeoxycholic acid |
| DCA | deoxycholic acid |
| IDCA | isodeoxycholic acid |
| 7-keto-LCA | 7-ketolithocholic acid |
| 12-keto-LCA | 12-ketolithocholic acid |
| APCA | apocholic acid |
| GLCA | glycolithocholic acid |
| TLCA | taurolithocholic acid |
| AILCA | alloisolithocholic acid |
| ILCA | isolithocholic acid |
| LCA | lithocholic acid |
| DHLCA | dehydrolithocholic acid |
References
- Russell, D.W. Fifty years of advances in bile acid synthesis and metabolism. J. Lipid Res. 2009, 50, S120–S125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.D. The role of bile acids in the overall regulation of steroid metabolism. Arch. Intern. Med. 1972, 130, 493–505. [Google Scholar] [CrossRef]
- Boyer, J.L. Bile formation and secretion. Compr. Physiol. 2013, 3, 1035–1078. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, A.F. Bile Acids: The Good, the Bad, and the Ugly. News Physiol. Sci. 1999, 14, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copple, B.L.; Jaeschke, H.; Klaassen, C.D. Oxidative stress and the pathogenesis of cholestasis. Semin. Liver Dis. 2010, 30, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Han, S.I.; Mitchell, C.; Gupta, S.; Studer, E.; Grant, S.; Hylemon, P.B.; Dent, P. Bile acids induce mitochondrial ROS, which promote activation of receptor tyrosine kinases and signaling pathways in rat hepatocytes. Hepatology 2004, 40, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Natarajan, R.; Payne, S.G.; Studer, E.J.; Spiegel, S.; Dent, P.; Hylemon, P.B. Deoxycholic acid activates the c-Jun N-terminal kinase pathway via FAS receptor activation in primary hepatocytes: Role of acidic sphingomyelinase-mediated ceramide generation in FAS receptor activation. J. Biol. Chem. 2004, 279, 5821–5828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeschke, H.; Gores, G.J.; Cederbaum, A.I.; Hinson, J.A.; Pessayre, D.; Lemasters, J.J. Mechanisms of hepatotoxicity. Toxicol. Sci. 2002, 65, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.S.; Weisburger, J.H.; Wynder, E.L. Effects of high risk and low risk diets for colon carcinogenesis on fecal microflora and steroids in man. J. Nutr. 1975, 105, 878–884. [Google Scholar] [CrossRef]
- Nagengast, F.M.; Grubben, M.J.A.L.; van Munster, I.P. Role of bile acids in colorectal carcinogenesis. Eur. J. Cancer 1995, 31, 1067–1070. [Google Scholar] [CrossRef] [Green Version]
- Larson, A.M.; Polson, J.; Fontana, R.J.; Davern, T.J.; Lalani, E.; Hynan, L.S.; Reisch, J.S.; SchiØdt, F.V.; Ostapowicz, G.; Shakil, A.O.; et al. Acetaminophen-induced acute liver failure: Results of a United States multicenter, prospective study. Hepatology 2005, 42, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.M. Acute liver failure in the United States. Semin. Liver Dis. 2003, 23, 217–226. [Google Scholar] [CrossRef]
- Woolbright, B.L.; McGill, M.R.; Staggs, V.S.; Winefield, R.D.; Gholami, P.; Olyaee, M.; Sharpe, M.R.; Curry, S.C.; Lee, W.M.; Jaeschke, H.; et al. Glycodeoxycholic acid levels as prognostic biomarker in acetaminophen-induced acute liver failure patients. Toxicol. Sci. 2014, 142, 436–444. [Google Scholar] [CrossRef] [Green Version]
- James, L.; Yan, K.; Pence, L.; Simpson, P.; Bhattacharyya, S.; Gill, P.; Letzig, L.; Kearns, G.; Beger, R. Comparison of Bile Acids and Acetaminophen Protein Adducts in Children and Adolescents with Acetaminophen Toxicity. PLoS ONE 2015, 10, e0131010. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, B.; Borude, P.; Edwards, G.; Walesky, C.; Cleveland, J.; Li, F.; Ma, X.; Apte, U. Role of bile acids in liver injury and regeneration following acetaminophen overdose. Am. J. Pathol. 2013, 183, 1518–1526. [Google Scholar] [CrossRef] [Green Version]
- Scherer, M.; Gnewuch, C.; Schmitz, G.; Liebisch, G. Rapid quantification of bile acids and their conjugates in serum by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B 2009, 877, 3920–3925. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kaneko, R.; Nomura, M.; Naito, H.; Kitamori, K.; Nakajima, T.; Ogawa, T.; Hattori, H.; Seno, H.; Ishii, A. Simple and rapid quantitation of 21 bile acids in rat serum and liver by UPLC-MS-MS: Effect of high fat diet on glycine conjugates of rat bile acids. Nagoya J. Med. Sci. 2013, 75, 57–72. [Google Scholar] [CrossRef]
- Han, J.; Liu, Y.; Wang, R.; Yang, J.; Ling, V.; Borchers, C.H. Metabolic profiling of bile acids in human and mouse blood by LC-MS/MS in combination with phospholipid-depletion solid-phase extraction. Anal. Chem. 2015, 87, 1127–1136. [Google Scholar] [CrossRef]
- Yousef, I.M.; Mignault, D.; Weber, A.M.; Tuchweber, B. Influence of dehydrocholic acid on the secretion of bile acids and biliary lipids in rats. Digestion 1990, 45, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Shefe, S.; Salen, G.; Hauser, S.; Dayal, B.; Batta, A.K. Metabolism of Iso-Bile Acids in the Rat. J. Biol. Chem. 1982, 257, 1401–1406. [Google Scholar]
- Voigt, W.; Thomas, P.J.; Hsia, S.L. Enzymatic studies of bile acid metabolism. I. 6Beta-hydroxylation of chenodeoxycholic and taurochenodeoxycholic acids by microsomal preparations of rat liver. J. Biol. Chem. 1968, 243, 3493–3499. [Google Scholar] [PubMed]
- Shiraki, K.; Ito, T.; Sugimoto, K.; Fuke, H.; Inoue, T.; Miyashita, K.; Yamanaka, T.; Suzuki, M.; Nabeshima, K.; Nakano, T.; et al. Different effects of bile acids, ursodeoxycholic acid and deoxycholic acid, on cell growth and cell death in human colonic adenocarcinoma cells. Int. J. Mol. Med. 2005, 16, 729–733. [Google Scholar] [PubMed]
- Cai, X.; Liu, Y.; Zhou, X.; Navaneethan, U.; Shen, B.; Guo, B. An LC-ESI-MS method for the quantitative analysis of bile acids composition in fecal materials. Biomed. Chromatogr. 2012, 26, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Tagliacozzi, D.; Mozzi, A.F.; Casetta, B.; Bertucci, P.; Bernardini, S.; Di Ilio, C.; Urbani, A.; Federici, G. Quantitative analysis of bile acids in human plasma by liquid chromatography-electrospray tandem mass spectrometry: A simple and rapid one-step method. Clin. Chem. Lab. Med. 2003, 41, 1633–1641. [Google Scholar] [CrossRef]
- Garcia-Canaveras, J.C.; Donato, M.T.; Castell, J.V.; Lahoz, A. Targeted profiling of circulating and hepatic bile acids in human, mouse, and rat using a UPLC-MRM-MS-validated method. J. Lipid Res. 2012, 53, 2231–2241. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bile Acid | RT (min) | 1st Transition (CE) | 2nd Transition (CE) | IS |
|---|---|---|---|---|
| GDHCA | 7.9 | 458.2/74.0 (−66) | 458.2/348.1 (−41) | CDCA-d4 |
| TDHCA | 8.2 | 508.2/80.0 (−123) | 508.2/124.1 (−67) | CDCA-d4 |
| T-ω-MCA | 10.1 | 514.2/80.0 (−135) | 514.2/107.0 (−82) | TCA-d4 |
| T-α-MCA | 10.7 | 514.2/80.0 (−135) | 514.2/107.0 (−82) | TCA-d4 |
| T-β-MCA | 11.1 | 514.2/80.0 (−135) | 514.2/124.0 (−65) | TCA-d4 |
| UCA | 11.9 | 407.2/407.2 (−15) | 407.2/343.1 (−46) | CA-d4 |
| di-oxo-LCA | 12.3 | 403.2/403.2 (−18) | 403.2/385.2 (−40) | CDCA-d4 |
| DHCA | 12.4 | 401.2/401.2 (−18) | 401.2/331.1 (−36) | CDCA-d4 |
| GHCA | 13.6 | 464.2/74.0 (−82) | 464.2/354.1 (−56) | GCA-d4 |
| THCA | 14.1 | 514.2/80.0 (−135) | 514.2/107.0 (−82) | TCA-d4 |
| ω-MCA | 15.1 | 407.2/407.2 (−15) | 407.2/371.1 (−43) | CA-d4 |
| GUDCA | 15.5 | 448.2/74.0 (−83) | 448.2/386.1 (−59) | GUDCA-d4 |
| NDCA | 15.8 | 377.2/377.2 (−15) | 377.2/331.1 (−46) | CDCA-d4 |
| α- MCA | 16.0 | 407.2/407.2 (−15) | 407.2/371.1 (−43) | CA-d4 |
| NCA | 16.1 | 393.2/393.2 (−15) | 393.2/375.1 (−45) | CDCA-d4 |
| GCA | 16.2 | 464.2/74.0 (−82) | 464.2/402.1 (−46) | GCA-d4 |
| 7-keto-DCA | 16.4 | 405.2/405.2 (−18) | 405.2/289.1 (−51) | CDCA-d4 |
| TUDCA | 16.4 | 498.2/80.0 (−130) | 498.2/107 (−82) | TUDCA-d4 |
| GHDCA | 16.6 | 448.2/74.0 (−83) | 448.2/386.1 (−59) | GUDCA-d4 |
| β-MCA | 16.9 | 407.2/407.2 (−15) | 407.2/371.1 (−43) | CA-d4 |
| TCA | 17.5 | 514.2/80.0 (−135) | 514.2/124.0 (−65) | TCA-d4 |
| 12-keto-CDCA | 18.4 | 405.2/405.2 (−18) | 405.2/387.1 (−45) | CDCA-d4 |
| ACA | 19.9 | 407.2/407.2 (−15) | 407.2/371.1 (−43) | CA-d4 |
| muro-CA | 20.3 | 391.2/391.2 (−15) | 391.2/343.1 (−53) | UDCA-d4 |
| 3-DHCA | 20.5 | 405.2/405.2 (−18) | 405.2/289.1 (−51) | CDCA-d4 |
| CA | 22.2 | 407.2/407.2 (−15) | 407.2/343.1 (−46) | CA-d4 |
| UDCA | 22.6 | 391.2/391.2 (−15) | 391.2/373.2 (−48) | UDCA-d4 |
| HDCA | 23.9 | 391.2/391.2 (−15) | 391.2/373.2 (−48) | CDCA-d4 |
| GCDCA | 24.0 | 448.2/74.0 (−83) | 448.2/404.2 (−46) | GCDCA-d4 |
| TCDCA | 25.3 | 498.2/80.0 (−130) | 498.2/124 (−64) | TCDCA-d4 |
| GDCA | 25.3 | 448.2/74.0 (−83) | 448.2/404.2 (−46) | GDCA-d4 |
| 7-keto-LCA | 26.1 | 389.2/389.2 (−18) | 389.2/354.1 (−43) | LCA-d4 |
| 6,7-diketo-LCA | 26.5 | 403.2/403.2 (−18) | 403.2/347.1 (−39) | LCA-d4 |
| NUDCA | 26.7 | 377.2/377.2 (−15) | 377.2/359.1 (−45) | CDCA-d4 |
| TDCA | 27.0 | 498.2/80.0 (−130) | 498.2/124 (−64) | TDCA-d6 |
| 12-keto-LCA | 27.1 | 389.2/389.2 (−18) | 389.2/354.1 (−43) | LCA-d4 |
| APCA | 27.6 | 389.2/389.2 (−18) | 389.2/371.1 (−43) | CDCA-d4 |
| CDCA | 31.1 | 391.2/391.2 (−15) | 391.2/373.2 (−48) | CDCA-d4 |
| DCA | 32.1 | 391.2/391.2 (−15) | 391.2/343.1 (−53) | DCA-d4 |
| GLCA | 34.6 | 432.2/74.0 (−66) | 432.2/388.1 (−45) | GLCA-d4 |
| TLCA | 36.6 | 482.2/80.0 (−135) | 482.2/107 (−80) | LCA-d4 |
| IDCA | 37.8 | 391.2/391.2 (−15) | 391.2/345.1 (−45) | DCA-d4 |
| AILCA | 39.1 | 375.2/375.2 (−15) | 375.2/45 (−50) | LCA-d4 |
| ILCA | 39.7 | 375.2/375.2 (−15) | 375.2/45 (−50) | LCA-d4 |
| LCA | 42.5 | 375.2/375.2 (−15) | 375.2/45 (−50) | LCA-d4 |
| DHLCA | 43.1 | 373.2/373.2 (−18) | 373.2/45 (−50) | LCA-d4 |
| RT (min) | Q1 (m/z) | Q3 (m/z) | CE (V) | |
|---|---|---|---|---|
| GUDCA-d4 | 15.5 | 452.3 | 74.0 | −41 |
| GCA-d4 | 16.2 | 468.3 | 74.0 | −45 |
| TUDCA-d4 | 17.4 | 502.3 | 80.0 | −73 |
| TCA-d4 | 18.7 | 518.3 | 80.0 | −80 |
| CA-d4 | 22.1 | 411.3 | 411.3 | −15 |
| UDCA-d4 | 22.5 | 395.3 | 395.3 | −15 |
| GCDCA-d4 | 23.8 | 452.3 | 74.0 | −37 |
| GDCA-d4 | 25.3 | 452.3 | 74.0 | −41 |
| TCDCA-d4 | 26.8 | 502.3 | 80.0 | −80 |
| TDCA-d6 | 28.7 | 504.3 | 80.0 | −80 |
| CDCA-d4 | 31 | 395.3 | 395.3 | −15 |
| DCA-d4 | 32 | 395.3 | 395.3 | −15 |
| GLCA-d4 | 34.5 | 436.3 | 74.0 | −41 |
| LCA-d4 | 42.4 | 379.3 | 379.3 | −15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prinville, V.; Ohlund, L.; Sleno, L. Targeted Analysis of 46 Bile Acids to Study the Effect of Acetaminophen in Rat by LC-MS/MS. Metabolites 2020, 10, 26. https://doi.org/10.3390/metabo10010026
Prinville V, Ohlund L, Sleno L. Targeted Analysis of 46 Bile Acids to Study the Effect of Acetaminophen in Rat by LC-MS/MS. Metabolites. 2020; 10(1):26. https://doi.org/10.3390/metabo10010026
Chicago/Turabian StylePrinville, Vivaldy, Leanne Ohlund, and Lekha Sleno. 2020. "Targeted Analysis of 46 Bile Acids to Study the Effect of Acetaminophen in Rat by LC-MS/MS" Metabolites 10, no. 1: 26. https://doi.org/10.3390/metabo10010026
APA StylePrinville, V., Ohlund, L., & Sleno, L. (2020). Targeted Analysis of 46 Bile Acids to Study the Effect of Acetaminophen in Rat by LC-MS/MS. Metabolites, 10(1), 26. https://doi.org/10.3390/metabo10010026