
Lipidomics-Based Comparison of Molecular Compositions of Green, Yellow, and Red Bell Peppers

 ,
,
Abstract
1. Introduction
2. Results
2.1. Metabolomics Reveals Several Lipid Compounds in Pepper
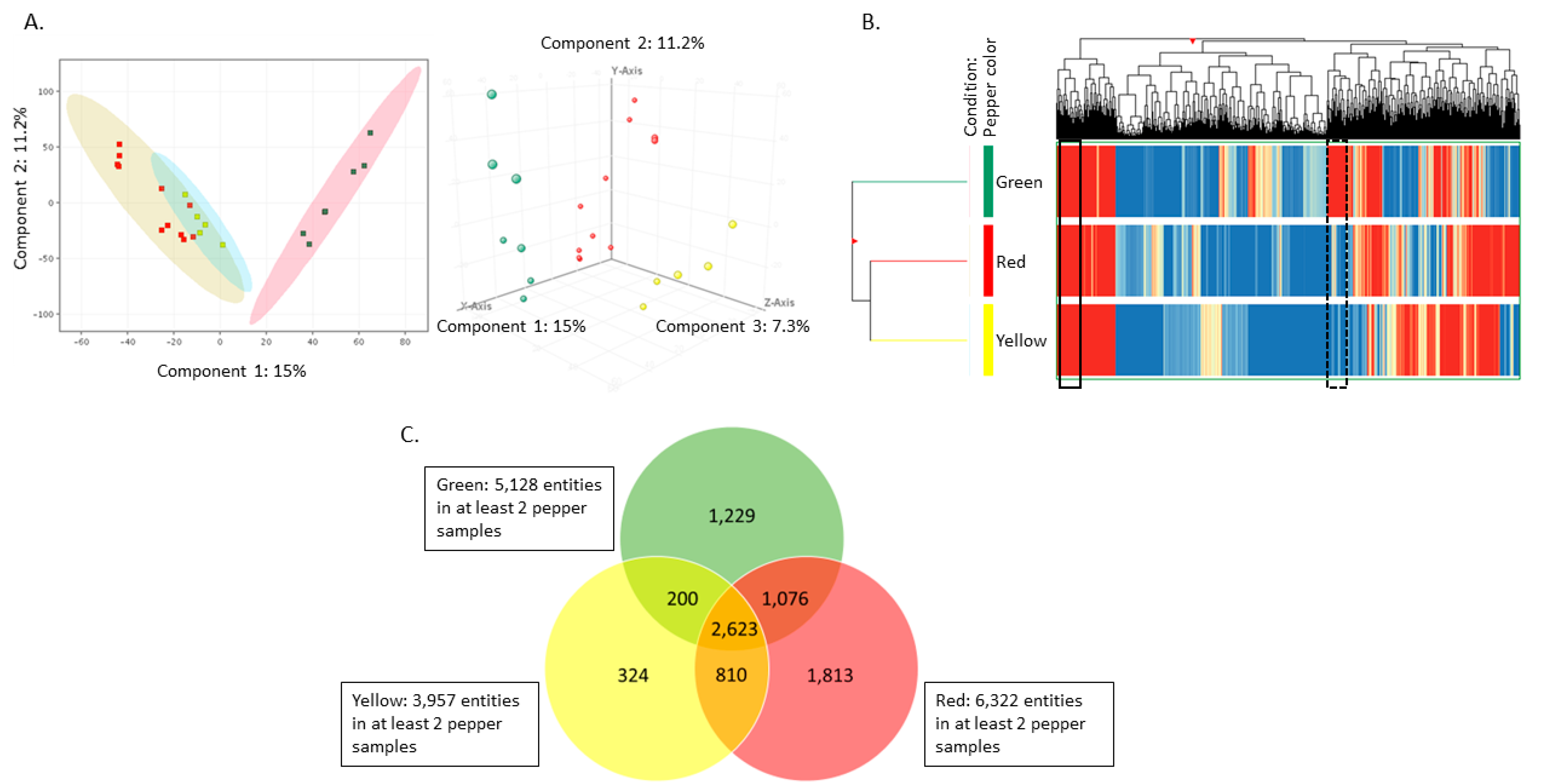
2.2. Principal Component Analysis (PCA) and Hierarchical Clustering (HC) Illustrate Pepper Samples Group Together Based on Color
2.3. Untargeted Lipidomics Reveals Several Compounds That Differ between Green, Red, and Yellow Peppers
2.4. Pairwise Comparisons of Green, Yellow, and Red Peppers
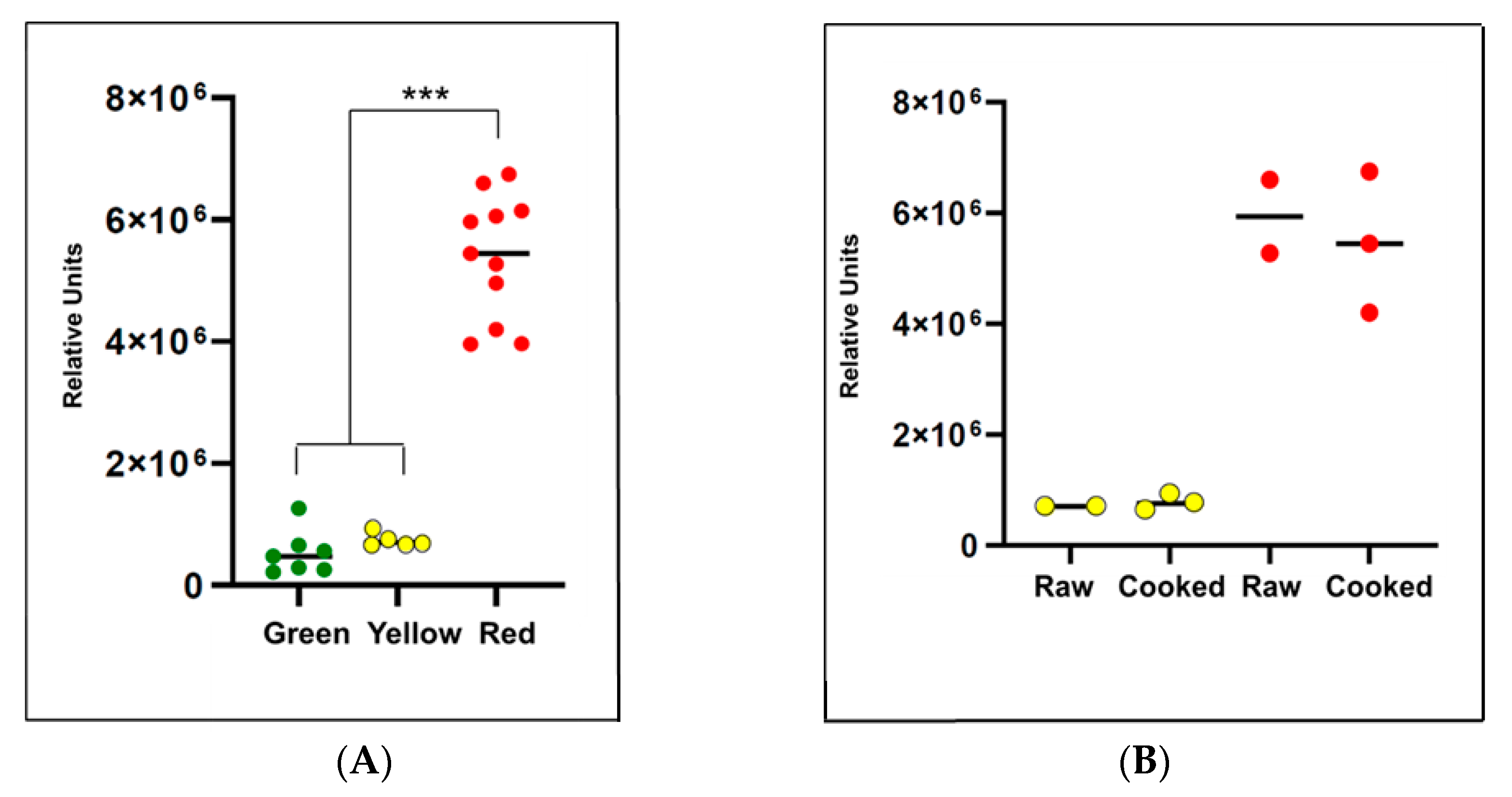
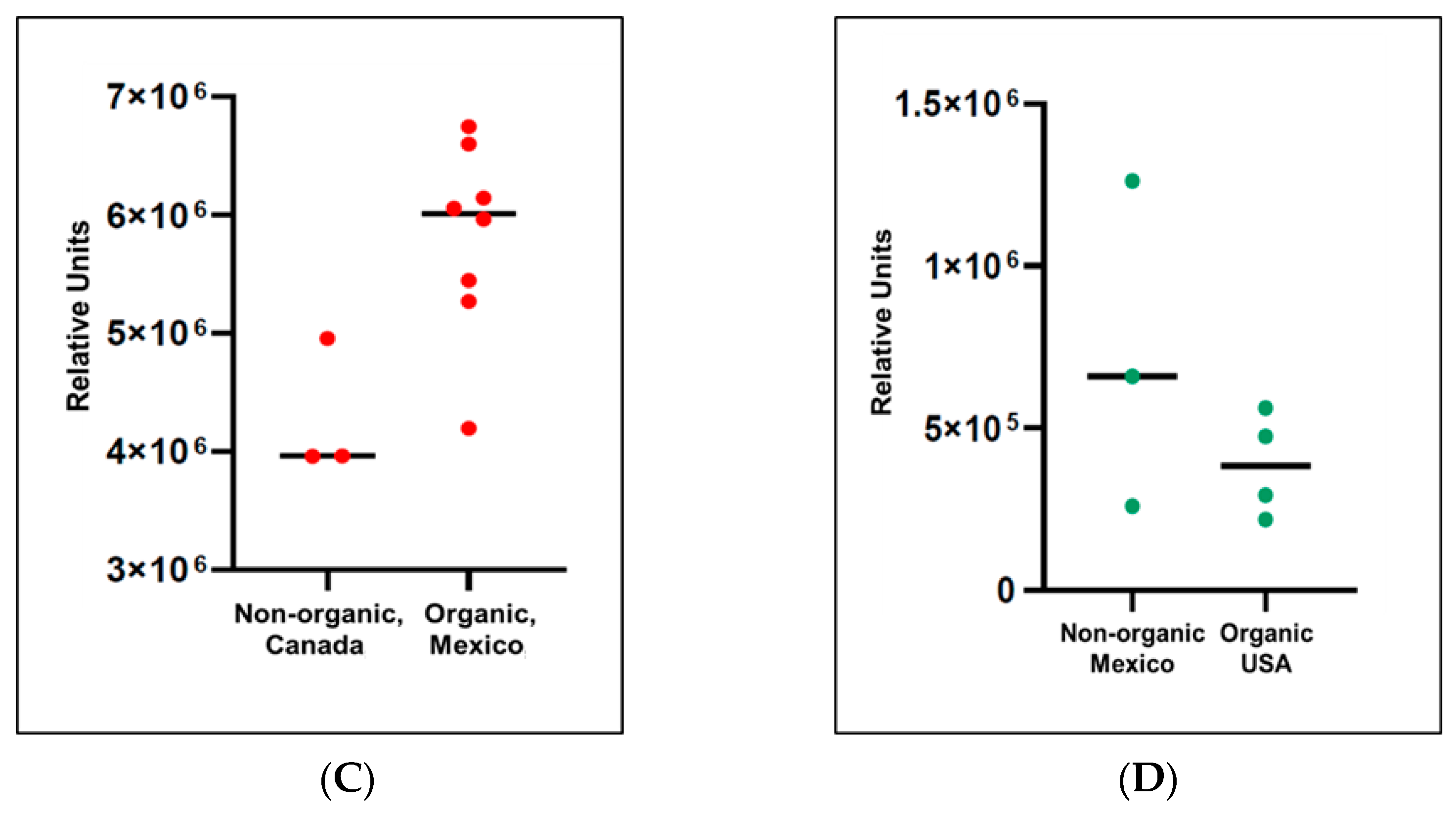
2.5. Evaluation of Pepper Attributes for Beta-Cryptoxanthin
3. Discussion
4. Materials and Methods
4.1. Chemicals, Standards, and Reagents
4.2. Bell Pepper Sample Preparation
4.3. Liquid Chromatography (LC) Mass Spectrometry (MS)
4.4. Mass Spectrometry (MS)
4.5. Data Processing
4.6. Compound Annotation
4.7. Tandem MS (MS/MS)
4.8. Statistical Analysis
4.9. Data Visualizations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldberg, J.P.; Sliwa, S.A. Communicating actionable nutrition messages: Challenges and opportunities. Proc. Nutr. Soc. 2011, 70, 26–37. [Google Scholar] [CrossRef]
- Ulaszewska, M.M.; Weinert, C.H.; Trimigno, A.; Portmann, R.; Lacueva, C.A.; Badertscher, R.; Brennan, L.; Brunius, C.; Bub, A.; Capozzi, F.; et al. Nutrimetabolomics: An Integrative Action for Metabolomic Analyses in Human Nutritional Studies. Mol. Nutr. Food Res. 2019, 63, e1800384. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Hu, F.B. Metabolomics-Based Dietary Biomarkers in Nutritional Epidemiology-Current Status and Future Opportunities. Mol. Nutr. Food Res. 2018, 63, e1701064. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, F.; Bordoni, A. Foodomics: A new comprehensive approach to food and nutrition. Genes Nutr. 2012, 8, 1–4. [Google Scholar] [CrossRef]
- Illner, A.-K.; Freisling, H.; Boeing, H.; Huybrechts, I.; Crispim, S.P.; Slimani, N. Review and evaluation of innovative technologies for measuring diet in nutritional epidemiology. Int. J. Epidemiol. 2012, 41, 1187–1203. [Google Scholar] [CrossRef]
- Jenab, M.; Slimani, N.; Bictash, M.; Ferrari, P.; Bingham, S.A. Biomarkers in nutritional epidemiology: Applications, needs and new horizons. Qual. Life Res. 2009, 125, 507–525. [Google Scholar] [CrossRef]
- Maruvada, P.; Lampe, J.W.; Wishart, D.S.; Barupal, D.; Chester, D.N.; Dodd, D.; Djoumbou-Feunang, Y.; Dorrestein, P.C.; Dragsted, L.O.; Draper, J.; et al. Perspective: Dietary Biomarkers of Intake and Exposure—Exploration with Omics Approaches. Adv. Nutr. 2019, 11, 200–215. [Google Scholar] [CrossRef] [PubMed]
- McNamara, A.E.; Brennan, L. Potential of food intake biomarkers in nutrition research. Proc. Nutr. Soc. 2020, 79, 487–497. [Google Scholar] [CrossRef]
- Reisdorph, N.A.; Hendricks, A.E.; Tang, M.; Doenges, K.A.; Reisdorph, R.M.; Tooker, B.C.; Quinn, K.; Borengasser, S.J.; Nkrumah-Elie, Y.; Frank, D.N.; et al. Nutrimetabolomics reveals food-specific compounds in urine of adults consuming a DASH-style diet. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; Sanchez, E.; Muñoz-Marquez, E.; Sida-Arreola, J.P.; Flores-Cordova, M.A. Bioactive Compounds and Antioxidant Activity in Different Grafted Varieties of Bell Pepper. Antioxidants 2015, 4, 427–446. [Google Scholar] [CrossRef]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in Phytochemical and Antioxidant Activity of Selected Pepper Cultivars (CapsicumSpecies) As Influenced by Maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorsi, I.; Cacciola, F.; Utczas, M.; Inferrera, V.; Giuffrida, D.; Donato, P.; Dugo, P.; Mondello, L. Characterization of the pigment fraction in sweet bell peppers (Capsicum annuumL.) harvested at green and overripe yellow and red stages by offline multidimensional convergence chromatography/liquid chromatography-mass spectrometry. J. Sep. Sci. 2016, 39, 3281–3291. [Google Scholar] [CrossRef]
- Delgado-Vargas, F.; Jiménez, A.R.; Paredes-López, O. Natural Pigments: Carotenoids, Anthocyanins, and Betalains—Characteristics, Biosynthesis, Processing, and Stability. Crit. Rev. Food Sci. Nutr. 2000, 40, 173–289. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.-R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Secondary Metabolites of Capsicum Species and Their Importance in the Human Diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar] [CrossRef]
- Hassan, N.M.; Yusof, N.A.; Yahaya, A.F.; Rozali, N.N.M.; Othman, R. Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants 2019, 8, 469. [Google Scholar] [CrossRef]
- Kim, T.J.; Choi, J.; Kim, K.W.; Kil Ahn, S.; Ha, S.-H.; Choi, Y.; Park, N.I.; Kim, J.K. Metabolite Profiling of Peppers of Various Colors Reveals Relationships Between Tocopherol, Carotenoid, and Phytosterol Content. J. Food Sci. 2017, 82, 2885–2893. [Google Scholar] [CrossRef] [PubMed]
- Thuphairo, K.; Sornchan, P.; Suttisansanee, U. Bioactive Compounds, Antioxidant Activity and Inhibition of Key Enzymes Relevant to Alzheimer’s Disease from Sweet Pepper (Capsicum annuum) Extracts. Prev. Nutr. Food Sci. 2019, 24, 327–337. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Chemical Analysis Working Group (CAWG). Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Allison, P.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Harrington, R.A.; Adhikari, V.; Rayner, M.; Scarborough, P. Nutrient composition databases in the age of big data: FoodDB, a comprehensive, real-time database infrastructure. BMJ Open 2019, 9, e026652. [Google Scholar] [CrossRef]
- Wang, Y.; Bryant, S.H.; Cheng, T.; Wang, J.; Gindulyte, A.; Shoemaker, B.A.; Thiessen, P.A.; He, S.; Zhang, J. PubChem BioAssay: 2017 update. Nucleic Acids Res. 2017, 45, D955–D963. [Google Scholar] [CrossRef] [PubMed]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Amin, A.U.; Darusman, L.K.; et al. KNApSAcK Family Databases: Integrated Metabolite–Plant Species Databases for Multifaceted Plant Research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Miller, N.J.; Sampson, J.; Candeias, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef]
- Burri, B.J.; Chang, J.S.T.; Neidlinger, T.R. β-Cryptoxanthin- and α-carotene-rich foods have greater apparent bioavailability than β-carotene-rich foods in Western diets. Br. J. Nutr. 2010, 105, 212–219. [Google Scholar] [CrossRef]
- Ho, Y.-S.; Chen, C.-H.; Wang, Y.-J.; Pestell, R.G.; Albanese, C.; Chen, R.; Chang, M.-C.; Jeng, J.-H.; Lin, S.-Y.; Liang, Y.-C.; et al. Tobacco-specific carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) induces cell proliferation in normal human bronchial epithelial cells through NFκB activation and cyclin D1 up-regulation. Toxicol. Appl. Pharmacol. 2005, 205, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Min, J. Serum carotenoid levels and risk of lung cancer death in US adults. Cancer Sci. 2014, 105, 736–743. [Google Scholar] [CrossRef]
- Ni, Y.; Nagashimada, M.; Zhan, L.; Nagata, N.; Kobori, M.; Sugiura, M.; Ogawa, K.; Kaneko, S.; Ota, T. Prevention and Reversal of Lipotoxicity-Induced Hepatic Insulin Resistance and Steatohepatitis in Mice by an Antioxidant Carotenoid, β-Cryptoxanthin. Endocrinol. 2015, 156, 987–999. [Google Scholar] [CrossRef]
- Ryan, L.; O’Connell, O.; O’Sullivan, L.; Aherne, S.A.; O’Brien, N.M. Micellarisation of Carotenoids from Raw and Cooked Vegetables. Plant Foods Hum. Nutr. 2008, 63, 127–133. [Google Scholar] [CrossRef]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In vitro investigation of the bioaccessibility of carotenoids from raw, frozen and boiled red chili peppers (Capsicum annuum). Eur. J. Nutr. 2013, 53, 501–510. [Google Scholar] [CrossRef]
- Zhang, D.; Hamauzu, Y. Phenolics, ascorbic acid, carotenoids and antioxidant activity of broccoli and their changes during conventional and microwave cooking. Food Chem. 2004, 88, 503–509. [Google Scholar] [CrossRef]
- Hwang, I.G.; Shin, Y.J.; Lee, S.; Lee, J.; Yoo, S.M. Effects of Different Cooking Methods on the Antioxidant Properties of Red Pepper (Capsicum annuum L.). Prev. Nutr. Food Sci. 2012, 17, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Russo, V.M.; Howard, L.R. Carotenoids in pungent and non-pungent peppers at various developmental stages grown in the field and glasshouse. J. Sci. Food Agric. 2002, 82, 615–624. [Google Scholar] [CrossRef]
- Hallmann, E.; Marszałek, K.; Lipowski, J.; Jasińska, U.; Kazimierczak, R.; Średnicka-Tober, D.; Rembiałkowska, E. Polyphenols and carotenoids in pickled bell pepper from organic and conventional production. Food Chem. 2019, 278, 254–260. [Google Scholar] [CrossRef]
- Hallmann, E.; Kazimierczak, R.; Marszałek, K.; Drela, N.; Kiernozek, E.; Toomik, P.; Matt, D.; Luik, A.; Rembiałkowska, E. The Nutritive Value of Organic and Conventional White Cabbage (Brassica OleraceaL. Var.Capitata) and Anti-Apoptotic Activity in Gastric Adenocarcinoma Cells of Sauerkraut Juice Produced Therof. J. Agric. Food Chem. 2017, 65, 8171–8183. [Google Scholar] [CrossRef] [PubMed]
- Al-Niaimi, F.; Chiang, N.Y.Z. Topical Vitamin C and the Skin: Mechanisms of Action and Clinical Applications. J. Clin. Aesthetic Dermatol. 2017, 10, 14–17. [Google Scholar]
- Sha, S.; Xu, D.; Wang, Y.; Zhao, W.; Li, X. Antihypertensive effects of fargesin in vitro and in vivo via attenuating oxidative stress and promoting nitric oxide release. Can. J. Physiol. Pharmacol. 2016, 94, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Ren, Y.-J.; Zhang, J.-J.; Luo, X.-P.; Yu, Z.-L.; Ren, G.-Y.; Sun, A.-N.; Deng, C.; Wang, Z.-T.; Dou, W. Anti-Inflammatory Effects of Fargesin on Chemically Induced Inflammatory Bowel Disease in Mice. Molecules 2018, 23, 1380. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.-H.; Kim, M.-S.; Le, M.-Q.; Song, Y.-S.; Bak, Y.; Ryu, H.-W.; Oh, S.-R.; Yoon, D.-Y. Fargesin exerts anti-inflammatory effects in THP-1 monocytes by suppressing PKC-dependent AP-1 and NF-ĸB signaling. Phytomedicine 2017, 24, 96–103. [Google Scholar] [CrossRef]
- Crews, C.; Chiodini, A.; Granvogl, M.; Hamlet, C.; Hrncirik, K.; Kuhlmann, J.; Lampen, A.; Scholz, G.; Weisshaar, R.; Wenzl, T.; et al. Analytical approaches for MCPD esters and glycidyl esters in food and biological samples: A review and future perspectives. Food Addit. Contam. Part A 2013, 30, 11–45. [Google Scholar] [CrossRef]
- Murakami, T.; Emoto, A.; Matsuda, H.; Yoshikawa, M. Medicinal Foodstuffs. XXI. Structures of New Cucurbitane-Type Triterpene Glycosides, Goyaglycosides-a, -b, -c, -d, -e, -f, -g, and -h, and New Oleanane-Type Triterpene Saponins, Goyasaponins I, II, and III, from the Fresh Fruit of Japanese Momordica charantia L. Chem. Pharm. Bull. 2001, 49, 54–63. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-Originating ACE Inhibitors, Including Antihypertensive Peptides, as Preventive Food Components in Blood Pressure Reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Guang, C.; Phillips, R.D. Plant Food-Derived Angiotensin I Converting Enzyme Inhibitory Peptides. J. Agric. Food Chem. 2009, 57, 5113–5120. [Google Scholar] [CrossRef]
- Campbell, W.C.; Fisher, M.H.; O Stapley, E.; Albers-Schonberg, G.; A Jacob, T. Ivermectin: A potent new antiparasitic agent. Science 1983, 221, 823–828. [Google Scholar] [CrossRef]
- Navrátilová, M.; Stuchlíková, L.R.; Moťková, K.; Szotáková, B.; Skálová, L.; Langhansová, L.; Podlipná, R. The Uptake of Ivermectin and Its Effects in Roots, Leaves and Seeds of Soybean (Glycine max). Molecules 2020, 25, 3655. [Google Scholar] [CrossRef]
- Reynolds, R.; Chappel, C. Sucrose Acetate Isobutyrate (SAIB): Historical Aspects of its use in Beverages and a Review of Toxicity Studies Prior to 1998. Food Chem. Toxicol. 1998, 36, 81–93. [Google Scholar] [CrossRef]
- Breithaupt, D.E.; Yahia, E.M.; Velázquez, F.J.V. Comparison of the absorption efficiency of α- and β-cryptoxanthin in female Wistar rats. Br. J. Nutr. 2007, 97, 329–336. [Google Scholar] [CrossRef][Green Version]
- De Azevedo-Meleiro, C.H.; Rodriguez-Amaya, D.B. Qualitative and quantitative differences in the carotenoid composition of yellow and red peppers determined by HPLC-DAD-MS. J. Sep. Sci. 2009, 32, 3652–3658. [Google Scholar] [CrossRef]
- Cruickshank-Quinn, C.; Quinn, K.D.; Powell, R.; Yang, Y.; Armstrong, M.; Mahaffey, S.; Reisdorph, R.; Reisdorph, N. Multi-step preparation technique to recover multiple metabolite compound classes for in-depth and informative metabolomic analysis. J. Vis. Exp. 2014. [Google Scholar] [CrossRef]
- Yang, Y.; Cruickshank, C.; Armstrong, M.; Mahaffey, S.; Reisdorph, R.; Reisdorph, N. New sample preparation approach for mass spectrometry-based profiling of plasma results in improved coverage of metabolome. J. Chromatogr. A 2013, 1300, 217–226. [Google Scholar] [CrossRef]
- O’Donnell, V.B.; Dennis, E.A.; Wakelam, M.J.O.; Subramaniam, S. LIPID MAPS: Serving the next generation of lipid researchers with tools, resources, data, and training. Sci. Signal. 2019, 12, eaaw2964. [Google Scholar] [CrossRef] [PubMed]
- Choquette, S.J.; Duewer, D.L.; Sharpless, K.E. NIST Reference Materials: Utility and Future. Annu. Rev. Anal. Chem. 2020, 13, 453–474. [Google Scholar] [CrossRef] [PubMed]
- Neveu, V.; Perez-Jimenez, J.; Vos, F.; Crespy, V.; Du Chaffaut, L.; Mennen, L.; Knox, C.; Eisner, R.; Cruz, J.; Wishart, D.; et al. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database 2010, 2010, bap024. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Stein, S.E. Estimating probabilities of correct identification from results of mass spectral library searches. J. Am. Soc. Mass Spectrom. 1994, 5, 316–323. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching molecular structure databases with tandem mass spectra using CSI:FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef] [PubMed]
- Sorokina, M.; Merseburger, P.; Rajan, K.; Yirik, M.A.; Steinbeck, C. COCONUT online: Collection of Open Natural Products database. J. Chemin. 2021, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; A Kapono, C.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Dreher, K. Putting the Plant Metabolic Network Pathway Databases to Work: Going Offline to Gain New Capabilities. Adv. Struct. Saf. Stud. 2014, 1083, 151–171. [Google Scholar] [CrossRef]
- Banerjee, P.; Erehman, J.; Gohlke, B.-O.; Wilhelm, T.; Preissner, R.; Dunkel, M. Super Natural II—A database of natural products. Nucleic Acids Res. 2015, 43, D935–D939. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compound Name | p-Value * | FDR | Green vs. Red ^ | Green vs. Yellow ^ | Red vs. Yellow ^ | Class |
|---|---|---|---|---|---|---|
| Beta-Cryptoxanthin | 0.000074 | 0.0080 | 0.010 | 0.677 | 0.035 | Carotenoid |
| PC(O-20:0/O-1:0) | 0.000128 | 0.0080 | 0.012 | 0.459 | 0.059 | Phospholipid/PC |
| PC(O-20:0/O-1:0) | 0.000129 | 0.0080 | 0.012 | 0.454 | 0.059 | Phospholipid/PC |
| PI(18:2/13:0) | 0.000154 | 0.0080 | 0.021 | 0.035 | 0.384 | Phospholipid/PI |
| DG(22:6/18:0) | 0.000769 | 0.0186 | 0.705 | 0.047 | 0.060 | Glycerolipid/DG |
| PG(22:2/20:0) | 0.001613 | 0.0318 | 0.052 | 0.117 | 0.999 | Phospholipid/PG |
| DG(20:5/14:0) | 0.002207 | 0.0366 | 0.053 | 0.071 | 0.761 | Glycerolipid/DG |
| TG(22:2/15:0/20:5) | 0.003347 | 0.0458 | 0.001 | 0.077 | 0.390 | Glycerolipid/TG |
| PI(P-20:0/14:1) | 0.003822 | 0.0458 | 0.014 | 0.002 | 0.331 | Phospholipid/PI |
| MGDG(18:1/18:1) | 0.003996 | 0.0458 | 0.999 | 0.095 | NA | Glycerolipid/MG |
| TG(18:4/20:5/18:4) | 0.004664 | 0.0458 | 0.018 | 0.002 | 0.240 | Glycerolipid/TG |
| Sucrose Acetate Isobutyrate (SAIB) | 0.006477 | 0.0560 | 0.008 | 0.387 | 0.276 | O-glycosyl |
| PI(22:1/16:0) | 0.008388 | 0.0611 | 0.017 | 0.030 | 0.957 | Phospholipid/PI |
| TG(22:4/20:2n6/22:5) | 0.008509 | 0.0611 | 0.097 | 0.303 | 0.796 | Glycerolipid/TG |
| PG(22:2/20:0) | 0.008535 | 0.0611 | 0.132 | 0.144 | 0.767 | Phospholipid/PG |
| DG(16:1/16:1) | 0.009819 | 0.0631 | 0.101 | 0.491 | NA | Glycerolipid/DG |
| DG(22:2/14:1) | 0.013877 | 0.0795 | 0.014 | 0.399 | 0.369 | Glycerolipid/DG |
| 2-ethenyl-2,4b,8,8-tetramethyl-tetradecahydrophenanthrene-3,5,10a-triol | 0.015343 | 0.0817 | 0.100 | 0.464 | 0.566 | Natural Product |
| Fargesin | 0.018186 | 0.0895 | 0.276 | 0.534 | 0.155 | Psoralen, from Fruits |
| Ascorbyl linoleate | 0.021860 | 0.1026 | 0.160 | 0.358 | 0.940 | Vitamin C metabolite |
| TG(22:4/15:0/22:4) | 0.025945 | 0.1075 | 0.255 | 0.211 | 0.710 | Glycerolipid/TG |
| TG(22:4/15:0/22:4) | 0.028741 | 0.1146 | 0.248 | 0.250 | 0.824 | Glycerolipid/TG |
| DG(18:4/15:0) | 0.030030 | 0.1154 | 0.136 | 0.717 | 0.462 | Glycerolipid/DG |
| PI(P-16:0/18:3) | 0.031557 | 0.1198 | 0.131 | 0.044 | 0.587 | Phospholipid/PI |
| Ascorbyl linoleate | 0.032964 | 0.1222 | 0.514 | 0.164 | 0.309 | Vitamin C metabolite |
| Ivermectin B1b | 0.033354 | 0.1222 | 0.110 | 0.049 | 0.676 | Antiparasitic drug |
| TG(20:4/15:0/22:5) | 0.035029 | 0.1268 | 0.912 | 0.046 | 0.062 | Glycerolipid/TG |
| Glycidyl oleate | 0.037508 | 0.1293 | 0.174 | 0.436 | 0.891 | Carboxylic ester |
| PE(15:0/20:0) | 0.038016 | 0.1293 | 0.162 | 0.575 | NA | Phospholipid/PE |
| PC(14:0/20:5) | 0.038145 | 0.1293 | 0.147 | 0.892 | 0.359 | Phospholipid/PC |
| Ascorbyl linoleate | 0.040104 | 0.1316 | 0.217 | 0.333 | 0.997 | Vitamin C metabolite |
| PC(15:0/18:4) | 0.041993 | 0.1364 | 0.245 | 0.294 | 0.926 | Phospholipid/PC |
| MGDG(18:2/18:3) | 0.043317 | 0.1392 | 0.398 | 0.220 | 0.553 | Glycerolipid/MG |
| Goyaglycoside g | 0.043762 | 0.1392 | 0.268 | 0.269 | 0.860 | Cucurbitacin glycosides |
| Ramipril | 0.045603 | 0.1408 | 0.991 | 0.253 | NA | Drug |
| PA(P-16:0/18:4) | 0.047472 | 0.1438 | 0.518 | 0.211 | 0.436 | Phospholipid/PA |
| all-trans-retinyl oleate | 0.048268 | 0.1443 | 0.388 | 0.466 | 0.052 | Prenol lipid |
| PC(14:1/20:5) | 0.050091 | 0.1448 | 0.196 | 0.590 | NA | Phospholipid/PC |
| Archaetidylglycerol-myo-inositol | 0.052834 | 0.1499 | 0.505 | 0.662 | 0.281 | Lipid |
| Color | Organic Status | Country of Origin | Season | Condition | Replicates |
|---|---|---|---|---|---|
| Green | Non-organic | Mexico | Fall 2018 | Raw | 3 |
| Green | Organic | The United States | Fall 2018 | Raw | 4 |
| Red | Organic | Mexico | Fall 2018 | Raw | 3 |
| Red | Non-organic | Canada | Fall 2018 | Raw | 3 |
| Yellow | Organic | Unknown | Spring 2019 | Raw/cooked | 2/3 |
| Red | Organic | Mexico | Spring 2019 | Raw/cooked | 2/3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutliff, A.K.; Saint-Cyr, M.; Hendricks, A.E.; Chen, S.S.; Doenges, K.A.; Quinn, K.; Westcott, J.; Tang, M.; Borengasser, S.J.; Reisdorph, R.M.; et al. Lipidomics-Based Comparison of Molecular Compositions of Green, Yellow, and Red Bell Peppers. Metabolites 2021, 11, 241. https://doi.org/10.3390/metabo11040241
Sutliff AK, Saint-Cyr M, Hendricks AE, Chen SS, Doenges KA, Quinn K, Westcott J, Tang M, Borengasser SJ, Reisdorph RM, et al. Lipidomics-Based Comparison of Molecular Compositions of Green, Yellow, and Red Bell Peppers. Metabolites. 2021; 11(4):241. https://doi.org/10.3390/metabo11040241
Chicago/Turabian StyleSutliff, Aimee K., Martine Saint-Cyr, Audrey E. Hendricks, Samuel S. Chen, Katrina A. Doenges, Kevin Quinn, Jamie Westcott, Minghua Tang, Sarah J. Borengasser, Richard M. Reisdorph, and et al. 2021. "Lipidomics-Based Comparison of Molecular Compositions of Green, Yellow, and Red Bell Peppers" Metabolites 11, no. 4: 241. https://doi.org/10.3390/metabo11040241
APA StyleSutliff, A. K., Saint-Cyr, M., Hendricks, A. E., Chen, S. S., Doenges, K. A., Quinn, K., Westcott, J., Tang, M., Borengasser, S. J., Reisdorph, R. M., Campbell, W. W., Krebs, N. F., & Reisdorph, N. A. (2021). Lipidomics-Based Comparison of Molecular Compositions of Green, Yellow, and Red Bell Peppers. Metabolites, 11(4), 241. https://doi.org/10.3390/metabo11040241