
Analysis of Host Jejunum Transcriptome and Associated Microbial Community Structure Variation in Young Calves with Feed-Induced Acidosis


Abstract
:1. Introduction
2. Results
2.1. Acidosis Model Induced by Feed
2.2. Transcriptome Changes in Host Jejunum Tissue
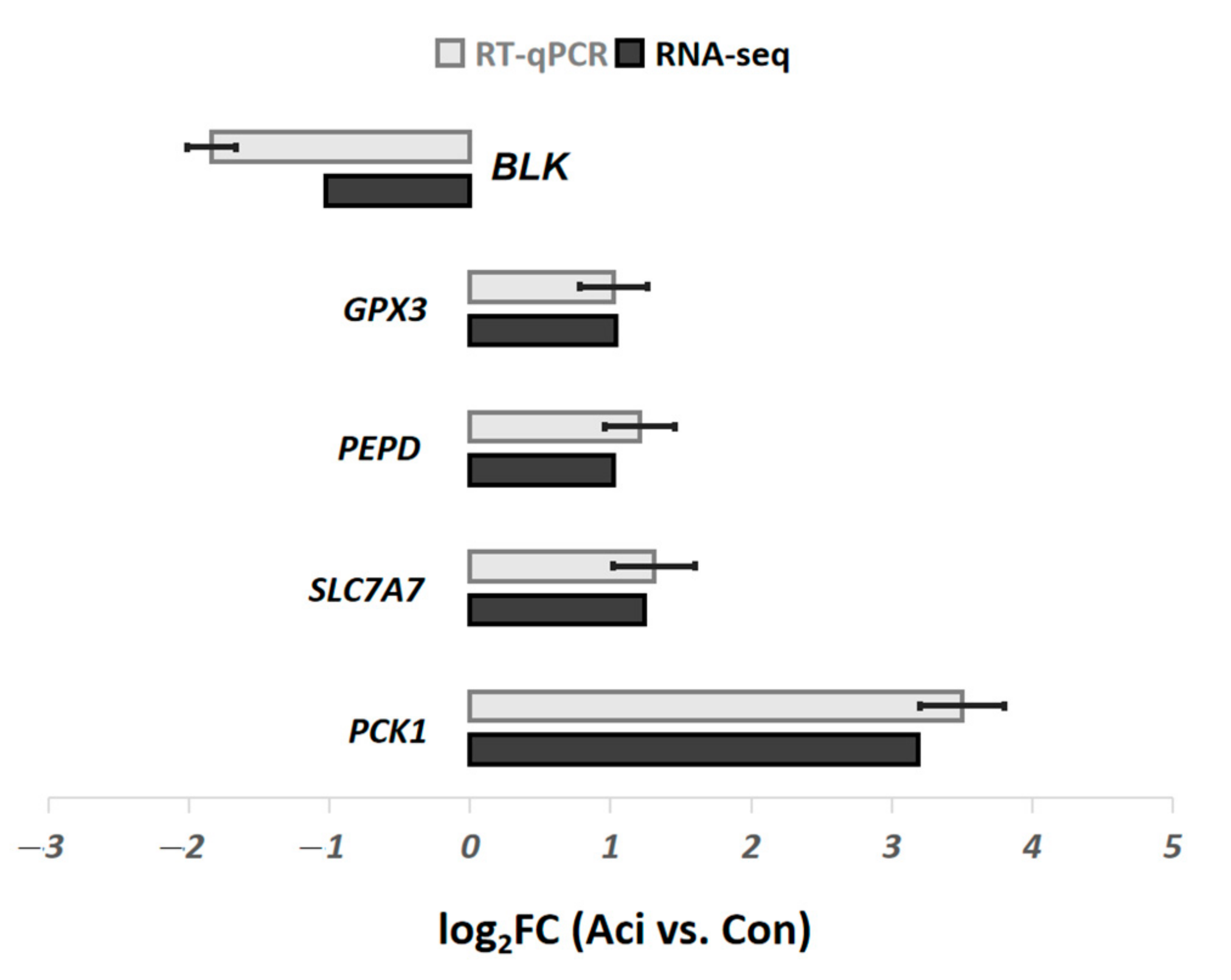
2.2.1. Differentially Expressed Genes in Jejunum
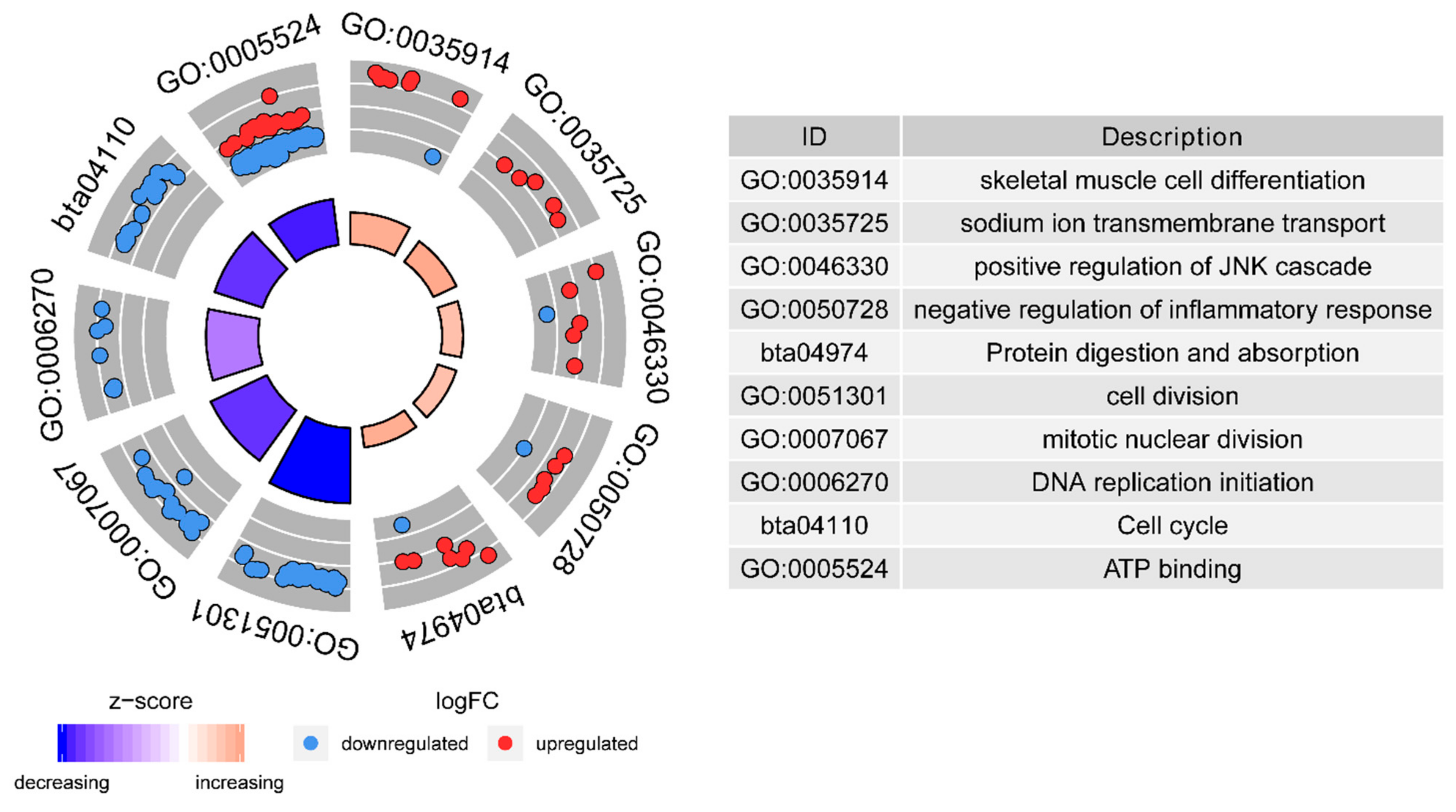
2.2.2. Functional Annotation of DEGs
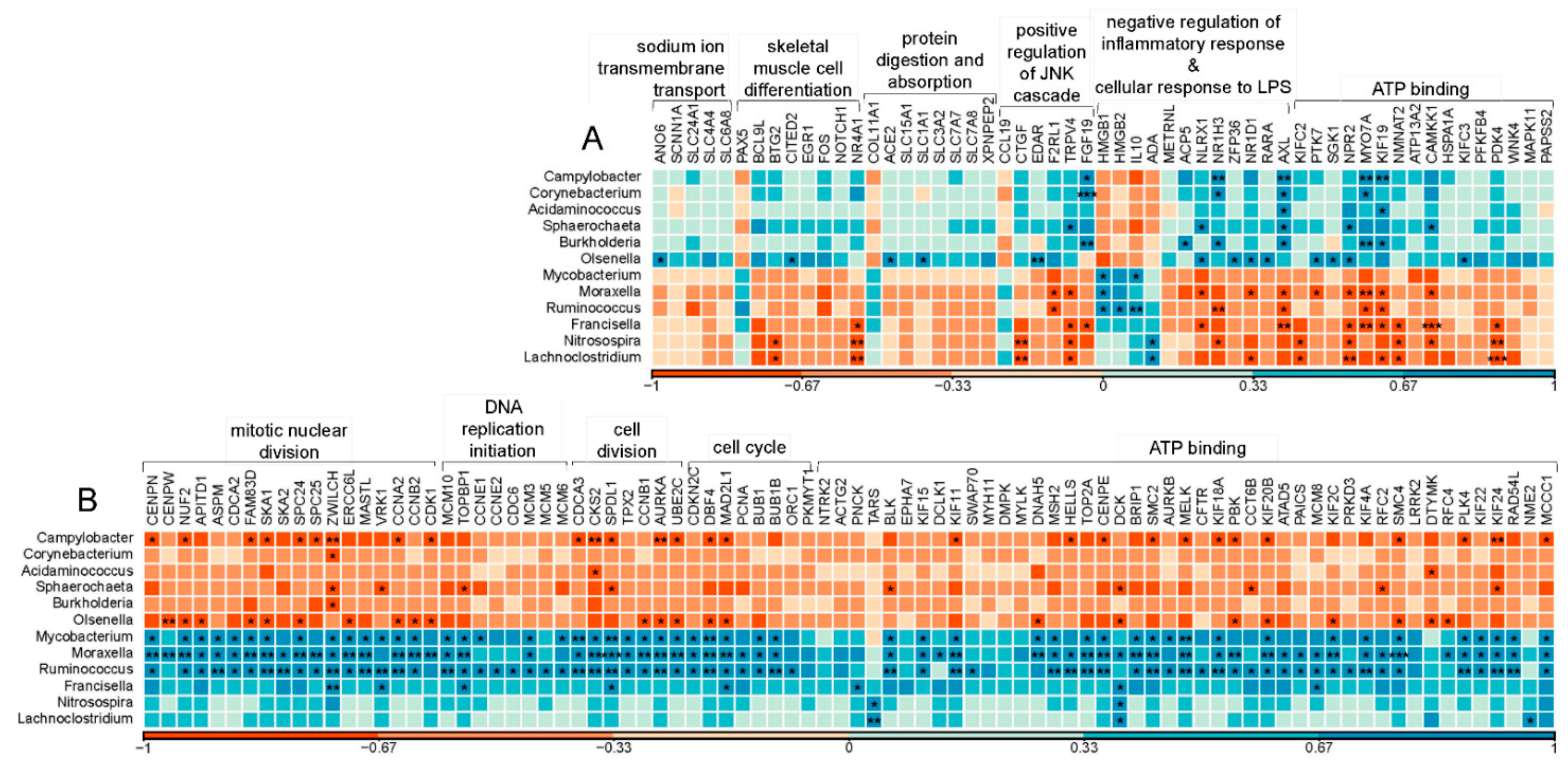
2.3. Jejunum Active Microorganisms and Its Association with Jejunum mRNA Expression Changes
3. Discussion
3.1. Jejunum Remodeling at the Transcriptome Level Caused by Acidosis-Induced Feed
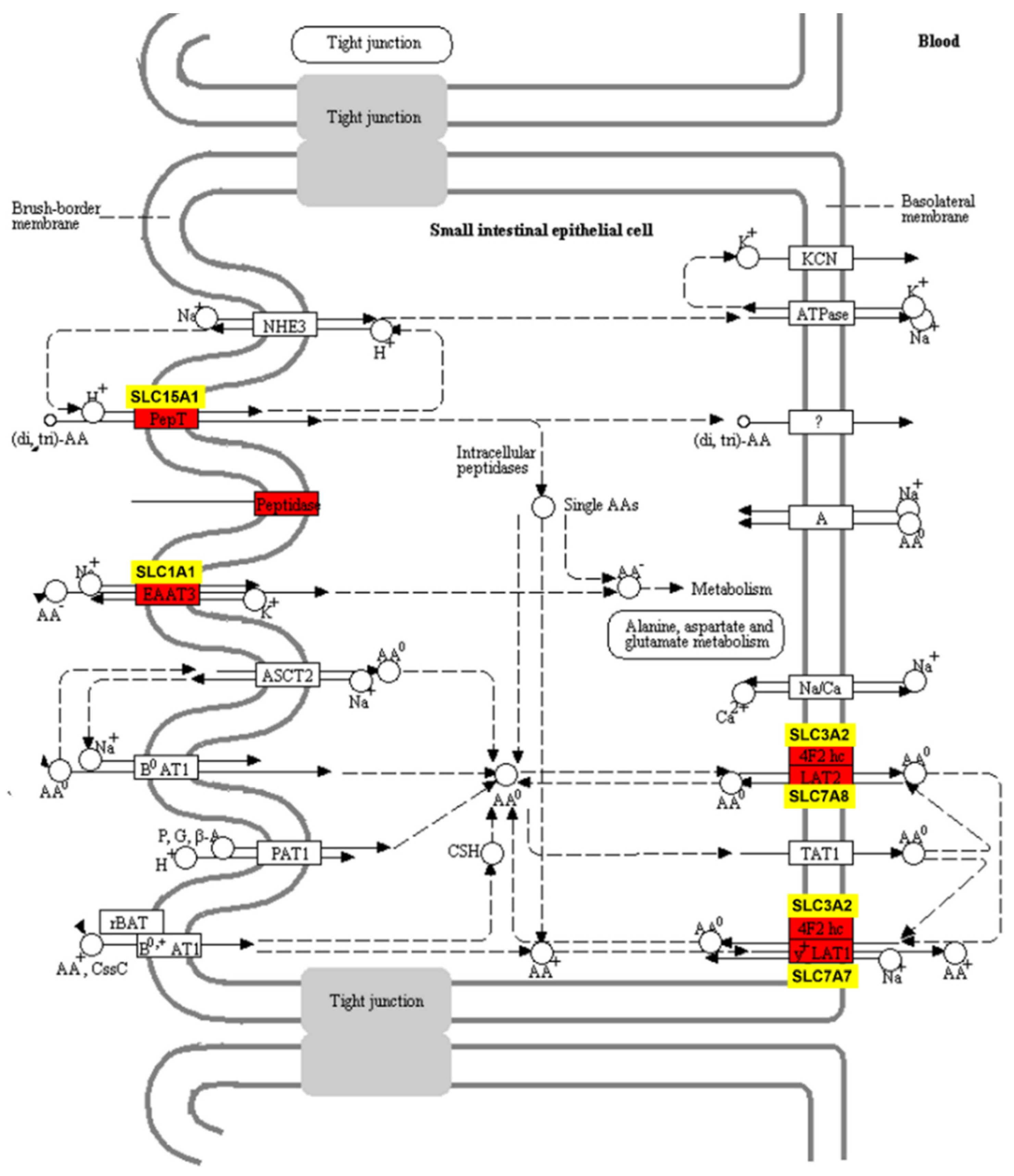
3.2. Significant Expression Changes in Nutrient Transporters and Sodium Ion Transmembrane Transport in the Jejunum of Acidotic Calves
3.3. Immune Response in Host Jejunum Tissue of Young Calves Treated with Acidosis-Inducing Diet
3.4. Active Jejunal Microbes and Its Association with Host mRNA Expression Changes
4. Materials and Methods
4.1. Acidosis Model Induced by Feed Remodeling
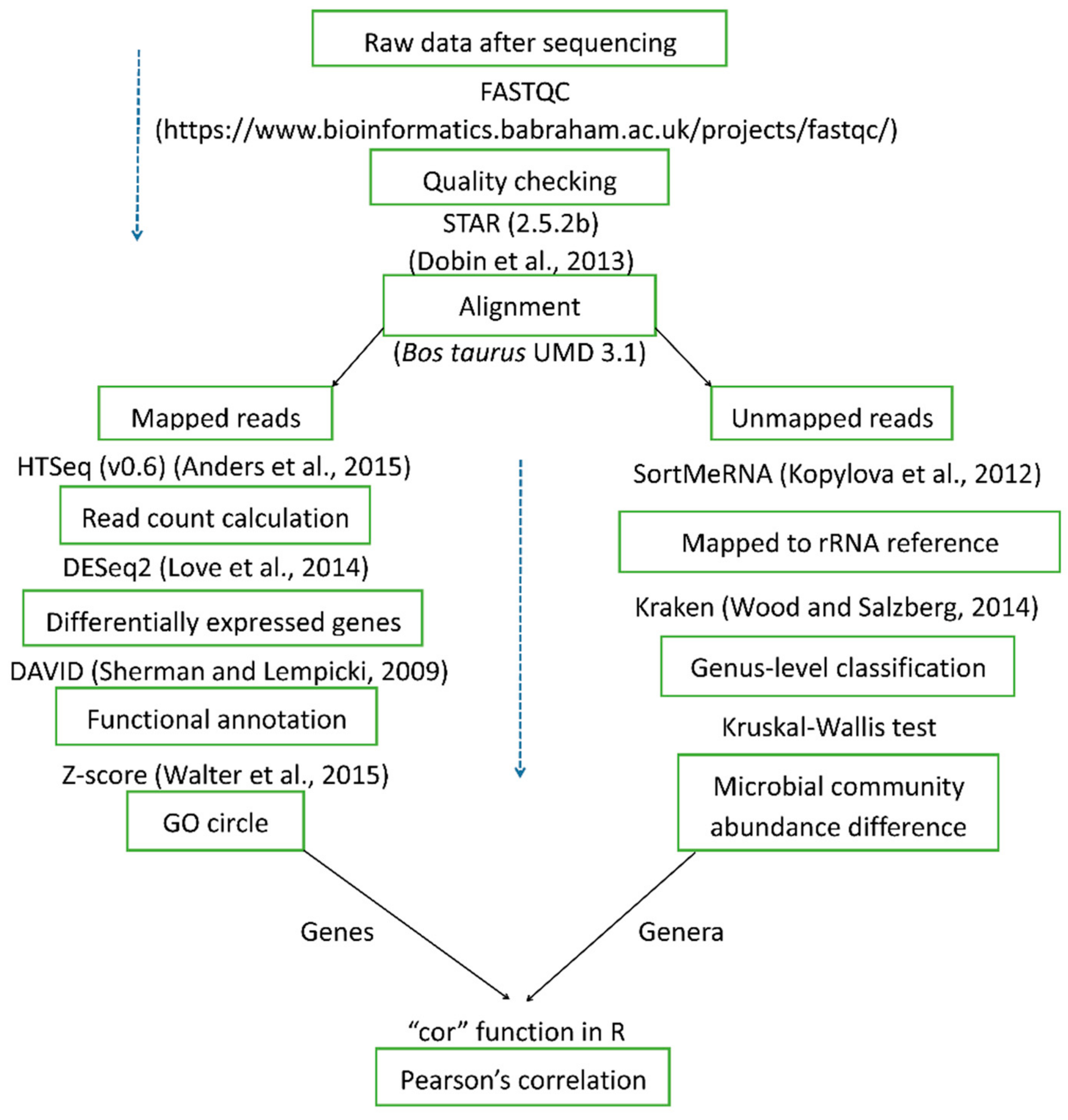
4.2. RNA Sequencing, Bioinformatics, and Statistical Analysis
4.3. RT-qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rey, M.; Enjalbert, F.; Combes, S.; Cauquil, L.; Bouchez, O.; Monteils, V. Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 2014, 116, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L.; Hurley, D.J.; Reber, A.J. Neonatal immune development in the calf and its impact on vaccine response. Vet. Clin. N. Am. Food A 2008, 24, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Pacha, J. Development of Intestinal Transport Function in Mammals. Physiol. Rev. 2000, 80, 1633–1667. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wang, Y.; Deng, Y.; Cao, Z.; Li, S.; Wang, J. Effects of feeding untreated, pasteurized and acidified waste milk and bunk tank milk on the performance, serum metabolic profiles, immunity, and intestinal development in Holstein calves. J. Anim. Sci. Biotechno. 2017, 8, 53. [Google Scholar] [CrossRef]
- Hammon, H.M.; Frieten, D.; Gerbert, C.; Koch, C.; Dusel, G.; Weikard, R.; Kuhn, C. Different milk diets have substantial effects on the jejunal mucosal immune system of pre-weaning calves, as demonstrated by whole transcriptome sequencing. Sci. Rep. 2018, 8, 1693. [Google Scholar] [CrossRef] [Green Version]
- Poudel, P.; Froehlich, K.; Casper, D.P.; St-Pierre, B. Feeding Essential Oils to Neonatal Holstein Dairy Calves Results in Increased Ruminal Prevotellaceae Abundance and Propionate Concentrations. Microorganisms 2019, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Aragona, K.M.; Suarez-Mena, F.X.; Dennis, T.S.; Quigley, J.D.; Hu, W.; Hill, T.M.; Schlotterbeck, R.L. Effect of starter form, starch concentration, and amount of forage fed on Holstein calf growth from 2 to 4 months of age. J. Dairy Sci. 2020, 103, 2324–2332. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Steele, M.A.; Croom, J.; Kahler, M.; Alzahal, O.; Mcbride, B.W. Bovine rumen epithelium undergoes rapid structural adaptations during grain-induced subacute ruminal acidosis. Am. J. Physiol. Reg. I 2011, 300, R1515–R1523. [Google Scholar] [CrossRef] [Green Version]
- Gozho, G.N.; Plaizier, J.C.; Krause, D.O.; Kennedy, A.D.; Wittenberg, K.M. Subacute Ruminal Acidosis Induces Ruminal Lipopolysaccharide Endotoxin Release and Triggers an Inflammatory Response—ScienceDirect. J. Dairy Sci. 2005, 88, 1399–1403. [Google Scholar] [CrossRef] [Green Version]
- Zebeli, Q.; Dijkstra, J.; Tafaj, M.; Steingass, H.; Ametaj, B.N.; Drochner, W. Modeling the Adequacy of Dietary Fiber in Dairy Cows Based on the Responses of Ruminal pH and Milk Fat Production to Composition of the Diet. J. Dairy Sci. 2008, 91, 2046–2066. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Cao, Y.; Liu, N.; Yang, X.; Yao, J.; Yan, D. Subacute ruminal acidosis challenge changed in situ degradability of feedstuffs in dairy goats. J. Dairy Sci. 2014, 97, 5101–5109. [Google Scholar] [CrossRef] [Green Version]
- Lean, I.J.; Golder, H.M. Ruminal Acidosis: Beyond pH and Rumen. In Proceedings of the 81st Cornell Nutrition Conference for Feed Manufacturers, East Syracuse, NY, USA, 22–24 October 2019; pp. 127–145. [Google Scholar]
- Stefanska, B.; Czapa, W.; Pruszynska-Oszmaek, E.; Szczepankiewicz, D.; Fievez, V.; Komisarek, J.; Stajek, K.; Nowak, W. Subacute ruminal acidosis affects fermentation and endotoxin concentration in the rumen and relative expression of the CD14/TLR4/MD2 genes involved in lipopolysaccharide systemic immune response in dairy cows. J. Dairy Sci. 2018, 101, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Plaizier, J.C.; Yoon, I.; Khafipour, E.S.; Scott, M. Impact of Saccharomyces cerevisiae fermentation product and subacute ruminal acidosis on production, inflammation, and fermentation in the rumen and hindgut of dairy cows. Anim. Feed. Sci. Tech. 2016, 211, 50–60. [Google Scholar] [CrossRef]
- Tappenden, K.A. Pathophysiology of short bowel syndrome: Considerations of resected and residual anatomy. JPEN-Parenter. Enter. 2014, 38, 14S–22S. [Google Scholar] [CrossRef]
- Norman, D.A.; Morawski, S.G.; Fordtran, J.S. Influence of glucose, fructose, and water movement on calcium absorption in the jejunum. Gastroenterology 1980, 78, 22–25. [Google Scholar] [CrossRef]
- Krejs, G.J.; Browne, R.; Raskin, P. Effect of intravenous somatostatin on jejunal absorption of glucose, amino acids, water, and electrolytes. Gastroenterology 1980, 78, 26. [Google Scholar] [CrossRef]
- El Nageh, M. Siege de l’absorption intestinale des gamma globulins du colostrum. Chez le veau nouveau-ne. Ann. Med. Vet. 1967, 11, 380. [Google Scholar]
- El Nageh, M. Periode de permeabilite de l’intestin du veau nouvean-ne aux gamma globulin du colostrum. Ann. Med. Vet. 1967, 11, 370. [Google Scholar]
- Qi, M.; Tan, B.E.; Wang, J.; Li, J.; Yin, Y.L. Small intestinal transcriptome analysis revealed changes of genes involved in nutrition metabolism and immune responses in growth retardation piglets. J. Anim. Sci. 2019, 97, 3795–3808. [Google Scholar] [CrossRef]
- Yan, Z.; Cai, L.; Huang, X.; Sun, W.; Li, S.; Wang, P.; Yang, Q.; Jiang, T.; Gun, S. Histological and Comparative Transcriptome Analyses Provide Insights into Small Intestine Health in Diarrheal Piglets after Infection with Clostridium Perfringens Type C. Animals 2019, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Bertocchi, M.; Sirri, F.; Palumbo, O.; Luise, D.; Trevisi, P. Exploring Differential Transcriptome between Jejunal and Cecal Tissue of Broiler Chickens. Animals 2019, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, S.; Huq, S.; Yatsunenko, T.; Haque, R.; Mahfuz, M.; Alam, M.A.; Benezra, A.; DeStefano, J.; Meier, M.F.; Muegge, B.D.; et al. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature 2014, 510, 417–421. [Google Scholar] [CrossRef]
- Koch, C.; Gerbert, C.; Frieten, D.; Dusel, G.; Hammon, H.M. Effects of ad libitum milk replacer feeding and butyrate supplementation on the epithelial growth and development of the gastrointestinal tract in Holstein calves. J. Dairy Sci. 2019, 102, 8513–8526. [Google Scholar] [CrossRef]
- Gelsinger, S.L.; Coblentz, W.K.; Zanton, G.I.; Ogden, R.K.; Akins, M.S. Physiological effects of starter-induced ruminal acidosis in calves before, during, and after weaning. J. Dairy Sci. 2020, 103, 2762–2772. [Google Scholar] [CrossRef]
- Li, W.; Gelsinger, S.; Edwards, A.; Riehle, C.; Koch, D. Transcriptome analysis of rumen epithelium and meta-transcriptome analysis of rumen epimural microbial community in young calves with feed induced acidosis. Sci. Rep. 2019, 9, 4744. [Google Scholar] [CrossRef]
- Li, W.; Gelsinger, S.; Edwards, A.; Riehle, C.; Koch, D. Changes in meta-transcriptome of rumen epimural microbial community and liver transcriptome in young calves with feed induced acidosis. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bi, R.; Liu, P. Sample size calculation while controlling false discovery rate for differential expression analysis with RNA-sequencing experiments. BMC Bioinform. 2016, 17, 146. [Google Scholar] [CrossRef] [Green Version]
- Morgan, D.O. The Cell Cycle: Principles of Control; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Li, Z.; Xu, X.Z. Post-Translational Modifications of the Mini-Chromosome Maintenance Proteins in DNA Replication. Genes 2019, 10, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsburg, S.L. Eukaryotic MCM proteins: Beyond replication initiation. Microbiol. Mol. Biol. Rev. 2004, 68, 109–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognetti, S.; Riera, A.; Speck, C. Switch on the engine: How the eukaryotic replicative helicase MCM2-7 becomes activated. Chromosoma 2015, 124, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, A.; Schwob, E.; Méndez, J. Excess MCM proteins protect human cells from replicative stress by licensing backup origins of replication. Proc. Natl. Acad. Sci. USA 2008, 105, 8956–8961. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.Q.; Jackson, D.A.; Blow, J.J. Dormant origins licensed by excess Mcm2–7 are required for human cells to survive replicative stress. Gene. Dev. 2007, 21, 3331–3341. [Google Scholar] [CrossRef] [Green Version]
- Musiałek, M.W.; Rybaczek, D. Behavior of replication origins in Eukaryota–spatio-temporal dynamics of licensing and firing. Cell Cycle 2015, 14, 2251–2264. [Google Scholar] [CrossRef]
- Ryu, S.; Holzschuh, J.; Erhardt, S.; Ettl, A.-K.; Driever, W. Depletion of minichromosome maintenance protein 5 in the zebrafish retina causes cell-cycle defect and apoptosis. Proc. Natl. Acad. Sci. USA 2005, 102, 18467–18472. [Google Scholar] [CrossRef] [Green Version]
- Shivji, M.K.; Kenny, M.K.; Wood, R.D. Proliferating cell nuclear antigen is required for DNA excision repair. Cell 1992, 69, 367–374. [Google Scholar] [CrossRef]
- Essers, J.; Theil, A.F.; Baldeyron, C.; van Cappellen, W.A.; Houtsmuller, A.B.; Kanaar, R.; Vermeulen, W. Nuclear dynamics of PCNA in DNA replication and repair. Mol. Cell. Biol. 2005, 25, 9350–9359. [Google Scholar] [CrossRef] [Green Version]
- de Greeff, A.; Resink, J.W.; van Hees, H.M.J.; Ruuls, L.; Klaassen, G.J.; Rouwers, S.M.G.; Stockhofe-Zurwieden, N. Supplementation of piglets with nutrient-dense complex milk replacer improves intestinal development and microbial fermentation. J. Anim. Sci. 2016, 94, 1012–1019. [Google Scholar] [CrossRef]
- Song, Z.H.; Tong, G.; Xiao, K.; Jiao, L.F.; Ke, Y.L.; Hu, C.H. L-Cysteine protects intestinal integrity, attenuates intestinal inflammation and oxidant stress, and modulates NF-B and Nrf2 pathways in weaned piglets after LPS challenge. Innate Immun. 2016, 22, 152–161. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Marzec, K.; Burgess, A. The Oncogenic Functions of MASTL Kinase. Front. Cell Dev. Biol. 2018, 6, 162. [Google Scholar] [CrossRef]
- Voets, E.; Wolthuis, R.M. MASTL is the human ortholog of Greatwall kinase that facilitates mitotic entry, anaphase and cytokinesis. Cell Cycle 2010, 9, 3591–3601. [Google Scholar] [CrossRef]
- Heim, A.; Konietzny, A.; Mayer, T.U. Protein phosphatase 1 is essential for Greatwall inactivation at mitotic exit. EMBO Rep. 2015, 16, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Vigneron, S.; Robert, P.; Strub, J.M.; Cianferani, S.; Castro, A.; Lorca, T. Greatwall dephosphorylation and inactivation upon mitotic exit is triggered by PP1. J. Cell Sci. 2016, 129, 1329–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, S.; Fey, D.; McCloy, R.A.; Parker, B.L.; Mitchell, N.J.; Payne, R.J.; Daly, R.J.; James, D.E.; Caldon, C.E.; Watkins, D.N. PP1 initiates the dephosphorylation of MASTL, triggering mitotic exit and bistability in human cells. J. Cell Sci. 2016, 129, 1340–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmena, M.; Earnshaw, W.C. The cellular geography of aurora kinases. Nat. Rev. Mol. Cell Biol. 2003, 4, 842–854. [Google Scholar] [CrossRef]
- Ding, J.; Swain, J.E.; Smith, G.D. Aurora kinase-A regulates microtubule organizing center (MTOC) localization, chromosome dynamics, and histone-H3 phosphorylation in mouse oocytes. Mol. Reprod. Dev. 2011, 78, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikonova, A.S.; Astsaturov, I.; Serebriiskii, I.G.; Dunbrack, R.L.; Golemis, E.A. Aurora A kinase (AURKA) in normal and pathological cell division. Cell. Mol. Life Sci. 2013, 70, 661–687. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.H.; Li, J.; Wu, B.Y. Gene expression profiling of the mouse gut: Effect of intestinal flora on intestinal health. Mol. Med. Rep. 2018, 17, 3667–3673. [Google Scholar] [CrossRef] [Green Version]
- Perland, E.; Fredriksson, R. Classification systems of secondary active transporters. Trends Pharmacol. Sci. 2017, 38, 305–315. [Google Scholar] [CrossRef]
- Broer, S. Amino acid transport across mammalian intestinal and renal epithelia. Physiol. Rev. 2008, 88, 249–286. [Google Scholar] [CrossRef]
- Woodward, A.D.; Holcombe, S.J.; Steibel, J.P.; Staniar, W.B.; Colvin, C.; Trottier, N.L. Cationic and neutral amino acid transporter transcript abundances are differentially expressed in the equine intestinal tract. J. Anim. Sci. 2010, 88, 1028–1033. [Google Scholar] [CrossRef] [Green Version]
- Rexhepaj, R.; Grahammer, F.; Volkl, H.; Remy, C.; Wagner, C.A.; Sandulache, D.; Artunc, F.; Henke, G.; Nammi, S.; Capasso, G.; et al. Reduced intestinal and renal amino acid transport in PDK1 hypomorphic mice. FASEB J. 2006, 20, 2214–2222. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.L.; Jiao, J.Z.; Zhou, C.S.; Tang, S.X.; Wang, M.; Kang, J.H.; Han, X.F.; Tan, Z.L. Effects of dietary methionine and lysine supplementation on nutrients digestion, serum parameters and mRNA expression of related aminoacid sensing and transporting genes in growing goats. Small Rumin. Res. 2018, 166, 1–6. [Google Scholar] [CrossRef]
- Pfeiffer, R.; Rossier, G.; Spindler, B.; Meier, C.; Kühn, L.; Verrey, F. Amino acid transport of y+ L-type by heterodimers of 4F2hc/CD98 and members of the glycoprotein-associated amino acid transporter family. EMBO J. 1999, 18, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Braun, D.; Wirth, E.K.; Wohlgemuth, F.; Reix, N.; Klein, M.O.; Gruters, A.; Kohrle, J.; Schweizer, U. Aminoaciduria, but normal thyroid hormone levels and signalling, in mice lacking the amino acid and thyroid hormone transporter Slc7a8. Biochem. J. 2011, 439, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Vanzant, E.; Harmon, D.; McLeod, K.; Boling, J.; Matthews, J. Ruminal and abomasal starch hydrolysate infusions selectively decrease the expression of cationic amino acid transporter mRNA by small intestinal epithelia of forage-fed beef steers. J. Dairy Sci. 2009, 92, 1124–1135. [Google Scholar] [CrossRef] [Green Version]
- Fei, Y.-J.; Kanai, Y.; Nussberger, S.; Ganapathy, V.; Leibach, F.H.; Romero, M.F.; Singh, S.K.; Boron, W.F.; Hediger, M.A. Expression cloning of a mammalian proton-coupled oligopeptide transporter. Nature 1994, 368, 563–566. [Google Scholar] [CrossRef]
- Shen, H.; Smith, D.E.; Brosius, F.C. Developmental expression of PEPT1 and PEPT2 in rat small intestine, colon, and kidney. Pediatr. Res. 2001, 49, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Luo, Y.Q.; Zhang, L.; Li, J.L.; Zhang, B.L.; Xing, S.; Zhu, Y.P.; Gao, F.; Zhou, G.H. Effects of cysteamine supplementation on the intestinal expression of amino acid and peptide transporters and intestinal health in finishing pigs. Anim. Sci. J. 2017, 88, 314–321. [Google Scholar] [CrossRef]
- Clarke, L.C.; Sweeney, T.; Curley, E.; Gath, V.; Duffy, S.K.; Vigors, S.; Rajauria, G.; O’Doherty, J.V. Effect of beta-glucanase and beta-xylanase enzyme supplemented barley diets on nutrient digestibility, growth performance and expression of intestinal nutrient transporter genes in finisher pigs. Anim. Feed. Sci. Tech. 2018, 238, 98–110. [Google Scholar] [CrossRef]
- Sora, I.; Richman, J.; Santoro, G.; Wei, H.; Wang, Y.; Vanderah, T.; Horvath, R.; Nguyen, M.; Waite, S.; Roeske, W.R. The cloning and expression of a human creatine transporter. Biochem. Bioph. Res. Co. 1994, 204, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Christie, D.L. Functional insights into the creatine transporter. In Creatine and Creatine Kinase in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2007; pp. 99–118. [Google Scholar]
- Takenaka, M.; Bagnasco, S.M.; Preston, A.S.; Uchida, S.; Yamauchi, A.; Kwon, H.M.; Handler, J.S. The canine betaine gamma-amino-n-butyric acid transporter gene: Diverse mRNA isoforms are regulated by hypertonicity and are expressed in a tissue-specific manner. Proc. Natl. Acad. Sci. USA 1995, 92, 1072–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braissant, O.; Henry, H. AGAT, GAMT and SLC6A8 distribution in the central nervous system, in relation to creatine deficiency syndromes: A review. J. Inherit. Metab. Dis. 2008, 31, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, C.; Waldvogel, H.; Dodd, J.; Gilbert, R.; Lowe, M.; Birch, N.; Faull, R.; Christie, D. Immunohistochemical localisation of the creatine transporter in the rat brain. Neuroscience 2009, 163, 571–585. [Google Scholar] [CrossRef]
- Taylor, J.; Mager, E.; Grosell, M. Basolateral NBCe1 plays a rate-limiting role in transepithelial intestinal HCO3–secretion, contributing to marine fish osmoregulation. J. Exp. Biol. 2010, 213, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.; Gregorio, S.F.; Egger, R.C.; Fuentes, J. Molecular and functional regionalization of bicarbonate secretion cascade in the intestine of the European sea bass (Dicentrarchus labrax). Comp. Biochem. Phys. A 2019, 233, 53–64. [Google Scholar] [CrossRef]
- Cui, Z.; Wu, S.; Li, J.; Yang, Q.E.; Chai, S.; Wang, L.; Wang, X.; Zhang, X.; Liu, S.; Yao, J. Effect of Alfalfa Hay and Starter Feeding Intervention on Gastrointestinal Microbial Community, Growth and Immune Performance of Yak Calves. Front. Microbiol. 2020, 11, 994. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Li, M.; Goonewardene, L.A.; Oba, M.; Guan, L.L. Effect of calf starter feeding on gut microbial diversity and expression of genes involved in host immune responses and tight junctions in dairy calves during weaning transition. J. Dairy Sci. 2013, 96, 3189–3200. [Google Scholar] [CrossRef]
- Philipson, C.W.; Bassaganya-Riera, J.; Viladomiu, M.; Kronsteiner, B.; Abedi, V.; Hoops, S.; Michalak, P.; Kang, L.; Girardin, S.E.; Hontecillas, R. Modeling the regulatory mechanisms by which NLRX1 modulates innate immune responses to Helicobacter pylori infection. PLoS ONE 2015, 10, e0137839. [Google Scholar] [CrossRef] [Green Version]
- Castano-Rodriguez, N.; Kaakoush, N.O.; Goh, K.-L.; Fock, K.M.; Mitchell, H.M. The NOD-like receptor signalling pathway in Helicobacter pylori infection and related gastric cancer: A case-control study and gene expression analyses. PLoS ONE 2014, 9, e0117870. [Google Scholar] [CrossRef]
- Miyata, K.S.; McCaw, S.E.; Patel, H.V.; Rachubinski, R.A.; Capone, J.P. The orphan nuclear hormone receptor LXR interacts with the peroxisome proliferator-activated receptor and inhibits peroxisome proliferator signaling. J. Biol. Chem. 1996, 271, 9189–9192. [Google Scholar] [CrossRef] [Green Version]
- McFadden, J.; Corl, B. Activation of liver X receptor (LXR) enhances de novo fatty acid synthesis in bovine mammary epithelial cells. J. Dairy Sci. 2010, 93, 4651–4658. [Google Scholar] [CrossRef]
- Meertens, L.; Labeau, A.; Dejarnac, O.; Cipriani, S.; Sinigaglia, L.; Bonnet-Madin, L.; Le Charpentier, T.; Hafirassou, M.L.; Zamborlini, A.; Cao-Lormeau, V.-M. Axl mediates ZIKA virus entry in human glial cells and modulates innate immune responses. Cell Rep. 2017, 18, 324–333. [Google Scholar] [CrossRef]
- Rothlin, C.V.; Ghosh, S.; Zuniga, E.I.; Oldstone, M.B.; Lemke, G. TAM receptors are pleiotropic inhibitors of the innate immune response. Cell 2007, 131, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Petri, R.; Schwaiger, T.; Penner, G.; Beauchemin, K.; Forster, R.; McKinnon, J.; McAllister, T. Changes in the rumen epimural bacterial diversity of beef cattle as affected by diet and induced ruminal acidosis. Appl. Environ. Microbiol. 2013, 79, 3744–3755. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, G.T. Microbial community composition along the digestive tract in forage- and grain-fed bison. BMC Vet. Res. 2017, 13, 253. [Google Scholar] [CrossRef] [Green Version]
- Wetzels, S.U.; Mann, E.; Pourazad, P.; Qumar, M.; Pinior, B.; Metzler-Zebeli, B.U.; Wagner, M.; Schmitz-Esser, S.; Zebeli, Q. Epimural bacterial community structure in the rumen of Holstein cows with different responses to a long-term subacute ruminal acidosis diet challenge. J. Dairy Sci. 2017, 100, 1829–1844. [Google Scholar] [CrossRef]
- Kim, Y.H.; Nagata, R.; Ohkubo, A.; Ohtani, N.; Kushibiki, S.; Ichijo, T.; Sato, S. Changes in ruminal and reticular pH and bacterial communities in Holstein cattle fed a high-grain diet. BMC Vet. Res. 2018, 14, 310. [Google Scholar] [CrossRef]
- Petri, R.M.; Kleefisch, M.T.; Metzler-Zebeli, B.U.; Zebeli, Q.; Klevenhusen, F. Changes in the Rumen Epithelial Microbiota of Cattle and Host Gene Expression in Response to Alterations in Dietary Carbohydrate Composition. Appl. Environ. Microbiol. 2018, 84, AEM.00384-18. [Google Scholar] [CrossRef] [Green Version]
- Vorwerk, H.; Huber, C.; Mohr, J.; Bunk, B.; Bhuju, S.; Wensel, O.; Spröer, C.; Fruth, A.; Flieger, A.; Schmidt-Hohagen, K. A transferable plasticity region in C ampylobacter coli allows isolates of an otherwise non-glycolytic food-borne pathogen to catabolize glucose. Mol. Microbiol. 2015, 98, 809–830. [Google Scholar] [CrossRef] [Green Version]
- Vegge, C.S.; Jansen van Rensburg, M.J.; Rasmussen, J.J.; Maiden, M.C.; Johnsen, L.G.; Danielsen, M.; MacIntyre, S.; Ingmer, H.; Kelly, D.J. Glucose metabolism via the Entner-Doudoroff pathway in Campylobacter: A rare trait that enhances survival and promotes biofilm formation in some isolates. Front. Microbiol. 2016, 7, 1877. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.J. Complexity and Versatility in the Physiology and Metabolism of Campylobacter jejuni. In Campylobacter, 3rd ed.; Amer Soc Microbiology: Washington, DC, USA, 2008; pp. 41–61. [Google Scholar]
- Moore, J.E.; Corcoran, D.; Dooley, J.S.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Megraud, F.; Millar, B.C.; O’Mahony, R.; et al. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef] [Green Version]
- Sauerwein, R.; Horrevorts, A.; Bisseling, J. Septic abortion associated withCampylobacter fetus subspeciesfetus infection: Case report and review of the literature. Infection 1993, 21, 331–333. [Google Scholar] [CrossRef]
- Kraatz, M.; Wallace, R.J.; Svensson, L. Olsenella umbonata sp. nov., a microaerotolerant anaerobic lactic acid bacterium from the sheep rumen and pig jejunum, and emended descriptions of Olsenella, Olsenella uli and Olsenella profusa. Int. J. Syst. Evol. Micr. 2011, 61, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Malmuthuge, N.; Griebel, P.J. Taxonomic identification of commensal bacteria associated with the mucosa and digesta throughout the gastrointestinal tracts of preweaned calves. Appl. Environ. Microbiol. 2014, 80, 2021–2028. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Z.; Liu, K.Z.; Wang, Z.S.; Bai, X.; Peng, Q.H.; Jin, L. Bacterial Community Diversity Associated with Different Utilization Efficiencies of Nitrogen in the Gastrointestinal Tract of Goats. Front. Microbiol. 2019, 10, 239. [Google Scholar] [CrossRef]
- Gyuraszova, M.; Kovalcikova, A.; Gardlik, R. Association between oxidative status and the composition of intestinal microbiota along the gastrointestinal tract. Med. Hypotheses 2017, 103, 81–85. [Google Scholar] [CrossRef]
- Suen, G.; Stevenson, D.M.; Bruce, D.C.; Chertkov, O.; Copeland, A.; Cheng, J.-F.; Detter, C.; Detter, J.C.; Goodwin, L.A.; Han, C.S. Complete genome of the cellulolytic ruminal bacterium Ruminococcus albus 7. Am. Soc. Microbiol. 2011, 193, 5574–5575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Liu, F.; Cai, W.; Jia, C.; Song, J. Diet-induced changes in bacterial communities in the jejunum and their associations with bile acids in Angus beef cattle. Anim. Microbiome 2020, 2, 33. [Google Scholar] [CrossRef]
- Vangylswyk, N.O. Succiniclasticum Ruminis Gen-Nov, Sp-Nov, A Ruminal Bacterium Converting Succinate to Propionate as the Sole Energy-Yielding Mechanism. Int. J. Syst. Bacteriol. 1995, 45, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Kim, J.; Kim, J.; Kim, I. Amino acid digestibility of single cell protein from Corynebacterium ammoniagenes in growing pigs. Anim. Feed. Sci. Tech. 2013, 180, 111–114. [Google Scholar] [CrossRef]
- Cook, G.M.; Wells, J.E.; Russell, J.B. Ability of acidaminococcus-fermentans to oxidize trans-aconitate and decrease the accumulation of tricarballylate, a toxic end-product of ruminal fermentation. Appl. Environ. Microbiol. 1994, 60, 2533–2537. [Google Scholar] [CrossRef] [Green Version]
- Myer, P.R.; Wells, J.E.; Smith, T.P.L.; Kuehn, L.A.; Freetly, H.C. Microbial community profiles of the jejunum from steers differing in feed efficiency. J. Anim. Sci. 2016, 94, 327–338. [Google Scholar] [CrossRef]
- Wang, J.; Han, Y.; Zhao, J.Z.; Zhou, Z.J.; Fan, H. Pyrosequencing-based analysis of the complex microbiota located in the gastrointestinal tracts of growing-finishing pigs. Anim Prod. Sci. 2019, 59, 870–878. [Google Scholar] [CrossRef]
- Zhu, Y.Z.; Shi, C.; Niu, Q.Y.; Wang, J.; Zhu, W.Y. Dynamic changes in morphology, gene expression and microbiome in the jejunum of compensatory-growth rats induced by protein restriction. Microb. Biotechnol. 2018, 11, 734–746. [Google Scholar] [CrossRef]
- Lu, J.; Idris, U.; Harmon, B.; Hofacre, C.; Maurer, J.J.; Lee, M.D. Diversity and succession of the intestinal bacterial community of the maturing broiler chicken. Appl. Environ. Microbiol. 2003, 69, 6816–6824. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.P.; Xiang, Y.; Zhou, W.D.; Chen, J.G.; Li, K.F.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef]
- Balows, A.; Trüper, H.G.; Dworkin, M.; Harder, W.; Schleifer, K.H. The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Springer: Heidelberg, Germany, 1992. [Google Scholar]
- Kristensen, N.B.; Engbaek, M.; Vestergaard, M.; Harmon, D.L. Technical note: Ruminal cannulation technique in young Holstein calves: Effects of cannulation on feed intake, body weight gain, and ruminal development at six weeks of age. J. Dairy Sci. 2010, 93, 737–742. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, B.T. Extracting biological meaning from large gene lists with DAVID. In Chapter 13 in Current Protocols in Bioinformatics; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009. [Google Scholar]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Burge, S.W.; Daub, J.; Eberhardt, R.; Tate, J.; Barquist, L.; Nawrocki, E.P.; Eddy, S.R.; Gardner, P.P.; Bateman, A. Rfam 11.0: 10 years of RNA families. Nucleic Acids Res. 2013, 41, D226–D232. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [Green Version]
- Tuo, L.; Xiang, J.; Pan, X.; Hu, J.; Tang, H.; Liang, L.; Xia, J.; Hu, Y.; Zhang, W.; Huang, A.; et al. PCK1 negatively regulates cell cycle progression and hepatoma cell proliferation via the AMPK/p27(Kip1) axis. J. Exp. Clin. Cancer Res. 2019, 38, 50. [Google Scholar] [CrossRef] [Green Version]
- Sperandeo, P.; Lau, F.K.; Carpentieri, A.; De Castro, C.; Molinaro, A.; Deho, G.; Silhavy, T.J.; Polissi, A. Functional analysis of the protein machinery required for transport of lipopolysaccharide to the outer membrane of Escherichia coli. J. Bacteriol. 2008, 190, 4460–4469. [Google Scholar] [CrossRef] [Green Version]
- Mitsubuchi, H.; Nakamura, K.; Matsumoto, S.; Endo, F. Inborn errors of proline metabolism. J. Nutr. 2008, 138, 2016S–2020S. [Google Scholar] [CrossRef]
- Takebe, G.; Yarimizu, J.; Saito, Y.; Hayashi, T.; Nakamura, H.; Yodoi, J.; Nagasawa, S.; Takahashi, K. A comparative study on the hydroperoxide and thiol specificity of the glutathione peroxidase family and selenoprotein P. J. Biol. Chem. 2002, 277, 41254–41258. [Google Scholar] [CrossRef] [Green Version]
- Compeer, E.B.; Janssen, W.; van Royen-Kerkhof, A.; van Gijn, M.; van Montfrans, J.M.; Boes, M. Dysfunctional BLK in common variable immunodeficiency perturbs B-cell proliferation and ability to elicit antigen-specific CD4+ T-cell help. Oncotarget 2015, 6, 10759–10771. [Google Scholar] [CrossRef]
- Borowiec, M.; Liew, C.W.; Thompson, R.; Boonyasrisawat, W.; Hu, J.; Mlynarski, W.M.; El Khattabi, I.; Kim, S.H.; Marselli, L.; Rich, S.S.; et al. Mutations at the BLK locus linked to maturity onset diabetes of the young and beta-cell dysfunction. Proc. Natl. Acad. Sci. USA 2009, 106, 14460–14465. [Google Scholar] [CrossRef] [Green Version]
- Arseneau, J.R.; Laflamme, M.; Lewis, S.M.; Maicas, E.; Ouellette, R.J. Multiple isoforms of PAX5 are expressed in both lymphomas and normal B-cells. Brit. J. Haematol. 2009, 147, 328–338. [Google Scholar] [CrossRef]
- Schormann, N.; Ricciardi, R.; Chattopadhyay, D. Uracil-DNA glycosylases—structural and functional perspectives on an essential family of DNA repair enzymes. Protein Sci. 2014, 23, 1667–1685. [Google Scholar] [CrossRef] [Green Version]
- Wisnieski, F.; Calcagno, D.Q.; Leal, M.F.; dos Santos, L.C.; Gigek, C.D.; Chen, E.S.; Pontes, T.B.; Assumpcao, P.P.; de Assumpcao, M.B.; Demachki, S.; et al. Reference genes for quantitative RT-PCR data in gastric tissues and cell lines. World J. Gastroentero. 2013, 19, 7121–7128. [Google Scholar] [CrossRef]
- Die, J.V.; Baldwin, R.L.; Rowland, L.J.; Li, R.; Oh, S.; Li, C.J.; Connor, E.E.; Ranilla, M.J. Selection of internal reference genes for normalization of reverse transcription quantitative polymerase chain reaction (RT-qPCR) analysis in the rumen epithelium. PLoS ONE 2017, 12, e0172674. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Normalized Read Counts (Mean ± SEM) | p-Value | Description | |
|---|---|---|---|---|
| Con | Aci | |||
| Campylobacter | 738.22 ± 208.78 | 3581.05 ± 1312.38 | 0.021 | Gram-negative |
| Corynebacterium | 1096.51 ± 181.40 | 2828.44 ± 1278.40 | 0.021 | Gram-positive |
| Acidaminococcus | 658.54 ± 88.52 | 2119.04 ± 1371.72 | 0.021 | Gram-negative |
| Olsenella | 484.36 ± 174.05 | 1521.03 ± 908.39 | 0.043 | Gram-positive |
| Sphaerochaeta | 206.85 ± 44.44 | 520.41 ± 172.81 | 0.043 | -- |
| Burkholderia | 0.00 | 129.69 ± 116.15 | 0.047 | Gram-negative |
| Moraxella | 370.53 ± 64.52 | 60.63 ± 37.34 | 0.020 | Gram-negative |
| Francisella | 3510.84 ± 200.53 | 1768.27 ± 361.60 | 0.021 | Gram-negative |
| Nitrosospira | 4510.05 ± 690.55 | 1783.64 ± 546.93 | 0.021 | -- |
| Lachnoclostridium | 5588.09 ± 1076.92 | 1553.04 ± 356.10 | 0.021 | Butyrate producer |
| Mycobacterium | 176.47 ± 55.11 | 22.34 ± 22.34 | 0.038 | Cannot be stained |
| Ruminococcus | 1847.28 ± 380.77 | 546.59 ± 212.98 | 0.043 | Gram-positive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaowa, N.; Li, W.; Gelsinger, S.; Murphy, B.; Li, S. Analysis of Host Jejunum Transcriptome and Associated Microbial Community Structure Variation in Young Calves with Feed-Induced Acidosis. Metabolites 2021, 11, 414. https://doi.org/10.3390/metabo11070414
Gaowa N, Li W, Gelsinger S, Murphy B, Li S. Analysis of Host Jejunum Transcriptome and Associated Microbial Community Structure Variation in Young Calves with Feed-Induced Acidosis. Metabolites. 2021; 11(7):414. https://doi.org/10.3390/metabo11070414
Chicago/Turabian StyleGaowa, Naren, Wenli Li, Sonia Gelsinger, Brianna Murphy, and Shengli Li. 2021. "Analysis of Host Jejunum Transcriptome and Associated Microbial Community Structure Variation in Young Calves with Feed-Induced Acidosis" Metabolites 11, no. 7: 414. https://doi.org/10.3390/metabo11070414
APA StyleGaowa, N., Li, W., Gelsinger, S., Murphy, B., & Li, S. (2021). Analysis of Host Jejunum Transcriptome and Associated Microbial Community Structure Variation in Young Calves with Feed-Induced Acidosis. Metabolites, 11(7), 414. https://doi.org/10.3390/metabo11070414