A New Approach for the Production of Selenium-Enriched and Probiotic Yeast Biomass from Agro-Industrial by-Products in a Stirred-Tank Bioreactor

 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Starchy Hydrolysates Characterization
2.2. Fermentation in Stirred Tank Bioreactor (STR) for Selenium Enrichment
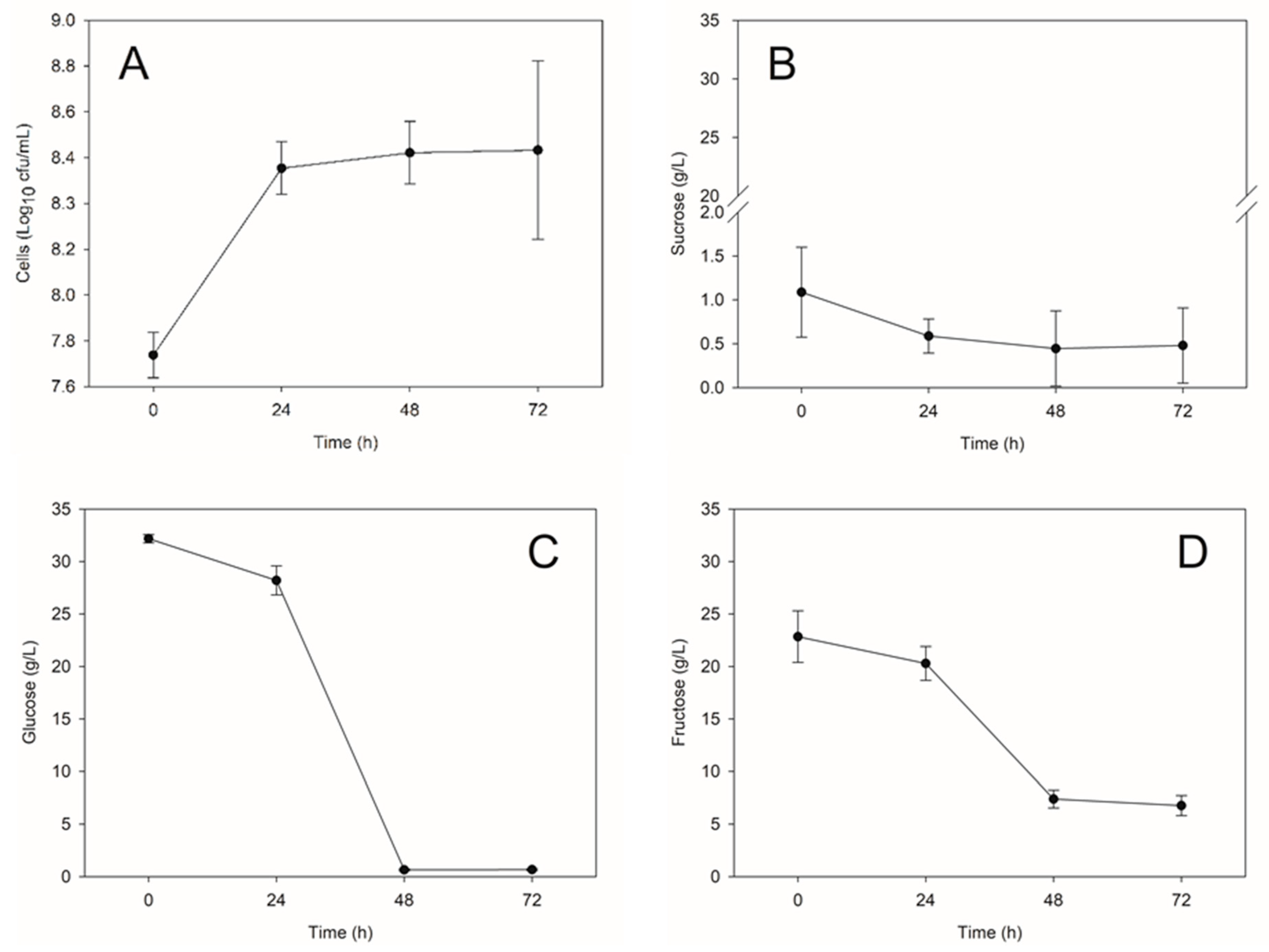
2.3. Evaluation of Biomass Production and β-Glucan Content of Probiotic Yeast in Stirred-Tank Bioreactor
3. Materials and Methods
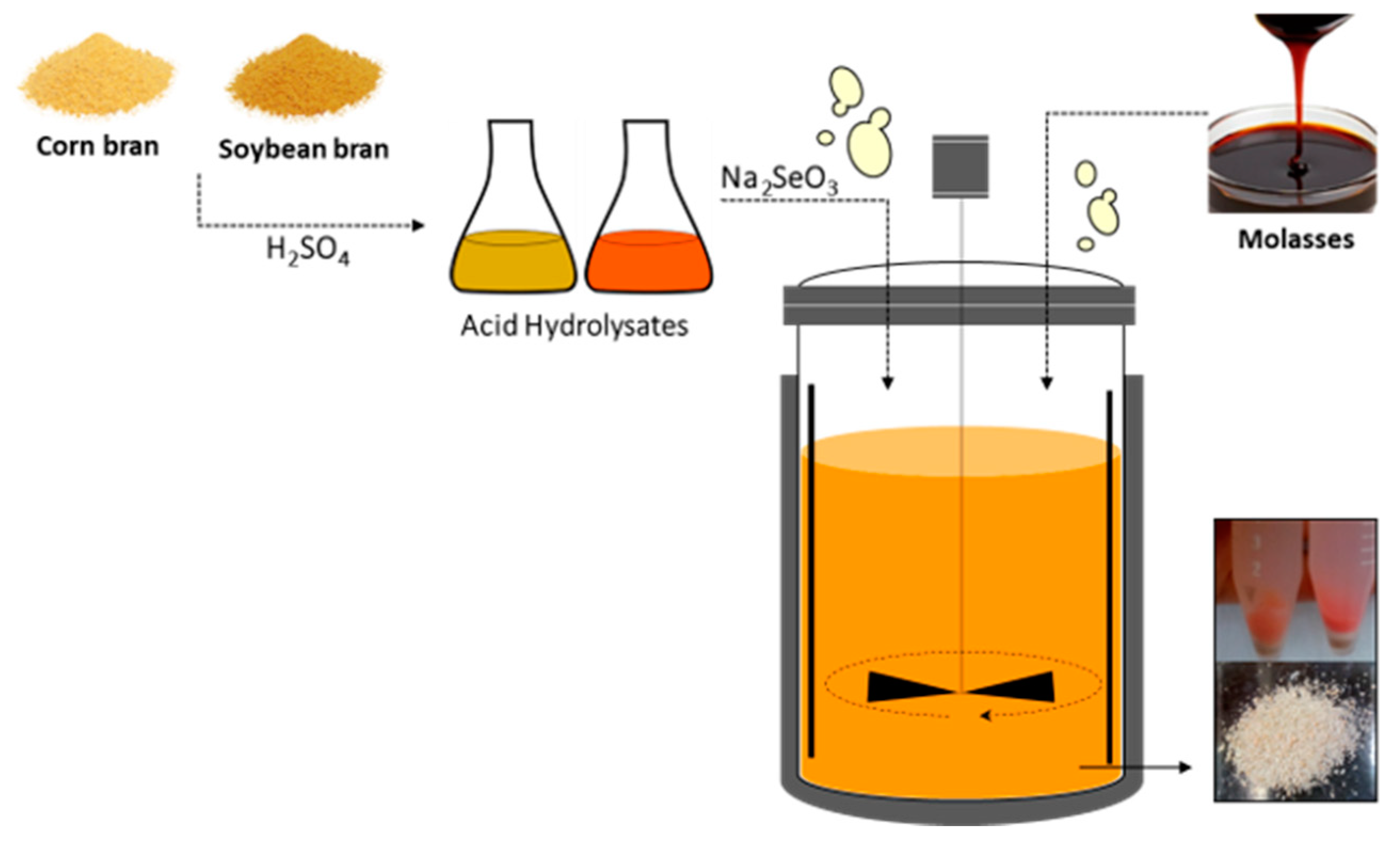
3.1. Obtention and Pretreatment of Agro-Industrial by-Products
3.2. Characterization of Starchy Acid Hydrolysates and Sugarcane Molasses
3.3. Microorganisms and Inoculum
3.4. Production of Se-Enriched Yeast Biomass in a Stirred-Tank Bioreactor
3.5. Production of Probiotic Yeast Biomass in a Stirred-Tank Bioreactor
3.6. Fermentation Analytical Methods and Kinetic Parameters
3.7. Determination of Selenium Uptake in the Se-Enriched Yeast Biomass
3.8. Determination of β-Glucan in Probiotic Yeast Biomass
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vohra, A.; Syal, P.; Madan, A. Probiotic yeasts in livestock sector. Anim. Feed Sci. Technol. 2016, 219, 31–47. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Probiotics in Animal Nutrition—Production, Impact and Regulation; FAO: Rome, Italy, 2016. [Google Scholar]
- Cangussu, A.S.R.; Cruz, P.A.; Aguiar, R.W.S.; Santos, G.R.; Campos, F.S.; Sobrinho, E.M.; Brandi, I.V.; Viana, K.F.; Nogueira, P.A.; Passos, F.J.V.; et al. Prediction of fermentative parameters from mathematical modeling using thermotolerant probiotic yeast. Bioresour. Technol. Rep. 2020, 11, 100522. [Google Scholar] [CrossRef]
- Guluarte, C.; Reyes-Becerril, M.; Gonzalez-Silvera, D.; Cuesta, A.; Ângulo, C.; Esteban, M.A. Probiotic properties and fatty acid composition of the yeast Kluyveromyces lactis M3. In vivo immunomodulatory activities in gilthead seabream (Sparus aurata). Fish Shellfish Immun. 2019, 94, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Enujiugha, V.N.; Badejo, A.A. Probiotic potentials of cereal-based beverages. Crit. Rev. Food Sci. Nutr. 2017, 57, 790–804. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Cui, S.W.; Xie, M. Bioactive Polysaccharides; Academic Press Elsevier: Cambridge, MA, USA, 2018; pp. 99–141. [Google Scholar]
- Orleans, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic applications of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Błażejak, S.; Gientka, I.; Bzducha-Wróbel, A. Accumulation and metabolism of selenium by yeast cells. Appl. Microbiol. Biotechnol. 2015, 99, 5373–5382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiyo, F.; Singh, M. Selenium nanoparticles: Potential in cancer gene and drug delivery. Nanomedicine 2017, 12, 1075–1089. [Google Scholar] [CrossRef]
- Paiva, F.A.; Netto, A.S.; Corrêa, L.B.; Silva, T.H.; Guimarães, I.C.S.B.; Del Claro, G.R.; Cunha, J.A.; Zanetti, M.A. Organic selenium supplementation increases muscle selenium content in growing lambs compared to inorganic source. Small Rumin. Res. 2019, 175, 57–64. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on lipid and amino acid metabolism in yeast cells. Biol. Trace Elem. Res. 2019, 187, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Bierla, K.; Jiménez-Lamana, J.; Kot, A.M.; Alcántara-Durán, J.; Piwowarek, K.; Błażejak, S.; Szpunar, J. Metabolic response of the yeast Candida utilis during enrichment in selenium. Int. J. Mol. Sci. 2020, 21, 5287. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.T.; Mahmud, A.; Javed, K.; Zahoor, I.; Mehmood, S.; Hussain, J.; Rehman, M.S. Organic and inorganic selenium in Aseel chicken diets: Effect on production performance. J. Appl. Poult. Res. 2018, 27, 292–298. [Google Scholar] [CrossRef]
- Shojadoost, B.; Taha-Abdelaziz, K.; Alkie, T.N.; Bekele-Yitbarek, A.; Barjesteh, N.; Laursen, A.; Smith, T.K.; Shojadoost, J.; Sharif, S. Supplemental dietary selenium enhances immune responses conferred by a vaccine against low pathogenicity avian influenza virus. Vet. Immunol. Immunop. 2020, 227, 110089. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, D.; Wei, G.; Liu, Z.; Ge, X. Selenium-enriched Candida utilis: Efficient preparation with l-methionine and antioxidant capacity in rats. J. Trace Elem. Med. Biol. 2013, 27, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yang, W.; Wang, M.; Miao, Y.; Cui, Z.; Li, Z.; Liang, D. Effects of selenium application on Se content and speciation in Lentinula edodes. Food Chem. 2018, 265, 182–188. [Google Scholar] [CrossRef]
- Mattioli, S.; Dal Bosco, A.; Duarte, J.M.M.; D’Amato, R.; Castellini, C.; Beone, G.M.; Fontanella, M.C.; Beghelli, D.; Regni, L.; Businelli, D.; et al. Use of Selenium-enriched olive leaves in the feed of growing rabbits: Effect on oxidative status, mineral profile and Selenium speciation of Longissimus dorsi meat. J. Trace Elem. Med. Bio. 2019, 51, 98–105. [Google Scholar] [CrossRef]
- Yue, S.; Zhang, H.; Zhen, H.; Lin, Z.; Qiao, Y. Selenium accumulation, speciation and bioaccessibility in selenium-enriched earthworm (Eisenia fetida). Microchem. J. 2019, 145, 1–8. [Google Scholar] [CrossRef]
- Ponce, M.; Giraldez, I.; Calero, S.; Ruiz-Azcona, P.; Morales, E.; Fernández-Díaz, C.; Hachero-Cruzado, I. Toxicity and biochemical transformation of selenium species in rotifer (Brachionus plicatilis) enrichments. Aquaculture 2017, 484, 105–111. [Google Scholar] [CrossRef]
- Cavrois-Rogacki, T.; Rolland, A.; Migaud, H.; Davie, A.; Monroig, A. Enriching Artemia nauplii with selenium from different sources and interactions with essential fatty acid incorporation. Aquaculture 2019, 520, 734677. [Google Scholar] [CrossRef]
- Ponce de León, C.A.; Bayón, M.M.; Paquin, C.; Caruso, J.A. Selenium incorporation into Saccharomyces cerevisiae cells: A study of different incorporation methods. J. Appl. Microbiol. 2002, 92, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Mapelli, V.; Hillestrøm, P.R.; Patil, K.; Larsen, E.H.; Olsson, L. The interplay between sulphur and selenium metabolism influences the intracellular redox balance in Saccharomyces cerevisiae. FEMS Yeast Res. 2011, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietary supplements market size, share & trend analysis report by ingredient (Botanicals, Vitamins, Minerals, Amino Acids, Enzymes), by product, by application, by end-use, and segment forecasts, 2018–2024. Available online: https://www.grandviewresearch.com/industry-analysis/dietary-supplements-market (accessed on 12 December 2020).
- Hamza, F.; Vaidya, A.; Apte, M.; Kumar, A.R.; Zinjarde, S. Selenium nanoparticle-enriched biomass of Yarrowia lipolytica enhances growth and survival of Artemia salina. Enzym. Microb. Technol. 2017, 106, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, S.; Khosravi-Darani, K.; Pourahmad, R.; Komeili, R. An experimental design for production of selenium-enriched yeast. World Appl. Sci. J. 2012, 19, 31–37. [Google Scholar] [CrossRef]
- Pérez-Corona, M.T.; Sánchez-Martínez, M.; Valderrama, M.J.; Rodríguez, M.E.; Cámara, C.; Madrid, Y. Selenium biotransformation by Saccharomyces cerevisiae and Saccharomyces bayanus during white wine manufacture: Laboratory-scale experiments. Food Chem. 2011, 124, 1050–1055. [Google Scholar] [CrossRef]
- Sánchez-Martínez, M.; Silva, E.G.P.; Pérez-Corona, T.; Cámara, C.; Ferreira, S.L.C.; Madrid, Y. Selenite biotransformation during brewing: Evaluation by HPLC–ICP-MS. Talanta 2012, 88, 272–276. [Google Scholar] [CrossRef]
- Martiniano, S.E.; Philippini, R.R.; Franco-Marcelino, P.R.; da Silva, S.S. Effect of selenium uptake on growth metabolism in yeasts for the production of enriched single-cell protein using agro-industrial by-products. Biomass Conv. Bioref. 2020. [Google Scholar] [CrossRef]
- Ingle, A.P.; Philippini, R.R.; Martiniano, S.E.; Franco-Marcelino, P.R.; Gupta, I.; Prasad, S.; da Silva, S.S. Bioresources and their Significance: Prospects and obstacles. In Current Developments in Biotechnology and Bioengineering Sustainable Bioresources for the Emerging Bioeconomy; Kataki, R., Pandey, A., Khanal, S.K., Pant, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 03–102. [Google Scholar]
- Martiniano, S.E.; Philippini, R.R.; Chandel, A.K.; Rosa, C.A.; Pagnocca, F.C.; da Silva, S.S. Evaluation of rice bran extract as a nitrogen source for improved hemicellulosic ethanol production from sugarcane bagasse by new xylose-fermenting yeast strains isolated from brazilian forests. Sugar Tech. 2014, 16, 1–8. [Google Scholar] [CrossRef]
- Philippini, R.R.; Martiniano, S.E.; Ingle, A.P.; Franco-Marcelino, P.R.; Silva, G.M.; Barbosa, F.G.; Santos, J.C.; Silva, S.S. Agroindustrial byproducts for the generation of biobased products: Alternatives for sustainable biorefineries. Front. Energy 2020, 8, 1–23. [Google Scholar] [CrossRef]
- Rostagno, H.S. Tabelas Brasileiras Para Aves e Suínos: Composição de Alimentos e Exigências Nutricionais, 4th ed.; UFV: Viçosa, Brazil, 2017. [Google Scholar]
- Lee, J.E.; Vadlani, P.V.; Faubion, J. Corn bran bioprocessing: Development of an integrated process for microbial lipids production. Bioresour. Technol. 2017, 243, 196–203. [Google Scholar] [CrossRef]
- Barzegar, S.; Wu, S.-B.; Noblet, J.; Swick, R.A. Metabolizable energy of corn, soybean meal and wheat for laying hens. Poult. Sci. J. 2019, 98, 5876–5882. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Ulven, C.A.; Johnson, M.A.; Durant, C.; Hossain, K.G. Pretreatment of wheat bran for suitable reinforcement in biocomposites. J. Renew Mater. 2017, 5, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.; Thanabalan, A.; Khanal, T.; Patterson, R.; Slominski, B.A.; Kiarie, E. Growth performance, gastrointestinalweight,microbialmetabolites and apparent retention of components in broiler chickens fed up to 11% rice bran in a corn-soybean meal diet without or with a multi-enzyme supplement. Anim. Nutr. 2019, 5, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Girio, F.M.; Fonseca, C.; Carvalheiro, F.; Duarte, L.C.; Marques, S.; Bogel-Łukasik, R. Hemicelluloses for fuel ethanol: A review. Bioresour. Technol. 2010, 101, 4775–4800. [Google Scholar] [CrossRef]
- Probst, K.V.; Vadlani, P.V. Production of single cell oil from Lipomyces starkeyi ATCC 56304 using biorefinery by-products. Bioresour. Technol. 2015, 198, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.F.; Guimarães, R.M.; Araújo, M.O.; Barcelos, K.R.; Carneiro, N.S.; Lima, D.S.; Santos, D.S.; Batista, K.A.; Fernandes, K.F.; Lima, M.C.P.M.; et al. Characterization of corn (Zea mays L.) bran as a new food ingredient for snack bars. LWT-Food Sci. Technol. 2018, 101, 812–818. [Google Scholar] [CrossRef]
- Reddy, K.N.; Duke, S.O. Soybean Mineral Composition and Glyphosate Use. In Processing and Impact on Active Components in Food; Preedy, V., Ed.; Academic Press Elsevier: Cambridge, MA, USA, 2015; pp. 369–376. [Google Scholar]
- Zhang, G.; Yao, X.; Wang, C.; Wang, D.; Wei, G. Transcriptome analysis reveals the mechanism underlying improved glutathione biosynthesis and secretion in Candida utilis during selenium enrichment. J. Biotechnol. 2019, 304, 89–96. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Kurek, E. Binding and conversion of selenium in Candida utilis ATCC 9950 yeasts in bioreactor culture. Molecules 2017, 22, 352. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Błażejak, S.; Płaczek, M. Spectrophotometric evaluation of selenium binding by Saccharomyces cerevisiae ATCC MYA-2200 and Candida utilis ATCC 9950 yeast. J. Trace Elem. Med. Biol. 2016, 35, 90–96. [Google Scholar] [CrossRef]
- Egressy-Molnár, O.; Ouerdane, L.; Győrfi, J.; Dernovics, M. Analogy in selenium enrichment and selenium speciation between selenized yeast Saccharomyces cerevisiae and Hericium erinaceus (lion’s mane mushroom). LWT-Food Sci. Technol. 2016, 68, 306–312. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on growth and antioxidative system of yeast cells. Mol. Biol. Rep. 2019, 46, 1797–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faramarzi, S.; Anzabi, Y.; Jafarizadeh-Malmiri, H. Selenium supplementation during fermentation with sugar beet molasses and Saccharomyces cerevisiae to increase bioethanol production. Green Process Synth. 2019, 8, 622–628. [Google Scholar] [CrossRef]
- Santos, J.F.; Canettieri, E.V.; Souza, S.M.A.; Rodrigues, R.C.L.B.; Martínez, E.A. Treatment of sugarcane vinasse from cachaça production for the obtainment of Candida utilis CCT 3469 biomass. Biochem. Eng. J. 2019, 148, 131–137. [Google Scholar] [CrossRef]
- Juszczyk, P.; Rymowicz, W.; Kita, A.; Rywińska, A. Biomass production by Yarrowia lipolytica yeast using waste derived from the production of ethyl esters of polyunsaturated fatty acids of flaxseed oil. Ind. Crop Prod. 2019, 138, 111590. [Google Scholar] [CrossRef]
- Nanis, I.; Hatzikamari, M.; Katharopoulos, E.; Boukouvala, E.; Ekateriniadou, L.; Litopoulou-Tzanetaki, E.; Gerasopoulos, D. Microbiological and physicochemical changes during fermentation of solid residue of olive mill wastewaters: Exploitation towards the production of an olive paste–type product. LWT-Food Sci. Technol. 2020, 117, 108671. [Google Scholar] [CrossRef]
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the World 2. Soybean—worldwide production, use, and constraints caused by pathogens and pests. Food Sec. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Ibáñez, M.A.; Blas, C.; Cámara, L.; Mateos, G.G. Chemical composition, protein quality and nutritive value of commercial soybean meals produced from beans from different countries: A meta-analytical study. Anim. Feed Sci. Tech. 2020, 267, 114531. [Google Scholar] [CrossRef]
- Dijken, J.P.; Scheffers, W.A. Redox balances in the metabolism of sugars by yeasts. FEMS Microbiol. Rev. 1986, 32, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, G.G.; Simili, F.F.; Augusto, J.G.; Bonacim, P.M.; Menegatto, L.S.; Gameiro, A.H. Economic gains from croplivestock integration in relation to conventional systems. R. Bras. Zootec. 2020, 49, e20190029. [Google Scholar] [CrossRef] [Green Version]
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. BB Rep. 2017, 10, 52–61. [Google Scholar] [CrossRef]
- Freitas, D.R.; Campos, J.M.S.; Marcondes, M.I.; Valadares Filho, S.C.; Franco, M.O.; Martins, E.C.; Rodrigues, B.M.C.; Oliveira, A.S. Levedura seca integral na alimentação de vacas lactantes. Arq. Bras. Med. Vet. Zootec. 2015, 67, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Porto, B.A.A.; Mangiapane, E.; Pessione, A.; Neves, M.J.; Pessione, E.; Martins, F.S. Evaluation of sodium selenite effects on the potential probiotic Saccharomyces cerevisiae UFMG A-905: A physiological and proteomic analysis. J. Funct. Foods 2015, 17, 828–836. [Google Scholar] [CrossRef] [Green Version]
- Rapoport, A.; Rusakova, A.; Khroustalyova, G.; Walker, G. Thermotolerance in Saccharomyces cerevisiae is linked to resistance toanhydrobiosis. Process Biochem. 2014, 49, 1889–1892. [Google Scholar] [CrossRef]
- Paula, B.P.; de Souza Lago, H.; Firmino, L.; Lemos Júnior, W.J.F.; Corrêa, M.F.D.; Guerra, A.F.; Pereira, K.S.; Coelho, M.A.Z. Technological features of Saccharomyces cerevisiae var. boulardii for potential probiotic wheat beer development. LWT-Food Sci. Technol. 2021, 135, 110233. [Google Scholar] [CrossRef]
- Singu, B.D.; Bhushette, P.R.; Uday, S.A. Thermo-tolerant Saccharomyces cerevisiae var. boulardii coated cornflakes as a potential probiotic vehicle. Food Biosci. 2020, 36, 100668. [Google Scholar] [CrossRef]
- Liu, F.; Zhu, M.J. Investigation on the production of carotenoid from molasses by Phaffia rhodozyma. Int. J. Mod. Biol. Med. 2017, 8, 1–13. [Google Scholar]
- Putra, M.D.; Abasaeedb, A.E.; Al-Zahranib, S.M. Prospective production of fructose and single cell protein from date palm waste. Electron. J. Biotechnol. 2020, 48, 46–52. [Google Scholar] [CrossRef]
- Vu, V.H.; Kim, K. High-Cell-Density Fed-Batch Culture of Saccharomyces cerevisiae KV-25 using molasses and corn steep liquor. J. Microbiol. Biotechnol. 2009, 19, 1603–1611. [Google Scholar] [CrossRef] [Green Version]
- Pengkumsri, N.; Sivamaruthi, B.S.; Sirilun, S.; Peerajan, S.; Kesika, P.; Chaiyasut, K.; Chaiyasut, C. Extraction of β-glucan from Saccharomyces cerevisiae: Comparison of different extraction methods and in vivo assessment of immunomodulatory effect in mice. Food Sci. Technol. Camp. 2017, 37, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.-Y.; Sung-Ho Son, S.-H.; Hong, S.-P.; Yi, S.-H.; Kang, S.H.; Lee, N.-K.; Paik, H.-D. Production of β-glucan, glutathione, and glutathione derivatives by probiotic Saccharomyces cerevisiae isolated from cucumber jangajji. LWT-Food Sci. Technol. 2019, 100, 114–118. [Google Scholar] [CrossRef]
- Kim, K.S.; Yun, H.S. Production of soluble B-glucan from the cell wall of Saccharomyces cerevisiae. Enzym. Microb. Technol. 2006, 39, 496–500. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosaiicyiic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Martins, D.S.R.; Leocádio, L.G.; Da Silveria, C.L.P. ICP-OES simultaneous determination of Ca, Cu, Fe, Mg, Mn, Na and P in biodiesel by axial and radial inductively coupled plasma-optical emission spectrometry. Anal Lett. 2008, 41, 1615–1622. [Google Scholar] [CrossRef]
- Yin, H.; Chen, Z.; Gu, Z.; Han, Y. Optimization of natural fermentative medium for selenium-enriched yeast by D-optimal mixture design. LWT-Food Sci. Technol. 2009, 42, 327–331. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Composition | Corn Bran Hydrolysate | Soybean Bran Hydrolysate |
|---|---|---|
| (g/L) | ||
| Total sugars | 175.72 ± 19.97 a,* | 38.61 ± 4.45 b |
| Reducing sugars | 47.01 ± 0.85 b | 12.48 ± 0.62 c,d |
| Glucose | 4.83 ± 1.2 c,d,e | 2.2 ± 1.16 d,e |
| Xylose | 3.69 ± 0.66 c,d,e | 4.41 ± 1.41 c,d,e |
| Arabinose | 3.35 ± 0.51 c,d,e | 0.44 ± 0.02 e |
| Cellobiose | 5.35 ± 0.28 c,d,e | 2.56 ± 0.27 d,e |
| Proteins | 7.82 ± 2.33 c,d,e | 13.52 ± 1.06 c |
| Phenols | 0.39 ± 0.05 e | 0.33 ± 0.05 e |
| Acetic acid | 0.33 ± 0.26 e | 0.42 ± 0.01 e |
| Glycerol | 0.67 ± 0.39 e | 1.69 ± 0.48 d,e |
| (mg/L) | ||
| 5-HMF | NA | 101 ± 13 |
| Furfural | NA | 10 ± 7 |
| Fe | 5.14 ± 0.90 b,* | 9.88 ± 8.94 b |
| Ca | 18.94 ± 8.89 b | 149.91 ± 56.9 b |
| Mg | 111.98 ± 29.66 b | 486.35 ± 167.41 b |
| Mn | NA | 1.19 ± 1.96 b |
| K | 463.38 ± 77.96 b | 4908.25 ± 1423.76 a |
| Na | NA | 10.90 ± 11.63 b |
| Se | 0.05 ± 0.04 b | 0.33 ± 0.11 b |
| Parameter | Time | ||
|---|---|---|---|
| 24 h | 48 | 72 | |
| YX/S (g/g) | 1.76 ± 0.28 | 1.77 ± 0.17 | 1.29 ± 0.07 |
| QX (g/L.h) | 0.25 ± 0.04 | 0.12 ± 0.01 | 0.09 ± 0.005 |
| YC (%) | 8.22 ± 0.26 | 5.50 ± 1.14 | 11.46 ± 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martiniano, S.E.; Fernandes, L.A.; Alba, E.M.; Philippini, R.R.; Tabuchi, S.C.T.; Kieliszek, M.; Santos, J.C.d.; da Silva, S.S. A New Approach for the Production of Selenium-Enriched and Probiotic Yeast Biomass from Agro-Industrial by-Products in a Stirred-Tank Bioreactor. Metabolites 2020, 10, 508. https://doi.org/10.3390/metabo10120508
Martiniano SE, Fernandes LA, Alba EM, Philippini RR, Tabuchi SCT, Kieliszek M, Santos JCd, da Silva SS. A New Approach for the Production of Selenium-Enriched and Probiotic Yeast Biomass from Agro-Industrial by-Products in a Stirred-Tank Bioreactor. Metabolites. 2020; 10(12):508. https://doi.org/10.3390/metabo10120508
Chicago/Turabian StyleMartiniano, Sabrina Evelin, Letícia Alves Fernandes, Edith Mier Alba, Rafael Rodrigues Philippini, Stephanie Caroline Tavares Tabuchi, Marek Kieliszek, Júlio César dos Santos, and Silvio Silvério da Silva. 2020. "A New Approach for the Production of Selenium-Enriched and Probiotic Yeast Biomass from Agro-Industrial by-Products in a Stirred-Tank Bioreactor" Metabolites 10, no. 12: 508. https://doi.org/10.3390/metabo10120508
APA StyleMartiniano, S. E., Fernandes, L. A., Alba, E. M., Philippini, R. R., Tabuchi, S. C. T., Kieliszek, M., Santos, J. C. d., & da Silva, S. S. (2020). A New Approach for the Production of Selenium-Enriched and Probiotic Yeast Biomass from Agro-Industrial by-Products in a Stirred-Tank Bioreactor. Metabolites, 10(12), 508. https://doi.org/10.3390/metabo10120508