Investigations on Metabolic Changes in Beagle Dogs Fed Probiotic Queso Blanco Cheese and Identification of Candidate Probiotic Fecal Biomarkers Using Metabolomics Approaches

 ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
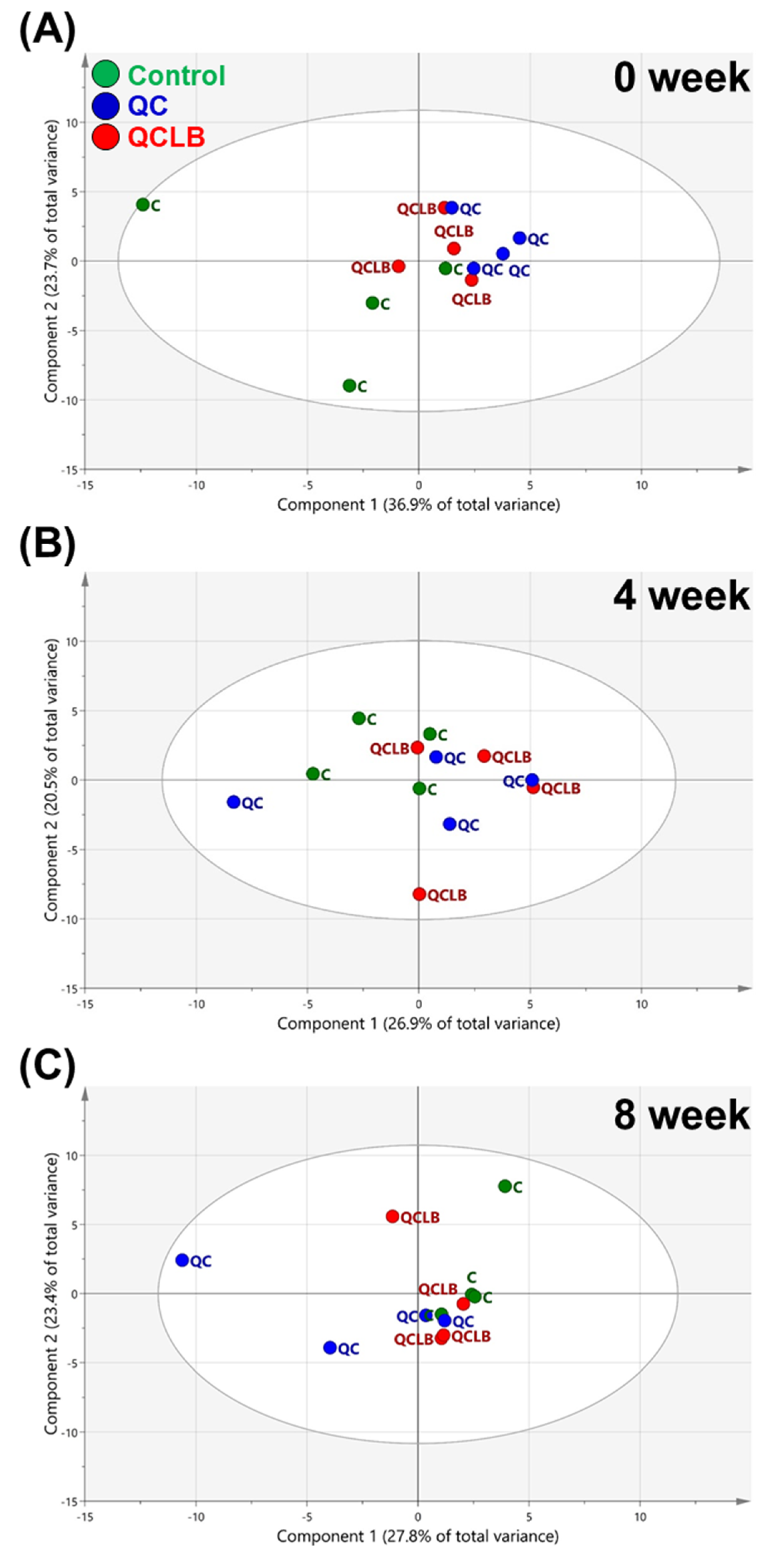
2.1. Comparison of Fecal Metabolic Profiles between the Three Dietary Groups
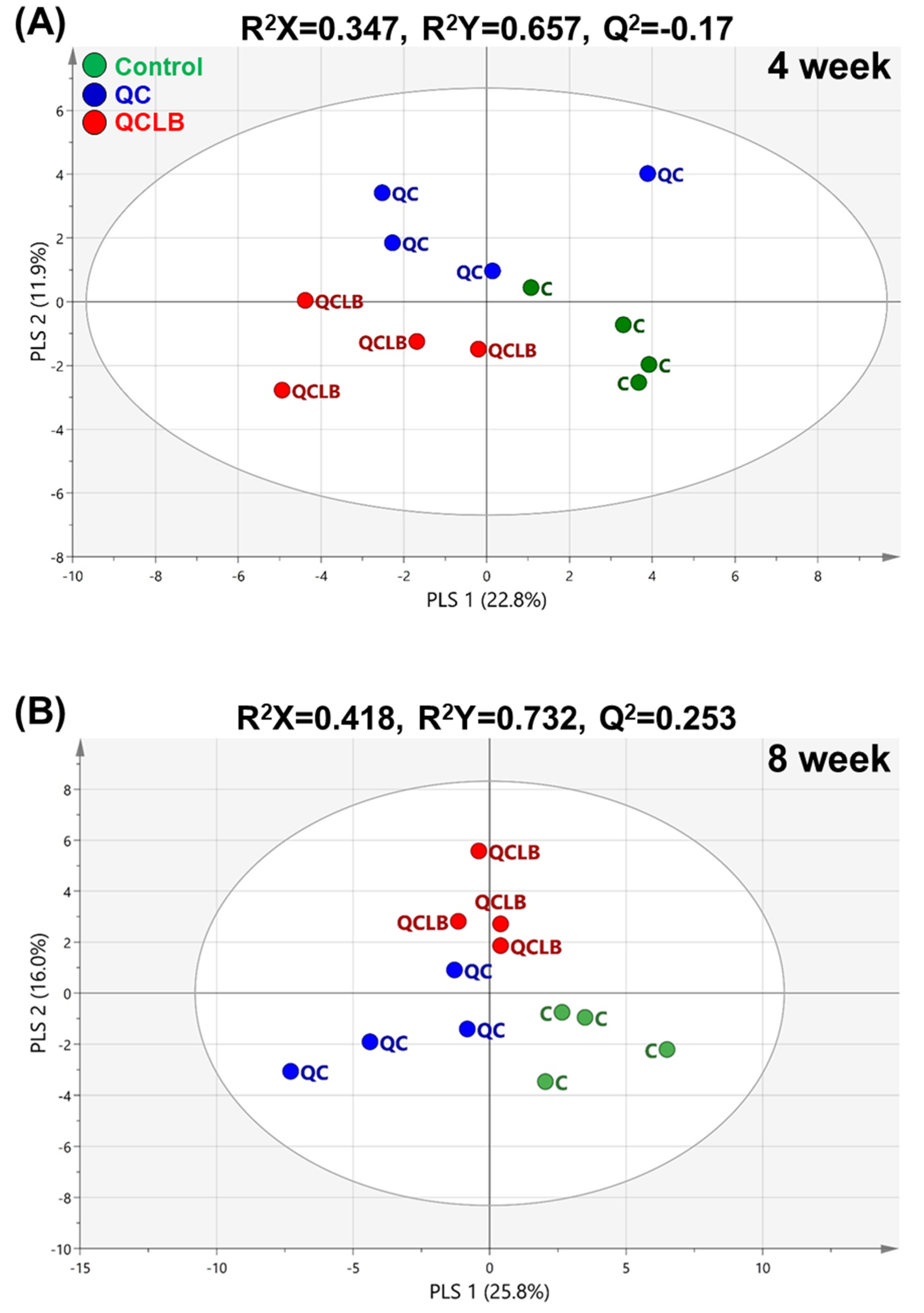
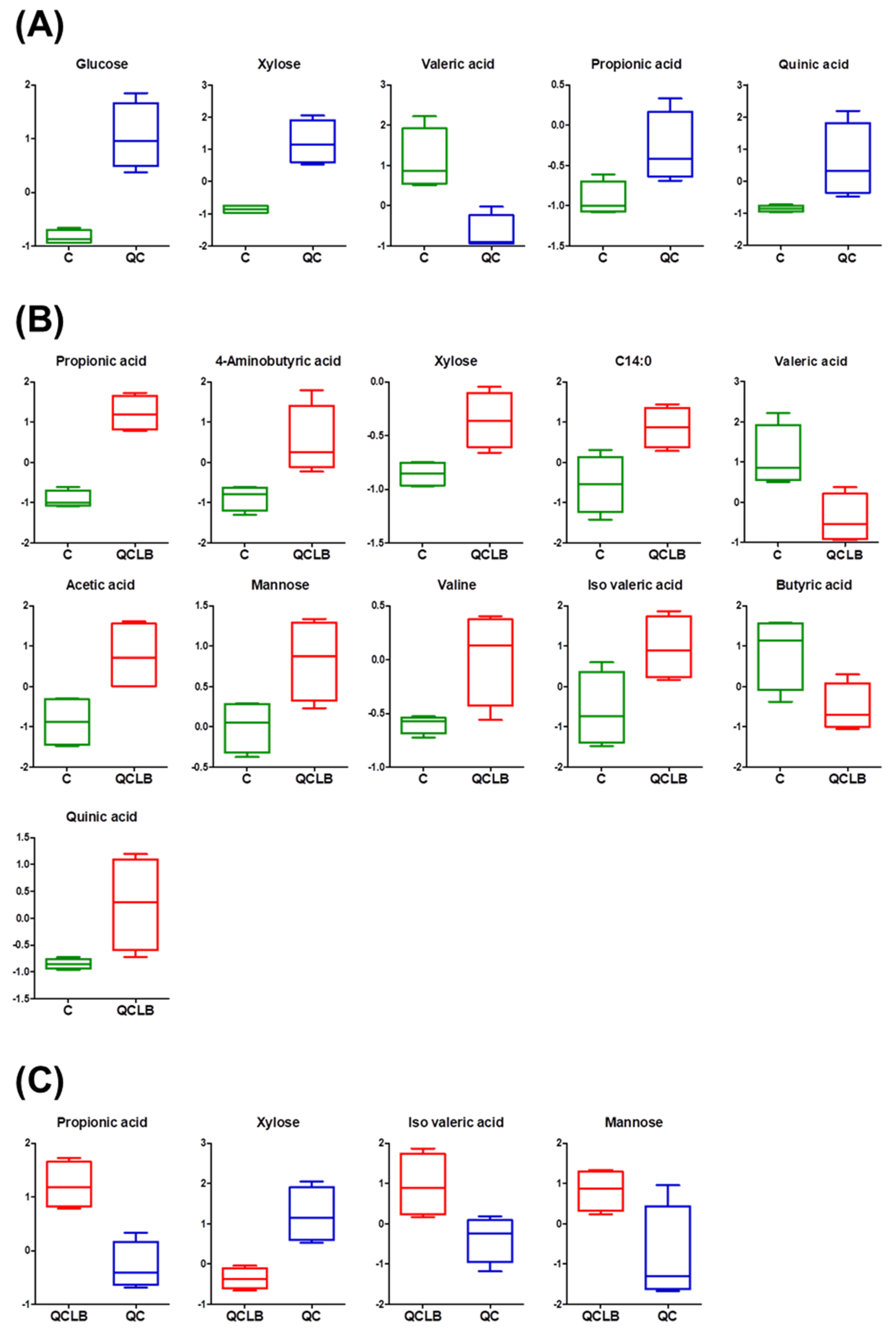
2.2. Identification of Potential Fecal Biomarkers Related to Metabolic Changes Induced by Probiotic Cheese Ingestion in Beagle Dogs
2.3. Visualization of Dynamic Metabolic Changes in Feces after Probiotic Cheese Ingestion
3. Materials and Methods
3.1. Preparation of Cheese
3.2. Animals and Experimental Design
3.3. Analysis of Volatile Fatty Acids and Indolic Compounds in Feces of Healthy Beagle Dogs
3.4. Analysis of Long Chain Fatty Acids in Feces of Healthy Beagle Dogs
3.5. Analysis of Hydrophilic Compounds in Feces of Healthy Beagle Dogs
3.6. Statistical Analysis
4. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics (isapp) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linares, D.M.; Gómez, C.; Renes, E.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Stanton, C. Lactic acid bacteria and bifidobacteria with potential to design natural biofunctional health–promoting dairy foods. Front. Microbiol. 2017, 8, 846. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Dis. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; Ross, P.; Stanton, C. Beneficial microbes: The pharmacy in the gut. Bioengineered 2016, 7, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenorio–Jiménez, C.; Martínez-Ramírez, M.J.; Tercero-Lozano, M.; Arraiza–Irigoyen, C.; Del Castillo-Codes, I.; Olza, J.; Plaza-Díaz, J.; Fontana, L.; Migueles, J.H.; Olivares, M.; et al. Evaluation of the effect of Lactobacillus reuteri V3401 on biomarkers of inflammation, cardiovascular risk and liver steatosis in obese adults with metabolic syndrome: A randomized clinical trial (PROSIR). BMC Complement. Altern. Med. 2018, 18, 306. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Y.; Jeffrey, M.P.; Johnson–Henry, K.C.; Green–Johnson, J.M.; Sherman, P.M. Impact of prebiotics, probiotics, and gut derived metabolites on host immunity. Lympho Sign J. 2017, 4, 1–24. [Google Scholar] [CrossRef]
- Elian, S.D.A.; Souza, E.L.S.; Vieira, A.T.; Teixeira, M.M.; Arantes, R.M.E.; Nicoli, J.R.; Martins, F.S. Bifidobacterium Longum Subsp. Infantis BB-02 Attenuates Acute Murine Experimental Model of Inflammatory Bowel Disease. Benef. Microbes 2015, 6, 277–286. [Google Scholar] [CrossRef]
- Tomosada, Y.; Villena, J.; Murata, K.; Chiba, E.; Shimazu, T.; Aso, H.; Iwabuchi, N.; Xiao, J.-Z.; Saito, T.; Kitazawa, H. Immunoregulatory effect of bifidobacteria strains in porcine intestinal epithelial cells through modulation of ubiquitin–editing enzyme A20 expression. PLOS ONE 2013, 8, e59259. [Google Scholar] [CrossRef]
- Mathipa, M.G.; Thantsha, M.S. Probiotic engineering: Towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 28. [Google Scholar] [CrossRef]
- Karimi, R.; Mortazavian, A.M.; Karami, M. Incorporation of Lactobacillus casei in Iranian ultrafiltered Feta cheese made by partial replacement of NaCl with KCl. J. Dairy Sci. 2012, 95, 4209–4222. [Google Scholar] [CrossRef]
- Lahtinen, S.J.; Forssten, S.; Aakko, J.; Granlund, L.; Rautonen, N.; Salminen, S.; Viitanen, M.; Ouwehand, A.C. Probiotic cheese containing Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus NCFM® modifies subpopulations of fecal lactobacilli and Clostridium difficile in the elderly. Age 2012, 34, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Coelho, L.P.; Kultima, J.R.; Costea, P.I.; Fournier, C.; Pan, Y.; Czarnecki-Maulden, G.; Hayward, M.R.; Forslund, S.K.; Schmidt, T.S.B.; Descombes, P.; et al. Similarity of the dog and human gut microbiomes in gene content and response to diet. Microbiome 2018, 6, 72. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, F.; Hou, Q.; Huang, W.; Liu, Y.; Zhang, H.; Sun, Z. Metagenomic analysis revealed beneficial effects of probiotics in improving the composition and function of the gut microbiota in dogs with diarrhoea. Food Funct. 2019, 10, 2618–2629. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Sim, J.H.; Min, T.S.; Choi, H.K. Metabolomics and Lipidomics Approaches in the Science of Probiotics: A Review. J. Med. Food 2018, 21, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Tarbah, F.A.; Mahler, H.; Temme, O.; Daldrup, T. An analytical method for the rapid screening of organophosphate pesticides in human biological samples and foodstuffs. Forensic Sci. Int. 2001, 121, 126–133. [Google Scholar] [CrossRef]
- Lotti, C.; Rubert, J.; Fava, F.; Tuohy, K.; Mattivi, F.; Vrhovsek, U. Development of a fast and cost–effective gas chromatography–mass spectrometry method for the quantification of short–chain and medium–chain fatty acids in human biofluids. Anal. Bioanal. Chem. 2017, 409, 5555–5567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Z.; Liu, O. Development and validation of a GC–FID method for quantitative analysis of oleic acid and related fatty acids. J. Pharm. Anal. 2015, 5, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, W.J.; Koal, T.; Wang, Y.; Kohl, M.; Enot, D.P.; Deigner, H.P. Targeted metabolomics for biomarker discovery. Angew. Chem. Int. Ed. 2010, 49, 5426–5445. [Google Scholar] [CrossRef]
- Wishart, D.S. Metabolomics: Applications to food science and nutrition research. Trends Food Sci. Technol. 2008, 19, 482–493. [Google Scholar] [CrossRef]
- Mozzi, F.; Ortiz, M.E.; Bleckwedel, J.; De Vuyst, L.; Pescuma, M. Metabolomics as a tool for the comprehensive understanding of fermented and functional foods with lactic acid bacteria. Food Res. Int. 2013, 54, 1152–1161. [Google Scholar] [CrossRef]
- Park, H.-E.; Kim, Y.J.; Do, K.-H.; Kim, J.K.; Ham, J.-S.; Lee, W.-K. Effects of queso blanco cheese containing Bifidobacterium longum KACC 91563 on the intestinal microbiota and short chain fatty acid in healthy companion dogs. Korean J. Food Sci. Anim. Resour. 2018, 38, 1261. [Google Scholar] [CrossRef] [PubMed]
- Park, H.E.; Kim, Y.J.; Kim, M.; Kim, H.; Do, K.H.; Kim, J.K.; Ham, J.-S.; Lee, W.K. Effects of Queso Blanco cheese containing Bifidobacterium longum KACC 91563 on fecal microbiota, metabolite and serum cytokine in healthy beagle dogs. Anaerobe 2020, 102234. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Parnell-Clunies, E.M.; Irvine, D.; Bullock, D. Composition and yield studies for Queso Blanco made in pilot plant and commercial trials with dilute acidulant solutions. J. Dairy Sci. 1985, 68, 3095–3103. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Taylor, T.D.; Ohno, H.; Hattori, M. Acetate-producing bifidobacteria protect the host from enteropathogenic infection via carbohydrate transporters. Gut Microbes 2012, 3, 449–454. [Google Scholar] [CrossRef] [Green Version]
- da Cruz, A.G.; Buriti, F.C.A.; de Souza, C.H.B.; Faria, J.A.F.; Saad, S.M.I. Probiotic cheese: Health benefits, technological and stability aspects. Trends Food Sci. Technol. 2009, 20, 344–354. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.; Lugo, M.; Major, A.; Mori-Akiyama, Y. GABA–producing Bifidobacterium dentium modulates visceral sensitivity in the intestine. Neurogastroenterol. Motil. 2017, 29, e12904. [Google Scholar] [CrossRef]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ–aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [Green Version]
- Thierry, A.; Maillard, M.-B. Production of cheese flavour compounds derived from amino acid catabolism by Propionibacterium freudenreichii. Lait 2002, 82, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Mäkeläinen, H.; Forssten, S.; Olli, K.; Granlund, L.; Rautonen, N.; Ouwehand, A. Probiotic lactobacilli in a semi-soft cheese survive in the simulated human gastrointestinal tract. Int. Dairy J. 2009, 19, 675–683. [Google Scholar] [CrossRef]
- Elsden, S.R.; Hilton, M.G. Volatile acid production from threonine, valine, leucine and isoleucine by clostridia. Arch. Microbiol. 1978, 117, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hou, C.; Wang, G.; Jia, H.; Yu, H.; Zeng, X.; Thacker, P.A.; Zhang, G.; Qiao, S. Lactobacillus reuteri I5007 modulates intestinal host defense peptide expression in the model of IPEC–J2 cells and neonatal piglets. Nutrients 2017, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staudigl, P.; Haltrich, D.; Peterbauer, C.K. L-Arabinose isomerase and D-xylose isomerase from Lactobacillus reuteri: Characterization, coexpression in the food grade host Lactobacillus plantarum, and application in the conversion of D–galactose and D-glucose. J. Agric. Food Chem. 2014, 62, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhao, G.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Production of exopolysaccharide by Bifidobacterium longum isolated from elderly and infant feces and analysis of priming glycosyltransferase genes. RSC Adv. 2017, 7, 31736–31744. [Google Scholar] [CrossRef] [Green Version]
- Ham, J.S.; Jeong, S.G.; Noh, Y.B.; Shin, J.H.; Han, G.S.; Chae, H.S.; Yoo, Y.M.; Ahn, J.N.; Lee, J.W.; Jo, C.; et al. Effects of gamma irradiation on Queso Blanco cheese. Korean J. Dairy Sci. Technol. 2007, 25, 15–20. [Google Scholar]
- Kim, Y.J.; Kim, J.G.; Lee, W.-K.; So, K.M.; Kim, J.K. Trial data of the anti–obesity potential of a high resistant starch diet for canines using Dodamssal rice and the identification of discriminating markers in feces for metabolic profiling. Metabolomics 2019, 15, 21. [Google Scholar] [CrossRef]
- Zheng, S.; Yu, M.; Lu, X.; Huo, T.; Ge, L.; Yang, J.; Wu, C.; Li, F. Urinary metabonomic study on biochemical changes in chronic unpredictable mild stress model of depression. Clin. Chim. Acta. 2010, 411, 204–209. [Google Scholar] [CrossRef]
- Eriksson, L.; Byrne, T.; Johansson, E.; Trygg, J.; Vikström, C. Multi–and Megavariate Data Analysis Basic Principles and Applications, 3rd ed.; Umetrics Academy: Umeå, Sweden, 2013; Volume 1. [Google Scholar]
- Oberg, J.; Spenger, C.; Wang, F.-H.; Andersson, A.; Westman, E.; Skoglund, P.; Sunnemark, D.; Norinder, U.; Klason, T.; Wahlund, L.-O. Age related changes in brain metabolites observed by 1H MRS in APP/PS1 mice. Neurobiol. Aging 2008, 29, 1423–1433. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Park, H.-E.; Lee, W.-K.; Ham, J.-S.; Park, S.U.; Kim, J.G.; Im, K.-H.; Kim, J.K. Investigations on Metabolic Changes in Beagle Dogs Fed Probiotic Queso Blanco Cheese and Identification of Candidate Probiotic Fecal Biomarkers Using Metabolomics Approaches. Metabolites 2020, 10, 305. https://doi.org/10.3390/metabo10080305
Kim YJ, Park H-E, Lee W-K, Ham J-S, Park SU, Kim JG, Im K-H, Kim JK. Investigations on Metabolic Changes in Beagle Dogs Fed Probiotic Queso Blanco Cheese and Identification of Candidate Probiotic Fecal Biomarkers Using Metabolomics Approaches. Metabolites. 2020; 10(8):305. https://doi.org/10.3390/metabo10080305
Chicago/Turabian StyleKim, Ye Jin, Ho-Eun Park, Wan-Kyu Lee, Jun-Sang Ham, Sang Un Park, Jae Geun Kim, Kyung-Hoan Im, and Jae Kwang Kim. 2020. "Investigations on Metabolic Changes in Beagle Dogs Fed Probiotic Queso Blanco Cheese and Identification of Candidate Probiotic Fecal Biomarkers Using Metabolomics Approaches" Metabolites 10, no. 8: 305. https://doi.org/10.3390/metabo10080305