Changes in the Fecal Metabolome Are Associated with Feeding Fiber Not Health Status in Cats with Chronic Kidney Disease



Abstract
:1. Introduction
2. Results
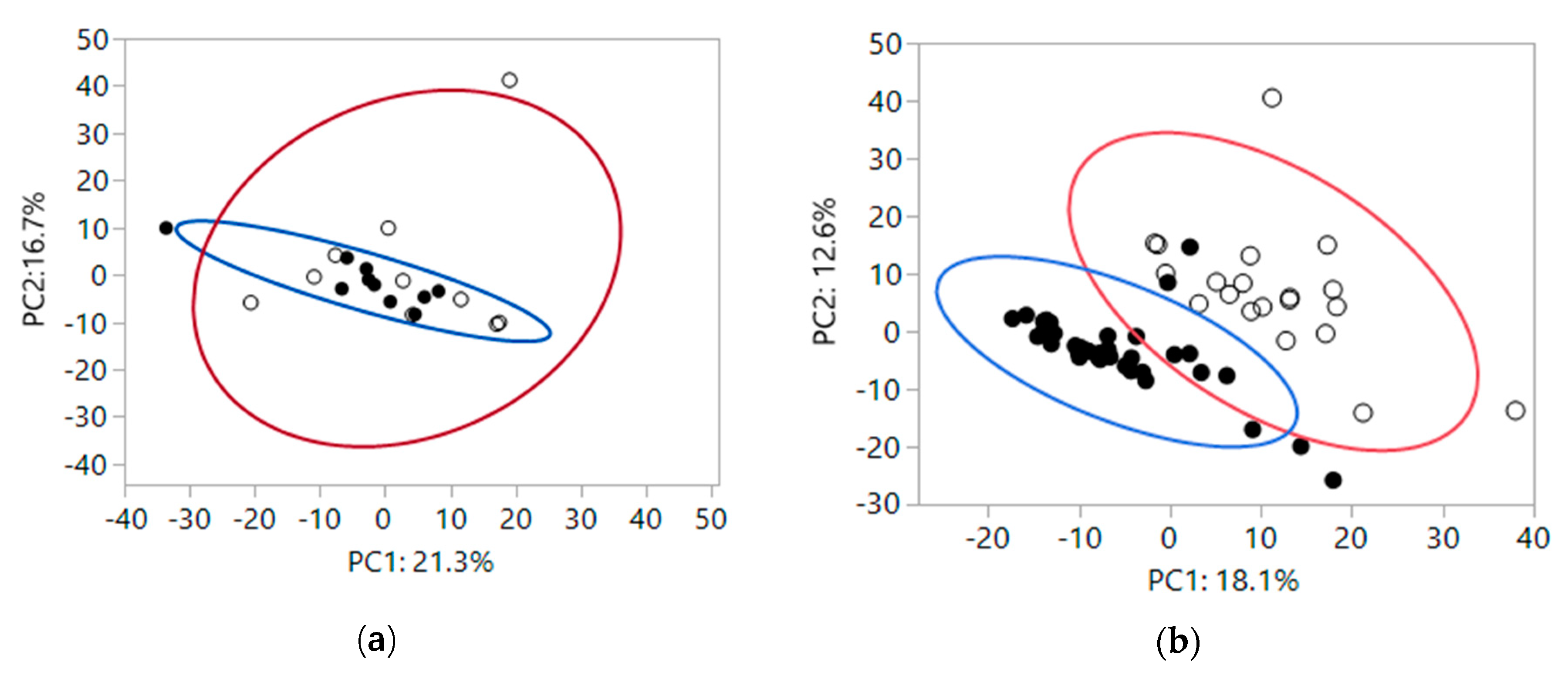
2.1. Fecal Metabolomics at Baseline and after Treatment with Different Fiber Sources
2.2. Treatment Differences in Fecal Metabolites Based on Health Status of Cats and Fiber Source Fed
3. Discussion
3.1. Fecal Metabololites Generated by the Host that Were Different after Treatment with Fiber Sources
3.2. Fecal Metabololites Generated by the Gut Microbes or by the Host that Were Different after Treatment with Fiber Sources
4. Materials and Methods
4.1. Participants and Study Design
4.2. Foods
4.3. Fecal Sample Collection and Fecal Metabolome
4.4. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hall, J.A.; Yerramilli, M.; Obare, E.; Yerramilli, M.; Jewell, D.E. Comparison of serum concentrations of symmetric dimethylarginine and creatinine as kidney function biomarkers in cats with chronic kidney disease. J. Vet. Intern. Med. 2014, 28, 1676–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.A.; Jackson, M.I.; Farace, G.; Yerramilli, M.; Jewell, D.E. Influence of dietary ingredients on lean body percent, uremic toxin concentrations, and kidney function in senior-adult cats. Metabolites 2019, 9, 238. [Google Scholar] [CrossRef] [Green Version]
- Ephraim-Gebreselassie, E.; Jackson, M.; Jewell, D. Nutritional intervention to reduce uremic toxin accumulation and cachexia in renal cats. In Current Developments in Nutrition; Odle, J., Ed.; American Society of Nutrition: Boston, MA, USA, 2018; Volume 2, p. 7. [Google Scholar]
- McGregor, D.O.; Dellow, W.J.; Robson, R.A.; Lever, M.; George, P.M.; Chambers, S.T. Betaine supplementation decreases post-methionine hyperhomocysteinemia in chronic renal failure. Kidney Int. 2002, 61, 1040–1046. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Pan, Y.; Wang, X.; Fan, C.Y.; Zhu, Q.; Li, J.M.; Wang, S.J.; Kong, L.D. Betaine reduces serum uric acid levels and improves kidney function in hyperuricemic mice. Planta Med. 2014, 80, 39–47. [Google Scholar] [CrossRef]
- Nallu, A.; Sharma, S.; Ramezani, A.; Muralidharan, J.; Raj, D. Gut microbiome in chronic kidney disease: Challenges and opportunities. Transl. Res. 2017, 179, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.Y.; Chen, D.Q.; Chen, L.; Liu, J.R.; Vaziri, N.D.; Guo, Y.; Zhao, Y.Y. Microbiome-metabolome reveals the contribution of gut-kidney axis on kidney disease. J. Transl. Med. 2019, 17, 5. [Google Scholar] [CrossRef] [Green Version]
- Rochus, K.; Janssens, G.P.; Hesta, M. Dietary fibre and the importance of the gut microbiota in feline nutrition: A review. Nutr. Res. Rev. 2014, 27, 295–307. [Google Scholar] [CrossRef]
- Vairakkani, R.; Fernando, M.E.; Raj, T.Y. Metabolome and microbiome in kidney diseases. Saudi J. Kidney Dis. Transpl. 2020, 31, 1–9. [Google Scholar] [CrossRef]
- Hall, J.A.; Jackson, M.I.; Jewell, D.E.; Ephraim, E. Chronic kidney disease in cats alters response of the plasma metabolome and fecal microbiome to dietary fiber. PLoS ONE 2020, 15, e0235480. [Google Scholar] [CrossRef]
- International Renal Interest Society. IRIS Staging of CKD (Modified 2019). Available online: http://www.iris-kidney.com/pdf/3_staging-of-ckd.pdf (accessed on 28 June 2020).
- Liu, S.; Liang, S.; Liu, H.; Chen, L.; Sun, L.; Wei, M.; Jiang, H.; Wang, J. Metabolite profiling of feces and serum in hemodialysis patients and the effect of medicinal charcoal tablets. Kidney Blood Press. Res. 2018, 43, 755–767. [Google Scholar] [CrossRef]
- Barrios, C.; Spector, T.D.; Menni, C. Blood, urine and faecal metabolite profiles in the study of adult renal disease. Arch. Biochem. Biophys. 2016, 589, 81–92. [Google Scholar] [CrossRef]
- Li, R.; Zeng, L.; Xie, S.; Chen, J.; Yu, Y.; Zhong, L. Targeted metabolomics study of serum bile acid profile in patients with end-stage renal disease undergoing hemodialysis. PeerJ 2019, 7, e7145. [Google Scholar] [CrossRef] [Green Version]
- Saggi, S.J.; Mercier, K.; Gooding, J.R.; Friedman, E.; Vyas, U.; Ranganathan, N.; Ranganathan, P.; McRitchie, S.; Sumner, S. Metabolic profiling of a chronic kidney disease cohort reveals metabolic phenotype more likely to benefit from a probiotic. Int. J. Probiotics Prebiotics 2017, 12, 43–54. [Google Scholar]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuno, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Fuke, N.; Nagata, N.; Suganuma, H.; Ota, T. Regulation of gut microbiota and metabolic endotoxemia with dietary factors. Nutrients 2019, 11, 2277. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Lee, M.J.; Cheung, C.; Ju, J.H.; Chen, Y.K.; Liu, B.; Hu, L.Q.; Yang, C.S. Analysis of multiple metabolites of tocopherols and tocotrienols in mice and humans. J. Agric. Food Chem. 2010, 58, 4844–4852. [Google Scholar] [CrossRef] [Green Version]
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sido, B.; Hack, V.; Hochlehnert, A.; Lipps, H.; Herfarth, C.; Droge, W. Impairment of intestinal glutathione synthesis in patients with inflammatory bowel disease. Gut 1998, 42, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Volani, C.; Paglia, G.; Smarason, S.V.; Pramstaller, P.P.; Demetz, E.; Pfeifhofer-Obermair, C.; Weiss, G. Metabolic signature of dietary iron overload in a mouse model. Cells 2018, 7, 264. [Google Scholar] [CrossRef] [Green Version]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Nagahashi, M.; Terracina, K.P.; Takabe, K. Emerging role of sphingosine-1-phosphate in inflammation, cancer, and lymphangiogenesis. Biomolecules 2013, 3, 408. [Google Scholar] [CrossRef] [Green Version]
- Merrill, A.H., Jr.; Schmelz, E.M.; Wang, E.; Dillehay, D.L.; Rice, L.G.; Meredith, F.; Riley, R.T. Importance of sphingolipids and inhibitors of sphingolipid metabolism as components of animal diets. J. Nutr. 1997, 127, 830S–833S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrill, A.H., Jr.; Schmelz, E.M.; Dillehay, D.L.; Spiegel, S.; Shayman, J.A.; Schroeder, J.J.; Riley, R.T.; Voss, K.A.; Wang, E. Sphingolipids--the enigmatic lipid class: Biochemistry, physiology, and pathophysiology. Toxicol. Appl. Pharmacol. 1997, 142, 208–225. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Lee, Y.S.; Lee, Y.M.; Oh, S.; Yun, Y.P.; Yoo, H.S. Elevation of sphingoid base 1-phosphate as a potential contributor to hepatotoxicity in fumonisin b1-exposed mice. Arch. Pharm. Res. 2007, 30, 962–969. [Google Scholar] [CrossRef]
- Heung, L.J.; Luberto, C.; Del Poeta, M. Role of sphingolipids in microbial pathogenesis. Infect. Immun. 2006, 74, 28–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppe, L.; Fouque, D.; Soulage, C.O. The role of gut microbiota and diet on uremic retention solutes production in the context of chronic kidney disease. Toxins 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koistinen, V.M.; Karkkainen, O.; Borewicz, K.; Zarei, I.; Jokkala, J.; Micard, V.; Rosa-Sibakov, N.; Auriola, S.; Aura, A.M.; Smidt, H.; et al. Contribution of gut microbiota to metabolism of dietary glycine betaine in mice and in vitro colonic fermentation. Microbiome 2019, 7, 103. [Google Scholar] [CrossRef]
- Bush, K.T.; Singh, P.; Nigam, S.K. Gut-derived uremic toxin handling in vivo requires oat-mediated tubular secretion in chronic kidney disease. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryniarski, M.A.; Hamarneh, F.; Yacoub, R. The role of chronic kidney disease-associated dysbiosis in cardiovascular disease. Exp. Biol. Med. 2019, 244, 514–525. [Google Scholar] [CrossRef]
- Yang, B.; Liu, D.; Li, C.Z.; Liu, F.Y.; Peng, Y.M.; Jiang, Y.S. 1-methylhydantoin cytotoxicity on renal proximal tubular cells in vitro. Ren. Fail. 2007, 29, 1025–1029. [Google Scholar] [CrossRef]
- Davies, R. The metabolomic quest for a biomarker in chronic kidney disease. Clin. Kidney J. 2018, 11, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.A.; Wojcicki, B.J.; Middelbos, I.S.; Vester, B.M.; Swanson, K.S.; Fahey, G.C., Jr. Dietary cellulose, fructooligosaccharides, and pectin modify fecal protein catabolites and microbial populations in adult cats. J. Anim. Sci. 2010, 88, 2978–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Chu, L.; Zhang, K.; Zhang, Y.; Jin, X.; Jiang, H. Mechanism underlying an elevated serum bile acid level in chronic renal failure patients. Int. Urol. Nephrol. 2015, 47, 345–351. [Google Scholar] [CrossRef]
- Jimenez, F.; Monte, M.J.; El-Mir, M.Y.; Pascual, M.J.; Marin, J.J. Chronic renal failure-induced changes in serum and urine bile acid profiles. Dig. Dis. Sci. 2002, 47, 2398–2406. [Google Scholar] [CrossRef]
- Mareckova, O.; Skala, I.; Marecek, Z.; Maly, J.; Kocandrle, V.; Schuck, O.; Blaha, J.; Prat, V. Bile composition in patients with chronic renal insufficiency. Nephrol. Dial. Transplant. 1990, 5, 423–425. [Google Scholar] [CrossRef]
- Wu, I.W.; Gao, S.S.; Chou, H.C.; Yang, H.Y.; Chang, L.C.; Kuo, Y.L.; Dinh, M.C.V.; Chung, W.H.; Yang, C.W.; Lai, H.C.; et al. Integrative metagenomic and metabolomic analyses reveal severity-specific signatures of gut microbiota in chronic kidney disease. Theranostics 2020, 10, 5398–5411. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, S.; Li, S.; Zhao, L.; Hao, Y.; Qin, J.; Zhang, L.; Zhang, C.; Bian, W.; Zuo, L.; et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut 2020. [Google Scholar] [CrossRef] [PubMed]
- NRC. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Stewart, M.L.; Timm, D.A.; Slavin, J.L. Fructooligosaccharides exhibit more rapid fermentation than long-chain inulin in an in vitro fermentation system. Nutr. Res. 2008, 28, 329–334. [Google Scholar] [CrossRef]
- Skinner, R.C.; Gigliotti, J.C.; Ku, K.M.; Tou, J.C. A comprehensive analysis of the composition, health benefits, and safety of apple pomace. Nutr. Rev. 2018, 76, 893–909. [Google Scholar] [CrossRef]
- de Godoy, M.R.C.; Hervera, M.; Swanson, K.S.; Fahey, G.C. Innovations in canine and feline nutrition: Technologies for food and nutrition assessment. Annu. Rev. Anim. Biosci. 2016, 4, 311–333. [Google Scholar] [CrossRef]
- Hall, J.A.; Melendez, L.D.; Jewell, D.E. Using gross energy improves metabolizable energy predictive equations for pet foods whereas undigested protein and fiber content predict stool quality. PLoS ONE 2013, 8, e54405. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.A.; Jackson, M.I.; Vondran, J.C.; Vanchina, M.A.; Jewell, D.E. Comparison of circulating metabolite concentrations in dogs and cats when allowed to freely choose macronutrient intake. Biol. Open 2018, 7. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Mean Values * | Group Effect ‡ | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| H Cats | CKD Cats | Overall Effect of Health | Overall Effect of Both Treatments | H Cats: Effect of Treatment with Food B versus Food A ¥ | CKD Cats: Effect of Treatment with Food B versus Food A ¥ | Overall Effect of Interaction | |||||
| Metabolites | BSL # | Food A | Food B | BSL # | Food A | Food B | P-Value | P-Value | P-Value | P-Value | P-Value |
| Tocopherol Metabolism | |||||||||||
| alpha-tocopherol | 1.11 | 1.00 § | 0.94 § | 1.03 | 0.99 | 0.93 § | 0.39 | 0.000 | 0.02 | 0.08 | 0.06 |
| alpha-tocopherol acetate | 0.94 | 1.09 | 0.89 | 0.83 | 1.02 § | 1.01 | 0.86 | 0.02 | 0.01 | 0.85 | 0.14 |
| delta-tocopherol | 1.31 | 0.97 § | 0.88 § | 1.23 | 0.96 § | 0.94 § | 0.91 | 0.000 | 0.02 | 0.74 | 0.09 |
| alpha-tocotrienol | 1.21 | 0.85 § | 1.02 § | 1.20 | 0.90 § | 1.07 | 0.56 | 0.000 | 0.003 | 0.000 | 0.26 |
| gamma-tocotrienol | 1.37 | 0.79 § | 0.85 § | 1.37 | 0.90 § | 1.00 § | 0.30 | 0.000 | 0.42 | 0.02 | 0.16 |
| gamma-CEHC | 1.51 | 1.20 § | 1.04 § | 0.93 | 0.67 | 0.68 | 0.04 | 0.01 | 0.69 | 0.78 | 0.66 |
| alpha-CEHC sulfate | 0.94 | 1.04 | 1.33 § | 0.63 | 0.64 | 0.88 | 0.15 | 0.03 | 0.13 | 0.66 | 0.68 |
| alpha-CEHC | 1.52 | 1.45 | 0.96 § | 0.97 | 0.83 | 0.59 | 0.10 | 0.01 | 0.02 | 0.23 | 0.71 |
| gamma-tocopherol/beta-tocopherol | 1.80 | 0.90 § | 0.97 § | 1.67 | 0.88 § | 1.02 § | 0.62 | 0.000 | 0.04 | 0.000 | 0.02 |
| Glutathione Metabolism | |||||||||||
| cysteinylglycine | 0.94 | 0.78 | 0.99 | 1.29 | 0.75 § | 1.00 § | 0.61 | 0.000 | 0.02 | 0.01 | 0.01 |
| 5-oxoproline | 1.59 | 1.80 | 1.61 | 1.49 | 1.19 | 1.79 | 0.30 | 0.53 | 0.63 | 0.22 | 0.43 |
| 2-hydroxybutyrate/2-hydroxyhydroxyisobutyrate | 0.86 | 1.25 | 1.38 | 0.86 | 0.93 | 1.85 | 0.66 | 0.04 | 0.91 | 0.04 | 0.31 |
| Sphingolipid Metabolism | |||||||||||
| sphinganine | 1.85 | 1.09 § | 1.04 § | 1.09 | 1.01 | 0.80 | 0.04 | 0.000 | 0.82 | 0.49 | 0.02 |
| sphingosine | 1.52 | 0.99 § | 0.96 § | 1.14 | 1.06 | 0.90 | 0.47 | 0.002 | 0.81 | 0.65 | 0.10 |
| sphingadienine | 1.11 | 0.93 § | 1.05 | 1.08 | 0.94 § | 1.07 | 0.94 | 0.000 | 0.03 | 0.004 | 0.57 |
| Glycine, Serine, and Threonine Metabolism | |||||||||||
| glycine | 0.90 | 1.20 § | 1.49 § | 0.84 | 0.78 | 0.85 | 0.01 | 0.003 | 0.08 | 0.36 | 0.01 |
| sarcosine | 0.96 | 1.03 | 1.20 | 0.85 | 0.77 | 1.02 | 0.18 | 0.53 | 0.49 | 0.59 | 0.96 |
| dimethylglycine | 1.06 | 1.38 § | 1.36 § | 0.80 | 1.11 | 1.49 § | 0.27 | 0.01 | 0.99 | 0.57 | 0.81 |
| betaine | 0.93 | 1.82 § | 2.56 § | 0.58 | 0.64 | 1.20 § | 0.003 | 0.000 | 0.14 | 0.01 | 0.08 |
| TCA Cycle | |||||||||||
| citrate | 0.90 | 1.24 | 1.63 | 0.94 | 0.89 | 1.01 | 0.18 | 0.43 | 0.76 | 0.46 | 0.52 |
| alpha-ketoglutarate | 0.91 | 1.79 | 1.19 | 0.98 | 1.04 | 1.69 § | 0.99 | 0.08 | 0.50 | 0.09 | 0.23 |
| succinate | 1.28 | 1.14 | 1.54 | 1.11 | 0.83 | 0.89 | 0.01 | 0.48 | 0.22 | 0.70 | 0.41 |
| fumarate | 1.49 | 1.00 | 0.89 | 1.54 | 1.25 | 1.35 | 0.72 | 0.27 | 0.75 | 0.56 | 0.44 |
| malate | 1.30 | 1.32 | 1.58 | 0.89 | 1.22 | 1.25 | 0.23 | 0.13 | 0.28 | 0.78 | 0.83 |
| Urea Cycle | |||||||||||
| arginine | 0.53 | 1.46 § | 1.35 § | 0.59 | 0.85 | 0.87 | 0.06 | 0.001 | 0.75 | 0.67 | 0.02 |
| argininosuccinate | 0.10 | 0.55 | 0.33 | 0.11 | 0.45 | 0.52 | 0.97 | 0.13 | 0.98 | 0.92 | 0.95 |
| ornithine | 1.21 | 1.21 | 1.06 | 1.08 | 1.11 | 0.99 | 0.23 | 0.65 | 0.42 | 0.74 | 0.78 |
| citrulline | 1.14 | 0.98 | 0.93 § | 1.07 | 1.03 | 0.93 | 0.98 | 0.05 | 0.68 | 0.29 | 0.62 |
| Valine, Arginine, and Proline Metabolism | |||||||||||
| norvaline | 0.72 | 2.34 § | 2.57 § | 0.79 | 1.44 § | 1.26 | 0.11 | 0.000 | 0.86 | 0.35 | 0.09 |
| 2-oxoarginine | 0.64 | 1.68 § | 1.32 § | 0.59 | 1.09 § | 1.45 § | 0.23 | 0.000 | 0.46 | 0.21 | 0.18 |
| homocitrulline | 0.85 | 1.11 § | 1.04 § | 0.98 | 1.59 § | 1.34 § | 0.42 | 0.000 | 0.79 | 0.99 | 0.15 |
| proline | 1.05 | 1.23 | 1.37 § | 1.35 | 0.88 § | 1.12 § | 0.46 | 0.06 | 0.33 | 0.12 | 0.001 |
| dimethylarginine (SDMA+ADMA) | 0.81 | 1.12 § | 1.11 § | 0.99 | 1.06 | 1.11 | 0.89 | 0.005 | 0.98 | 0.66 | 0.03 |
| N-acetylarginine | 0.80 | 1.24 § | 1.14 § | 0.86 | 1.04 | 0.98 | 0.34 | 0.02 | 0.64 | 0.55 | 0.20 |
| N-acetylcitrulline | 0.77 | 1.21 § | 1.15 § | 0.85 | 1.03 | 0.93 | 0.30 | 0.01 | 0.64 | 0.50 | 0.42 |
| N-acetylproline | 1.60 | 0.82 § | 0.82 § | 1.62 | 0.76 § | 0.71 § | 0.59 | 0.000 | 0.94 | 0.72 | 0.50 |
| N-delta-acetylornithine | 1.11 | 1.01 | 0.96 | 1.06 | 0.88 | 1.20 | 0.42 | 0.61 | 0.86 | 0.37 | 0.49 |
| N-alpha-acetylornithine | 1.24 | 1.17 | 0.98 | 1.14 | 0.93 | 0.85 § | 0.10 | 0.01 | 0.17 | 0.51 | 0.68 |
| N-methylhydroxyproline | 0.86 | 0.86 | 0.89 | 0.99 | 0.88 | 1.07 | 0.33 | 0.40 | 0.50 | 0.23 | 0.91 |
| trans-4-hydroxyproline | 0.75 | 1.31 § | 1.17 § | 0.98 | 0.99 | 1.08 | 0.32 | 0.10 | 0.58 | 0.66 | 0.03 |
| N-methylproline | 1.70 | 1.60 | 0.63 § | 1.68 | 1.47 | 0.75 | 0.94 | 0.002 | 0.001 | 0.16 | 0.38 |
| N-monomethylarginine | 0.82 | 0.78 | 1.17 § | 1.08 | 0.90 | 1.11 | 0.44 | 0.02 | 0.002 | 0.41 | 0.07 |
| argininate | 1.05 | 1.33 | 1.70 § | 0.86 | 0.82 | 0.93 | 0.01 | 0.20 | 0.18 | 0.59 | 0.11 |
| Mean Values * | Group Effect ‡ | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| H Cats | CKD Cats | Overall Effect of Health | Overall Effect of Both Treatments | H Cats: Effect of Treatment with Food B versus Food A ¥ | CKD Cats: Effect of Treatment with Food B versus Food A ¥ | Overall Effect of Interaction | |||||
| Metabolites | BSL # | Food A | Food B | BSL # | Food A | Food B | P-Value | P-Value | P-Value | P-Value | P-Value |
| Creatine Metabolism | |||||||||||
| guanidinoacetate | 1.83 | 1.07 | 0.84 § | 1.49 | 1.05 § | 1.43 | 0.84 | 0.02 | 0.46 | 0.28 | 0.42 |
| creatine | 0.98 | 1.16 | 1.54 | 1.42 | 0.76 | 1.45 | 0.26 | 0.20 | 0.37 | 0.33 | 0.78 |
| creatinine | 0.92 | 1.04 | 1.40 § | 1.10 | 0.72 | 1.38 | 0.26 | 0.16 | 0.24 | 0.19 | 0.38 |
| N-methylhydantoin | 1.00 | 1.24 | 1.53 | 0.88 | 0.26 § | 0.58 | 0.02 | 0.27 | 0.53 | 0.17 | 0.10 |
| Tryptophan Metabolism | |||||||||||
| tryptophan | 1.19 | 1.57 | 1.61 | 1.64 | 0.85 § | 0.83 § | 0.14 | 0.15 | 0.98 | 0.65 | 0.005 |
| N-acetyltryptophan | 1.25 | 1.21 | 1.24 | 1.67 | 0.85 § | 0.84 § | 0.36 | 0.000 | 0.79 | 0.76 | 0.02 |
| tryptophan betaine | 0.55 | 1.01 § | 1.03 § | 0.55 | 0.93 § | 0.92 § | 0.19 | 0.000 | 0.61 | 0.67 | 0.31 |
| kynurenine | 1.02 | 0.96 | 0.97 | 1.27 | 0.95 § | 1.10 | 0.74 | 0.02 | 0.96 | 0.19 | 0.17 |
| N-acetylkynurenine (2) | 0.79 | 0.96 | 1.38 § | 1.83 | 1.32 § | 1.34 | 0.36 | 0.07 | 0.08 | 0.12 | 0.02 |
| kynurenate | 1.03 | 0.95 | 0.92 | 1.20 | 1.07 | 1.30 | 0.30 | 0.26 | 0.59 | 0.06 | 0.22 |
| N-formylanthranilic acid | 1.59 | 1.04 § | 0.86 § | 1.61 | 1.32 § | 0.89 § | 0.81 | 0.000 | 0.15 | 0.14 | 0.81 |
| anthranilate | 1.12 | 0.82 | 0.74 | 1.45 | 0.75 | 0.70 § | 0.48 | 0.03 | 0.70 | 0.58 | 0.96 |
| xanthurenate | 1.25 | 1.22 | 0.88 § | 0.99 | 0.96 | 0.79 § | 0.05 | 0.000 | 0.001 | 0.03 | 0.61 |
| picolinate | 0.90 | 1.25 § | 0.98 | 1.10 | 1.08 | 0.96 | 0.87 | 0.08 | 0.04 | 0.62 | 0.10 |
| serotonin | 1.31 | 0.95 § | 0.83 § | 1.19 | 1.15 | 0.90 § | 0.65 | 0.000 | 0.14 | 0.07 | 0.02 |
| 5-hydroxyindoleacetate | 1.00 | 1.09 | 0.83 | 1.05 | 1.18 § | 0.90 | 0.59 | 0.02 | 0.01 | 0.14 | 0.25 |
| tryptamine | 1.85 | 0.93 | 0.76 § | 9.41 | 3.04 § | 6.76 | 0.05 | 0.001 | 0.41 | 0.08 | 0.15 |
| indolelactate | 0.87 | 2.28 § | 3.02 § | 1.28 | 0.79 § | 1.05 | 0.06 | 0.22 | 0.39 | 0.32 | 0.002 |
| indoleacetate | 1.17 | 0.85 | 0.89 | 0.69 | 0.93 § | 0.99 | 0.68 | 0.58 | 0.70 | 1.00 | 0.07 |
| indolepropionate | 1.77 | 1.94 | 1.77 | 2.57 | 0.85 | 0.85 | 0.70 | 0.05 | 0.18 | 0.77 | 0.74 |
| indole | 5.74 | 1.01 | 1.22 | 6.33 | 3.59 | 1.05 § | 0.46 | 0.05 | 0.91 | 0.42 | 0.45 |
| indole-3-carboxylic acid | 1.18 | 0.83 | 0.56 § | 0.95 | 1.06 | 0.81 | 0.53 | 0.24 | 0.39 | 0.83 | 0.09 |
| indoleacetylglycine | 0.99 | 1.15 | 2.34 § | 1.16 | 1.19 | 1.72 | 0.35 | 0.01 | 0.04 | 0.10 | 0.13 |
| 2-aminophenol | 2.38 | 0.93 § | 0.93 § | 2.62 | 0.86 § | 0.93 § | 0.62 | 0.000 | 0.96 | 0.89 | 0.14 |
| valeryltryptophan | 1.60 | 1.34 | 1.19 | 2.06 | 0.62 § | 0.60 § | 0.25 | 0.000 | 0.49 | 0.99 | 0.19 |
| Benzoate Metabolism | |||||||||||
| hippurate | 0.32 | 0.90 § | 1.85 § | 0.57 | 0.67 | 1.50 | 0.29 | 0.000 | 0.03 | 0.39 | 0.06 |
| 2-hydroxyhippurate (salicylurate) | 0.67 | 1.71 § | 2.07 § | 0.75 | 1.11 | 1.16 | 0.12 | 0.001 | 0.50 | 0.99 | 0.03 |
| 4-hydroxyhippurate | 0.84 | 1.39 § | 2.27 § | 1.23 | 1.11 | 0.98 | 0.09 | 0.02 | 0.03 | 0.78 | 0.01 |
| benzoate | 1.03 | 0.98 | 1.00 | 1.04 | 0.89 | 0.88 § | 0.25 | 0.12 | 0.79 | 0.43 | 0.29 |
| 4-hydroxybenzoate | 1.05 | 1.49 § | 1.37 § | 2.00 | 1.18 | 1.40 | 0.86 | 0.40 | 0.94 | 0.58 | 0.08 |
| catechol sulfate | 1.20 | 0.93 | 1.25 | 0.86 | 0.81 | 1.12 | 0.28 | 0.26 | 0.35 | 0.57 | 0.81 |
| 4-methylcatechol sulfate | 1.70 | 1.33 | 3.94 § | 1.36 | 0.80 | 2.93 § | 0.23 | 0.000 | 0.000 | 0.004 | 0.61 |
| p-cresol | 1.12 | 1.07 | 1.01 | 1.08 | 0.94 | 0.76 § | 0.31 | 0.03 | 0.42 | 0.25 | 0.66 |
| p-cresol sulfate | 1.31 | 0.84 | 1.17 | 0.87 | 0.75 | 0.79 | 0.24 | 0.71 | 0.20 | 0.92 | 0.60 |
| phenylpropionylglycine | 0.63 | 1.61 § | 2.99 § | 0.43 | 0.43 | 0.51 | 0.01 | 0.01 | 0.20 | 0.85 | 0.03 |
| 3-(3-hydroxyphenyl) propionate sulfate | 1.34 | 0.85 | 0.96 | 0.93 | 0.56 § | 0.73 | 0.33 | 0.09 | 0.52 | 0.76 | 0.63 |
| 2-(4-hydroxyphenyl) propionate | 0.45 | 0.80 § | 0.61 | 0.45 | 1.19 § | 0.83 § | 0.43 | 0.001 | 0.22 | 0.33 | 0.74 |
| 3-(3-hydroxyphenyl) propionate | 1.85 | 0.90 § | 0.75 § | 1.93 | 0.78 § | 0.86 | 0.60 | 0.000 | 0.67 | 0.62 | 0.56 |
| 3-(4-hydroxyphenyl) propionate | 1.29 | 1.04 | 1.07 | 1.10 | 1.02 | 1.22 | 0.64 | 0.27 | 0.96 | 0.16 | 0.61 |
| 3-phenylpropionate (hydrocinnamate) | 9.21 | 5.70 | 2.57 § | 7.63 | 16.56 | 11.69 | 0.83 | 0.14 | 0.09 | 0.37 | 0.01 |
| Mean Values * | Group Effect ‡ | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| H Cats | CKD Cats | Overall Effect of Health | Overall Effect of Both Treatments | H Cats: Effect of Treatment with Food B versus Food A ¥ | CKD Cats: Effect of Treatment with Food B versus Food A ¥ | Overall Effect of Interaction | |||||
| Metabolites | BSL # | Food A | Food B | BSL # | Food A | Food B | P-Value | P-Value | P-Value | P-Value | P-Value |
| Primary Bile Acid Metabolism | |||||||||||
| cholate | 1.41 | 1.10 | 1.20 | 1.41 | 0.94 § | 0.96 | 0.15 | 0.11 | 0.71 | 0.56 | 0.70 |
| glycocholate | 0.70 | 1.09 | 1.06 | 0.54 | 0.38 | 0.24 | 0.06 | 0.82 | 0.59 | 0.35 | 0.11 |
| taurocholate | 1.25 | 1.69 | 1.90 | 1.78 | 1.51 | 2.20 | 0.92 | 0.29 | 0.10 | 0.55 | 0.54 |
| chenodeoxycholate | 0.68 | 0.77 | 1.14 § | 0.74 | 0.74 | 1.11 § | 0.96 | 0.001 | 0.03 | 0.09 | 0.74 |
| taurochenodeoxycholate | 0.22 | 0.51 | 1.66 § | 0.61 | 0.68 | 2.50 § | 0.67 | 0.000 | 0.001 | 0.01 | 0.74 |
| beta-muricholate | 2.44 | 0.93 § | 0.79 § | 2.20 | 1.43 | 0.82 | 0.88 | 0.07 | 0.87 | 0.32 | 0.24 |
| cholate sulfate | 1.24 | 0.97 | 1.17 | 2.52 | 1.01 § | 1.16 | 0.35 | 0.05 | 0.64 | 0.15 | 0.12 |
| Secondary Bile Acid Metabolism | |||||||||||
| deoxycholate | 2.41 | 1.23 § | 0.87 § | 1.48 | 1.16 | 0.71 § | 0.52 | 0.000 | 0.05 | 0.12 | 0.59 |
| deoxycholic acid sulfate | 0.95 | 0.17 § | 0.43 | 0.82 | 0.11 § | 0.31 | 0.91 | 0.01 | 0.34 | 0.46 | 0.99 |
| taurodeoxycholate | 1.38 | 1.26 | 0.97 | 0.82 | 0.88 | 0.89 | 0.58 | 0.40 | 0.75 | 0.81 | 0.86 |
| lithocholate | 2.98 | 1.54 | 1.24 § | 2.19 | 2.15 | 1.39 | 0.88 | 0.13 | 0.32 | 0.76 | 0.33 |
| taurolithocholate 3-sulfate | 0.73 | 0.81 | 2.23 § | 2.51 | 1.36 | 2.73 | 0.61 | 0.003 | 0.01 | 0.05 | 0.62 |
| ursodeoxycholate | 1.10 | 0.87 | 1.02 | 1.27 | 1.35 | 1.19 | 0.49 | 0.55 | 0.13 | 0.91 | 0.25 |
| isoursodeoxycholate | 1.49 | 0.94 § | 0.78 § | 1.20 | 1.13 | 0.99 | 0.72 | 0.01 | 0.33 | 0.68 | 0.08 |
| tauroursodeoxycholate | 0.39 | 0.66 | 1.30 § | 0.51 | 0.55 | 1.33 § | 0.92 | 0.000 | 0.01 | 0.01 | 0.89 |
| dehydrolithocholate | 3.89 | 8.28 | 2.77 § | 2.97 | 1.51 | 4.38 | 0.77 | 0.62 | 0.23 | 0.26 | 0.10 |
| 7,12-diketolithocholate | 3.64 | 3.52 § | 1.14 § | 1.47 | 0.65 | 3.70 | 0.93 | 0.03 | 0.72 | 0.16 | 0.27 |
| 7-ketolithocholate | 1.68 | 0.86 | 1.09 | 1.18 | 1.30 | 2.16 | 0.32 | 0.46 | 0.33 | 0.43 | 0.99 |
| hyocholate | 0.68 | 0.94 § | 1.63 § | 0.56 | 0.73 | 1.69 § | 0.20 | 0.000 | 0.000 | 0.000 | 0.46 |
| 3-dehydrocholate | 2.18 | 1.33 | 1.28 | 1.26 | 0.86 | 1.33 | 0.19 | 0.14 | 0.90 | 0.46 | 0.65 |
| 12-dehydrocholate | 2.51 | 1.13 § | 1.42 | 1.22 | 0.92 | 1.29 | 0.07 | 0.02 | 0.60 | 0.20 | 0.76 |
| taurocholenate sulfate | 0.67 | 0.82 | 2.13 § | 1.54 | 0.82 | 2.02 | 0.77 | 0.002 | 0.01 | 0.03 | 0.40 |
| 7-ketodeoxycholate | 2.20 | 1.01 § | 0.94 § | 1.45 | 0.99 | 1.67 | 0.51 | 0.01 | 0.95 | 0.17 | 0.38 |
| 7alpha-hydroxycholestenone | 1.12 | 1.02 | 0.90 § | 1.07 | 1.05 | 1.04 | 0.56 | 0.03 | 0.03 | 0.88 | 0.08 |
| 3b-hydroxy-5-cholenoic acid | 1.45 | 1.11 | 1.06 | 1.03 | 1.20 | 1.09 | 0.67 | 0.84 | 0.60 | 0.99 | 0.12 |
| taurochenodeoxycholate sulfate | 0.63 | 0.68 | 1.51 § | 0.55 | 1.03 | 1.54 § | 0.75 | 0.01 | 0.14 | 0.40 | 0.89 |
| ursodeoxycholate sulfate (1) | 3.37 | 0.77 § | 1.99 | 4.00 | 0.66 § | 1.91 | 0.71 | 0.01 | 0.69 | 0.07 | 0.58 |
| ursocholate | 1.35 | 1.09 | 1.16 | 1.98 | 1.11 | 1.33 | 0.39 | 0.11 | 0.68 | 0.46 | 0.89 |
| Cats | Healthy | CKD |
|---|---|---|
| Age, years | 7.2 (1.3) | 8.0 (1.8) |
| Sex | 8 spayed females, 2 neutered males | 5 spayed females, 5 neutered males |
| Body weight, kg | 4.55 (0.68) | 5.40 (0.85) |
| Nutrient | Pre-Trial Food | Food A with scFOS | Food B with Apple Pomace |
|---|---|---|---|
| Moisture | 5.47 | 5.34 | 5.97 |
| Protein | 27.6 | 28.0 | 27.3 |
| Fat | 19.9 | 19.5 | 19.5 |
| Atwater Energy, 4 kcal/kg | 4101 | 4063 | 4032 |
| Ash | 4.50 | 4.42 | 4.59 |
| Crude Fiber | 1.3 | 2.0 | 2.1 |
| Insoluble Fiber | 4.0 | 3.4 | 5.1 |
| Soluble Fiber | 1.3 | 0.8 | 1.6 |
| Total Dietary Fiber | 5.3 | 4.2 | 6.7 |
| Calcium | 0.73 | 0.72 | 0.79 |
| Phosphorus | 0.64 | 0.51 | 0.47 |
| Sodium | 0.24 | 0.22 | 0.22 |
| ARA [20:4 (n−6)] | 0.04 | 0.04 | 0.04 |
| EPA [20:5 (n−3)] | <0.01 | 0.02 | 0.02 |
| DHA [22:6 (n−3)] | 0.01 | 0.03 | 0.02 |
| SFA 5 | 6.32 | 6.59 | 6.72 |
| MUFA 6 | 7.40 | 8.04 | 8.24 |
| PUFA 7 | 9.58 | 7.83 | 8.25 |
| Total FA | 18.56 | 18.61 | 19.16 |
| (n−6) FA 8 | 4.10 | 3.70 | 3.90 |
| (n−3) FA 9 | 0.69 | 0.22 | 0.23 |
| (n−6):(n−3) ratio | 5.9 | 16.8 | 17.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, J.A.; Jewell, D.E.; Ephraim, E. Changes in the Fecal Metabolome Are Associated with Feeding Fiber Not Health Status in Cats with Chronic Kidney Disease. Metabolites 2020, 10, 281. https://doi.org/10.3390/metabo10070281
Hall JA, Jewell DE, Ephraim E. Changes in the Fecal Metabolome Are Associated with Feeding Fiber Not Health Status in Cats with Chronic Kidney Disease. Metabolites. 2020; 10(7):281. https://doi.org/10.3390/metabo10070281
Chicago/Turabian StyleHall, Jean A., Dennis E. Jewell, and Eden Ephraim. 2020. "Changes in the Fecal Metabolome Are Associated with Feeding Fiber Not Health Status in Cats with Chronic Kidney Disease" Metabolites 10, no. 7: 281. https://doi.org/10.3390/metabo10070281
APA StyleHall, J. A., Jewell, D. E., & Ephraim, E. (2020). Changes in the Fecal Metabolome Are Associated with Feeding Fiber Not Health Status in Cats with Chronic Kidney Disease. Metabolites, 10(7), 281. https://doi.org/10.3390/metabo10070281