Evaluation of Non-Uniform Sampling 2D 1H–13C HSQC Spectra for Semi-Quantitative Metabolomics


Abstract
:1. Introduction
2. Results
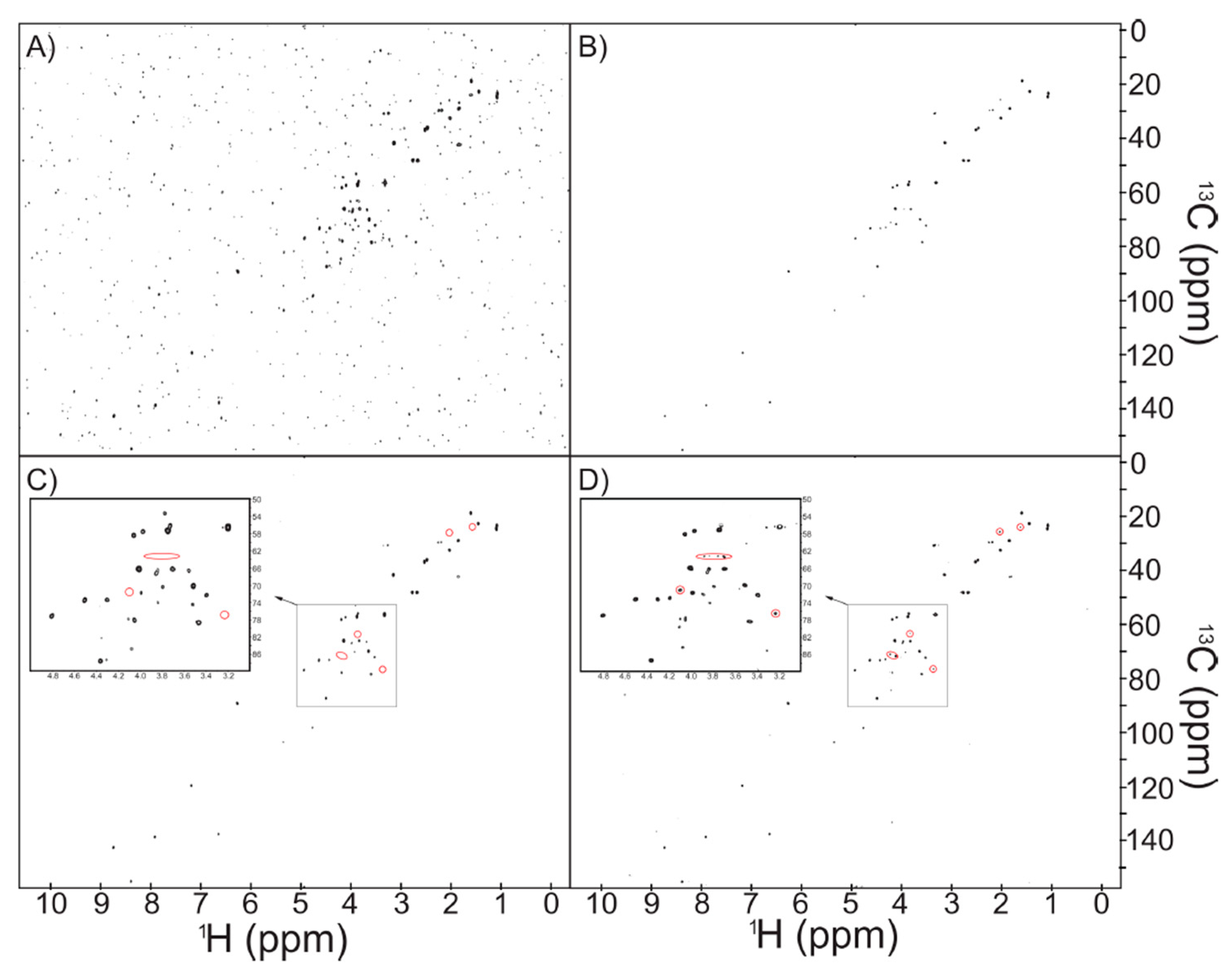
2.1. NUS Provides Enhanced Sensitivity
2.2. NUS Data Are Highly Linear
2.3. LOD and LOQ
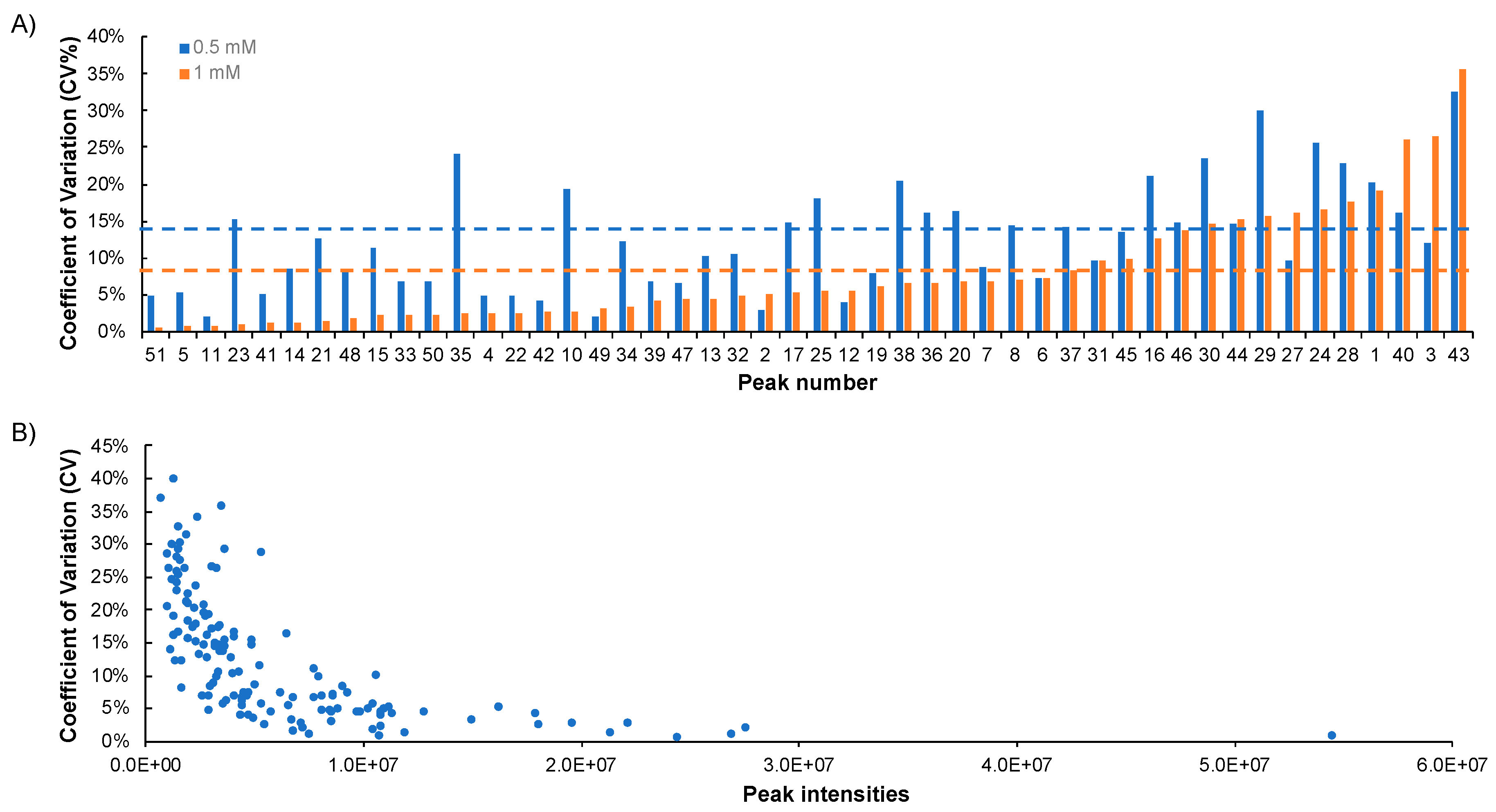
2.4. Repeatability of NUS
2.5. Stability of NUS
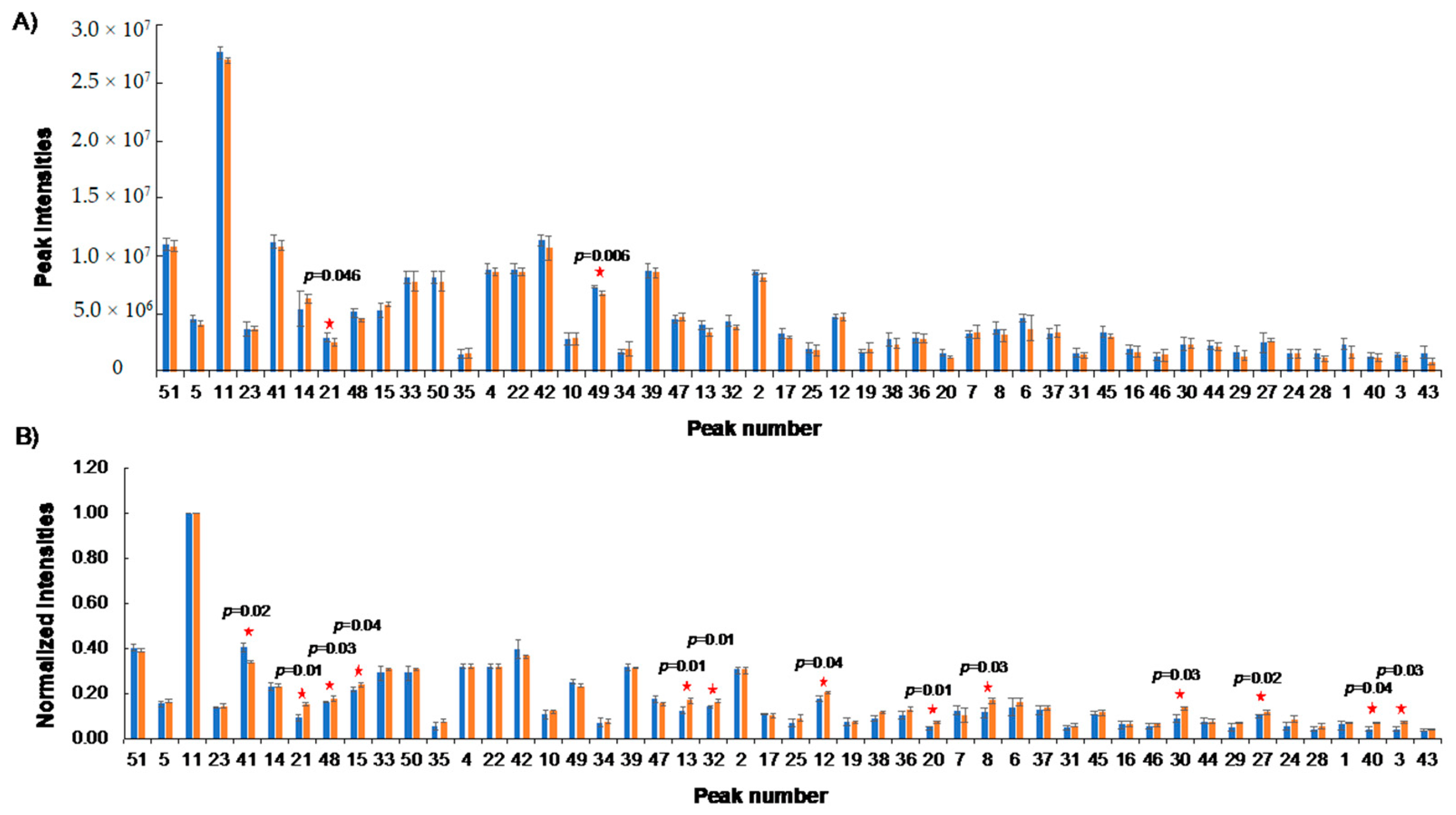
2.6. NUS Measurements across Systems
2.7. NUS on Plasma Sample
3. Materials and Methods
3.1. Sample Preparation
3.2. NMR Sample Preparation
3.3. NMR Experiments and Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Funct. Genomics 2002. [Google Scholar] [CrossRef]
- Nelson, D.; Cox, M. Lehninger Principles of Biochemistry, 4th ed.; Freeman and Company: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Ward, P.S.; Thompson, C.B. Signaling in control of cell growth and metabolism. Cold Spring Harb. Perspect. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Powers, R.; Riekeberg, E. New frontiers in metabolomics: From measurement to insight. F1000Research 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Krznar, P.; Erban, A.; Agazzi, A.; Martin-Levilain, J.; Supale, S.; Kopka, J.; Zamboni, N.; Maechler, P. Metabolomics identifies a biomarker revealing in vivo loss of functional β-cell mass before diabetes onset. Diabetes 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Le, W. Recent advances and perspectives of metabolomics-based investigations in Parkinson’s disease. Mol. Neurodegener. 2019, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchades-Carrasco, L.; Pineda-Lucena, A. Metabolomics Applications in Precision Medicine: An Oncological Perspective. Curr. Top. Med. Chem. 2017, 17, 2740–2751. [Google Scholar] [CrossRef] [Green Version]
- Wani, N.A.; Zhang, B.; Teng, K.Y.; Barajas, J.M.; Motiwala, T.; Hu, P.; Yu, L.; Bruschweiler, R.; Ghoshal, K.; Jacob, S.T. Reprograming of Glucose Metabolism by Zerumbone Suppresses Hepatocarcinogenesis. Mol. Cancer Res. 2018. [Google Scholar] [CrossRef] [Green Version]
- Cirulli, E.T.; Guo, L.; Swisher, C.L.; Shah, N.; Huang, L.; Napier, L.A.; Kirkness, E.F.; Spector, T.D.; Caskey, C.T.; Thorens, B.; et al. Profound Perturbation of the Metabolome in Obesity Is Associated with Health Risk. Cell Metab. 2019. [Google Scholar] [CrossRef] [Green Version]
- Ussher, J.R.; Elmariah, S.; Gerszten, R.E.; Dyck, J.R.B. The Emerging Role of Metabolomics in the Diagnosis and Prognosis of Cardiovascular Disease. J. Am. Coll. Cardiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.A.; Young, S.P. Metabolomics–A novel window into inflammatory disease. Swiss Med. Wkly. 2013. [Google Scholar] [CrossRef] [PubMed]
- Darst, B.F.; Koscik, R.L.; Hogan, K.J.; Johnson, S.C.; Engelman, C.D. Longitudinal plasma metabolomics of aging and sex. Aging 2019. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.E.; Ducrocq, A.J.; McDougall, D.J.; Garrett, T.J.; Yost, R.A. Comparison of blood plasma sample preparation methods for combined LC-MS lipidomics and metabolomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015. [Google Scholar] [CrossRef] [Green Version]
- Krug, S.; Kastenmüller, G.; Stückler, F.; Rist, M.J.; Skurk, T.; Sailer, M.; Raffler, J.; Römisch-Margl, W.; Adamski, J.; Prehn, C.; et al. The dynamic range of the human metabolome revealed by challenges. FASEB J. 2012, 26, 2607–2619. [Google Scholar] [CrossRef] [PubMed]
- Goodacre, R.; Vaidyanathan, S.; Dunn, W.B.; Harrigan, G.G.; Kell, D.B. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004, 22, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Bhinderwala, F.; Wase, N.; Dirusso, C.; Powers, R. Combining Mass Spectrometry and NMR Improves Metabolite Detection and Annotation. J. Proteome Res. 2018. [Google Scholar] [CrossRef]
- Bajad, S.; Shulaev, V. LC-MS-based metabolomics. Methods Mol. Biol. 2011, 708, 213–228. [Google Scholar] [CrossRef]
- Markley, J.L.; Brüschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr. Opin. Biotechnol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Lynn, K.S.; Cheng, M.L.; Chen, Y.R.; Hsu, C.; Chen, A.; Lih, T.M.; Chang, H.Y.; Huang, C.J.; Shiao, M.S.; Pan, W.H.; et al. Metabolite identification for mass spectrometry-based metabolomics using multiple types of correlated ion information. Anal. Chem. 2015, 87, 2143–2151. [Google Scholar] [CrossRef]
- McAlpine, J.B.; Chen, S.N.; Kutateladze, A.; Macmillan, J.B.; Appendino, G.; Barison, A.; Beniddir, M.A.; Biavatti, M.W.; Bluml, S.; Boufridi, A.; et al. The value of universally available raw NMR data for transparency, reproducibility, and integrity in natural product research. Nat. Prod. Rep. 2019, 36, 35–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dona, A.C.; Kyriakides, M.; Scott, F.; Shephard, E.A.; Varshavi, D.; Veselkov, K.; Everett, J.R. A guide to the identification of metabolites in NMR-based metabonomics/metabolomics experiments. Comput. Struct. Biotechnol. J. 2016, 14, 135–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schätzlein, M.P.; Becker, J.; Schulze-Sünninghausen, D.; Pineda-Lucena, A.; Herance, J.R.; Luy, B. Rapid two-dimensional ALSOFAST-HSQC experiment for metabolomics and fluxomics studies: Application to a 13C-enriched cancer cell model treated with gold nanoparticles. Anal. Bioanal. Chem. 2018, 410, 2793–2804. [Google Scholar] [CrossRef] [PubMed]
- Bingol, K.; Brüschweiler, R. Multidimensional APPROACHES to NMR-based metabolomics. Anal. Chem. 2014, 86, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The human metabolome database. Nucleic Acids Res. 2007. [Google Scholar] [CrossRef]
- Ulrich, E.L.; Akutsu, H.; Doreleijers, J.F.; Harano, Y.; Ioannidis, Y.E.; Lin, J.; Livny, M.; Mading, S.; Maziuk, D.; Miller, Z.; et al. BioMagResBank. Nucleic Acids Res. 2008. [Google Scholar] [CrossRef] [Green Version]
- Bingol, K.; Li, D.W.; Zhang, B.; Brüschweiler, R. Comprehensive metabolite identification strategy using multiple two-dimensional NMR spectra of a complex mixture implemented in the COLMARm web server. Anal. Chem. 2016. [Google Scholar] [CrossRef] [Green Version]
- Fardus-Reid, F.; Warren, J.; le Gresley, A. Validating heteronuclear 2D quantitative NMR. Anal. Methods. 2016, 8, 2013–2019. [Google Scholar] [CrossRef] [Green Version]
- Delaglio, F.; Walker, G.S.; Farley, K.; Sharma, R.; Hoch, J.; Arbogast, L.; Brinson, R.; Marino, J.P. Non-uniform sampling for all: More NMR spectral quality, less measurement time. Am. Pharm. Rev. 2017, 20, 339681. [Google Scholar]
- Hyberts, S.G.; Milbradt, A.G.; Wagner, A.B.; Arthanari, H.; Wagner, G. Application of iterative soft thresholding for fast reconstruction of NMR data non-uniformly sampled with multidimensional Poisson Gap scheduling. J. Biomol. NMR. 2012. [Google Scholar] [CrossRef] [Green Version]
- Mobli, M. Reducing seed dependent variability of non-uniformly sampled multidimensional NMR data. J. Magn. Reson. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suiter, C.L.; Paramasivam, S.; Hou, G.; Sun, S.; Rice, D.; Hoch, J.C.; Rovnyak, D.; Polenova, T. Sensitivity gains, linearity, and spectral reproducibility in nonuniformly sampled multidimensional MAS NMR spectra of high dynamic range. J. Biomol. NMR. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, M.T.; Ruben, D.; Griffin, R.G.; Herzfeld, J. Deterministic schedules for robust and reproducible non-uniform sampling in multidimensional NMR. J. Magn. Reson. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Schlippenbach, T.; Oefner, P.J.; Gronwald, W. Systematic Evaluation of Non-Uniform Sampling Parameters in the Targeted Analysis of Urine Metabolites by 1H,1H 2D NMR Spectroscopy. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Rai, R.K.; Sinha, N. Fast and accurate quantitative metabolic profiling of body fluids by nonlinear sampling of 1H-13C Two-dimensional nuclear magnetic resonance spectroscopy. Anal. Chem. 2012. [Google Scholar] [CrossRef]
- Le Guennec, A.; Giraudeau, P.; Caldarelli, S. Evaluation of fast 2D NMR for metabolomics. Anal. Chem. 2014. [Google Scholar] [CrossRef]
- Puig-Castellví, F.; Pérez, Y.; Piña, B.; Tauler, R.; Alfonso, I. Comparative analysis of 1 H NMR and 1 H- 13 C HSQC NMR metabolomics to understand the effects of medium composition in yeast growth. Anal. Chem. 2018, 90, 12422–12430. [Google Scholar] [CrossRef]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR. 1995. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Peak 1 | Peak 2 | Peak 3 | Peak 4 | Peak 5 | Peak 6 | Peak 7 |
|---|---|---|---|---|---|---|---|
| UDP | 1.00 | 1.00 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 |
| Cytidine | 0.80 | 1.00 | 0.99 | 1.00 | 1.00 | 1.00 | |
| Fructose | 0.99 | 1.00 | 0.99 | 1.00 | 1.00 | 1.00 | |
| Ribose 5-phosphate | 0.99 | 0.97 | 1.00 | 0.99 | 1.00 | 0.99 | |
| NAD | 0.99 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 0.99 |
| NAD | 1.00 | 0.99 | 0.99 | 1.00 | |||
| AMP | 0.97 | 0.99 | 0.96 | 0.98 | 0.99 | ||
| Glucose | 0.98 | 1.00 | 0.84 | 0.83 | 1.00 | ||
| Histidine | 0.97 | 1.00 | 0.99 | 1.00 | 1.00 | ||
| 2-Hydroxyglutaric acid | 0.99 | 0.99 | 0.99 | 0.99 | |||
| GTP | 0.99 | 0.99 | 1.00 | 0.99 | |||
| Leucine | 1.00 | 1.00 | 1.00 | 0.98 | |||
| Acetylcholine | 1.00 | 1.00 | 1.00 | ||||
| Cysteine | 1.00 | 1.00 | 0.97 | ||||
| Glucosamine | 0.94 | 0.99 | 1.00 | ||||
| Lysine | 1.00 | 1.00 | 1.00 | ||||
| Malic acid | 0.93 | 0.98 | 1.00 | ||||
| Alanine | 1.00 | 1.00 | |||||
| Arginine | 0.98 | 1.00 | |||||
| Choline | 1.00 | 1.00 | |||||
| Glutamic acid | 0.98 | 1.00 | |||||
| Glutamine | 1.00 | 1.00 | |||||
| Lactic acid | 1.00 | 0.95 | |||||
| Ornithine | 0.93 | 1.00 | |||||
| Citrate | 0.93 | ||||||
| Acetyl-phosphate | 1.00 | ||||||
| Fumaric acid | 0.99 | ||||||
| Pyruvic acid | 0.99 | ||||||
| Succinic acid | 1.00 |
| Metabolite | Peak 1 | Peak 2 | Peak 3 | Peak 4 | Peak 5 | Peak 6 | Peak 7 | Minimal Conc. (mM) |
|---|---|---|---|---|---|---|---|---|
| UDP | 0.013 | 0.021 | 0.022 | 0.024 | 0.016 | 0.021 | 0.026 | 0.013 |
| Cytidine | 0.033 | 0.018 | 0.019 | 0.016 | 0.024 | 0.023 | 0.016 | |
| Fructose | 0.035 | 0.028 | 0.044 | 0.056 | 0.026 | 0.026 | 0.026 | |
| Ribose 5-phosphate | 0.065 | 0.049 | 0.037 | 0.048 | 0.019 | 0.041 | 0.019 | |
| NAD | 0.034 | 0.018 | 0.024 | 0.013 | 0.020 | 0.021 | 0.022 | 0.013 |
| NAD | 0.024 | 0.024 | 0.029 | 0.020 | 0.020 | |||
| AMP | 0.013 | 0.018 | 0.029 | 0.022 | 0.025 | 0.013 | ||
| Glucose | 0.042 | 0.047 | 0.047 | 0.093 | 0.037 | 0.037 | ||
| Histidine | 0.036 | 0.034 | 0.022 | 0.022 | 0.027 | 0.022 | ||
| 2-Hydroxyglutaric acid | 0.039 | 0.046 | 0.032 | 0.022 | 0.022 | |||
| GTP | 0.033 | 0.021 | 0.023 | 0.026 | 0.021 | |||
| Leucine | 0.010 | 0.009 | 0.040 | 0.028 | 0.009 | |||
| Acetylcholine | 0.025 | 0.016 | 0.017 | 0.016 | ||||
| Cysteine | 0.058 | 0.038 | 0.047 | 0.038 | ||||
| Glucosamine | 0.058 | 0.048 | 0.028 | 0.028 | ||||
| Lysine | 0.048 | 0.021 | 0.019 | 0.019 | ||||
| Malic acid | 0.079 | 0.066 | 0.020 | 0.020 | ||||
| Alanine | 0.012 | 0.058 | 0.012 | |||||
| Arginine | 0.044 | 0.011 | 0.011 | |||||
| Choline | 0.019 | 0.023 | 0.019 | |||||
| Glutamic acid | 0.042 | 0.020 | 0.020 | |||||
| Glutamine | 0.020 | 0.019 | 0.019 | |||||
| Lactic acid | 0.011 | 0.067 | 0.011 | |||||
| Ornithine | 0.069 | 0.013 | 0.013 | |||||
| Acetyl-phosphate | 0.024 | 0.024 | ||||||
| Citrate | 0.014 | 0.014 | ||||||
| Fumaric acid | 0.025 | 0.025 | ||||||
| Pyruvic acid | 0.019 | 0.019 | ||||||
| Succinic acid | 0.009 | 0.009 |
| Metabolite | Peak 1 | Peak 2 | Peak 3 | Peak 4 | Peak 5 | Peak 6 | Peak 7 | Minimal Conc. (mM) |
|---|---|---|---|---|---|---|---|---|
| UDP | 0.044 | 0.071 | 0.075 | 0.081 | 0.055 | 0.068 | 0.087 | 0.044 |
| Cytidine | 0.111 | 0.061 | 0.064 | 0.054 | 0.081 | 0.076 | 0.054 | |
| Fructose | 0.117 | 0.094 | 0.148 | 0.187 | 0.086 | 0.086 | 0.086 | |
| Ribose 5-phosphate | 0.216 | 0.162 | 0.123 | 0.159 | 0.064 | 0.138 | 0.064 | |
| NAD | 0.112 | 0.059 | 0.081 | 0.044 | 0.067 | 0.071 | 0.075 | 0.044 |
| NAD | 0.079 | 0.080 | 0.097 | 0.066 | 0.066 | |||
| AMP | 0.044 | 0.059 | 0.098 | 0.073 | 0.083 | 0.044 | ||
| Glucose | 0.139 | 0.156 | 0.155 | 0.311 | 0.124 | 0.124 | ||
| Histidine | 0.121 | 0.113 | 0.074 | 0.075 | 0.092 | 0.074 | ||
| 2-Hydroxyglutaric acid | 0.131 | 0.152 | 0.106 | 0.072 | 0.072 | |||
| GTP | 0.111 | 0.070 | 0.077 | 0.088 | 0.070 | |||
| Leucine | 0.032 | 0.031 | 0.133 | 0.092 | 0.031 | |||
| Acetylcholine | 0.082 | 0.054 | 0.056 | 0.054 | ||||
| Cysteine | 0.194 | 0.126 | 0.158 | 0.126 | ||||
| Glucosamine | 0.194 | 0.160 | 0.094 | 0.094 | ||||
| Lysine | 0.161 | 0.069 | 0.064 | 0.064 | ||||
| Malic acid | 0.265 | 0.222 | 0.066 | 0.066 | ||||
| Alanine | 0.039 | 0.194 | 0.039 | |||||
| Arginine | 0.147 | 0.037 | 0.037 | |||||
| Choline | 0.065 | 0.077 | 0.065 | |||||
| Glutamic acid | 0.139 | 0.068 | 0.068 | |||||
| Glutamine | 0.066 | 0.064 | 0.064 | |||||
| Lactic acid | 0.038 | 0.223 | 0.038 | |||||
| Ornithine | 0.229 | 0.045 | 0.045 | |||||
| Acetylphosphate | 0.081 | 0.081 | ||||||
| Citrate | 0.048 | 0.048 | ||||||
| Fumaric acid | 0.084 | 0.084 | ||||||
| Pyruvic acid | 0.063 | 0.063 | ||||||
| Succinic acid | 0.087 | 0.087 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Powers, R.; O’Day, E.M. Evaluation of Non-Uniform Sampling 2D 1H–13C HSQC Spectra for Semi-Quantitative Metabolomics. Metabolites 2020, 10, 203. https://doi.org/10.3390/metabo10050203
Zhang B, Powers R, O’Day EM. Evaluation of Non-Uniform Sampling 2D 1H–13C HSQC Spectra for Semi-Quantitative Metabolomics. Metabolites. 2020; 10(5):203. https://doi.org/10.3390/metabo10050203
Chicago/Turabian StyleZhang, Bo, Robert Powers, and Elizabeth M. O’Day. 2020. "Evaluation of Non-Uniform Sampling 2D 1H–13C HSQC Spectra for Semi-Quantitative Metabolomics" Metabolites 10, no. 5: 203. https://doi.org/10.3390/metabo10050203