
Azoxymethane Alters the Plasma Metabolome to a Greater Extent in Mice Fed a High-Fat Diet Compared to an AIN-93 Diet


Abstract
1. Introduction
2. Results
2.1. Identified Metabolites and Their Group Separation
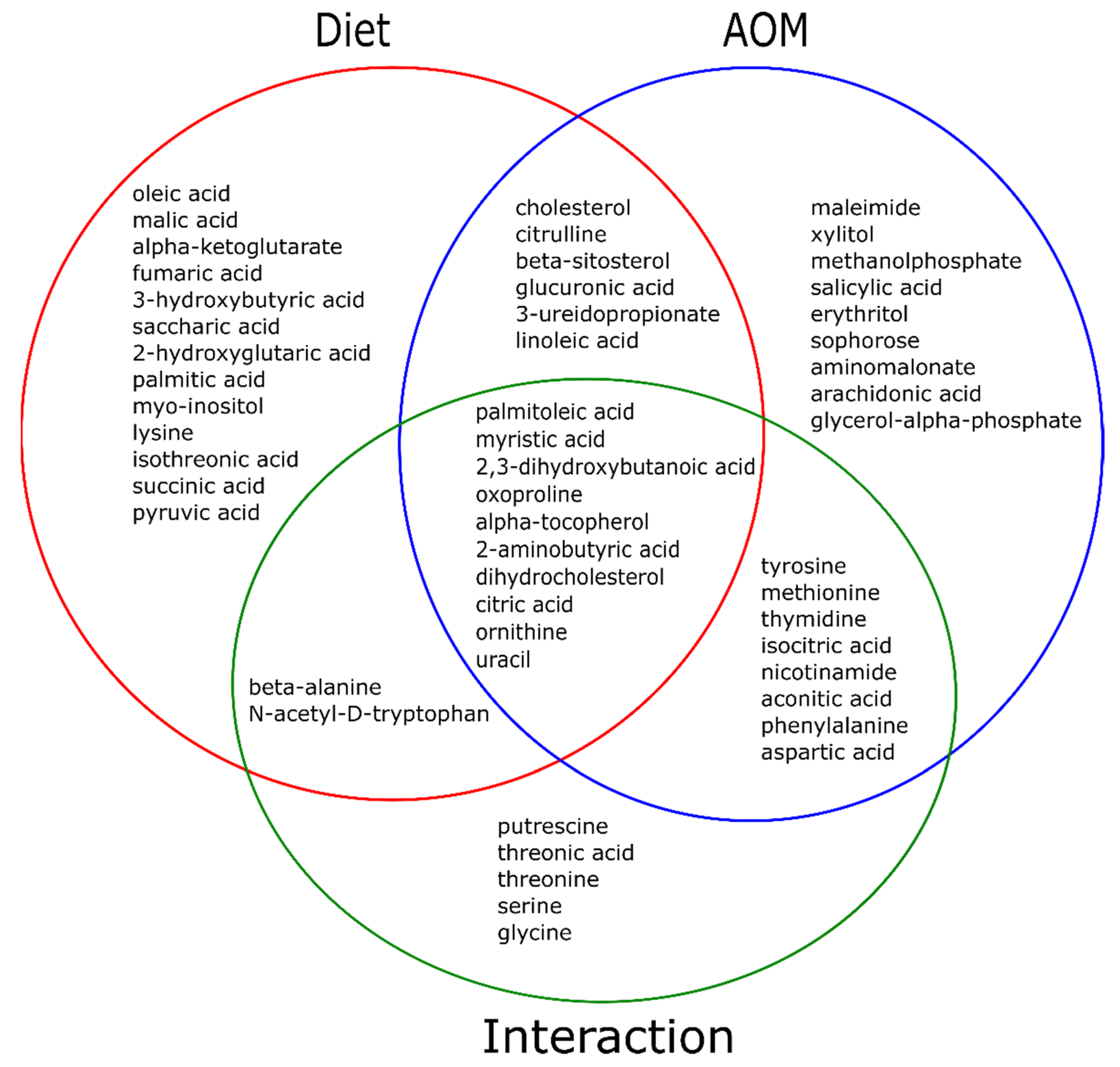
2.2. Diet and AOM Interaction
2.3. Metabolic Pathways of Altered Metabolites
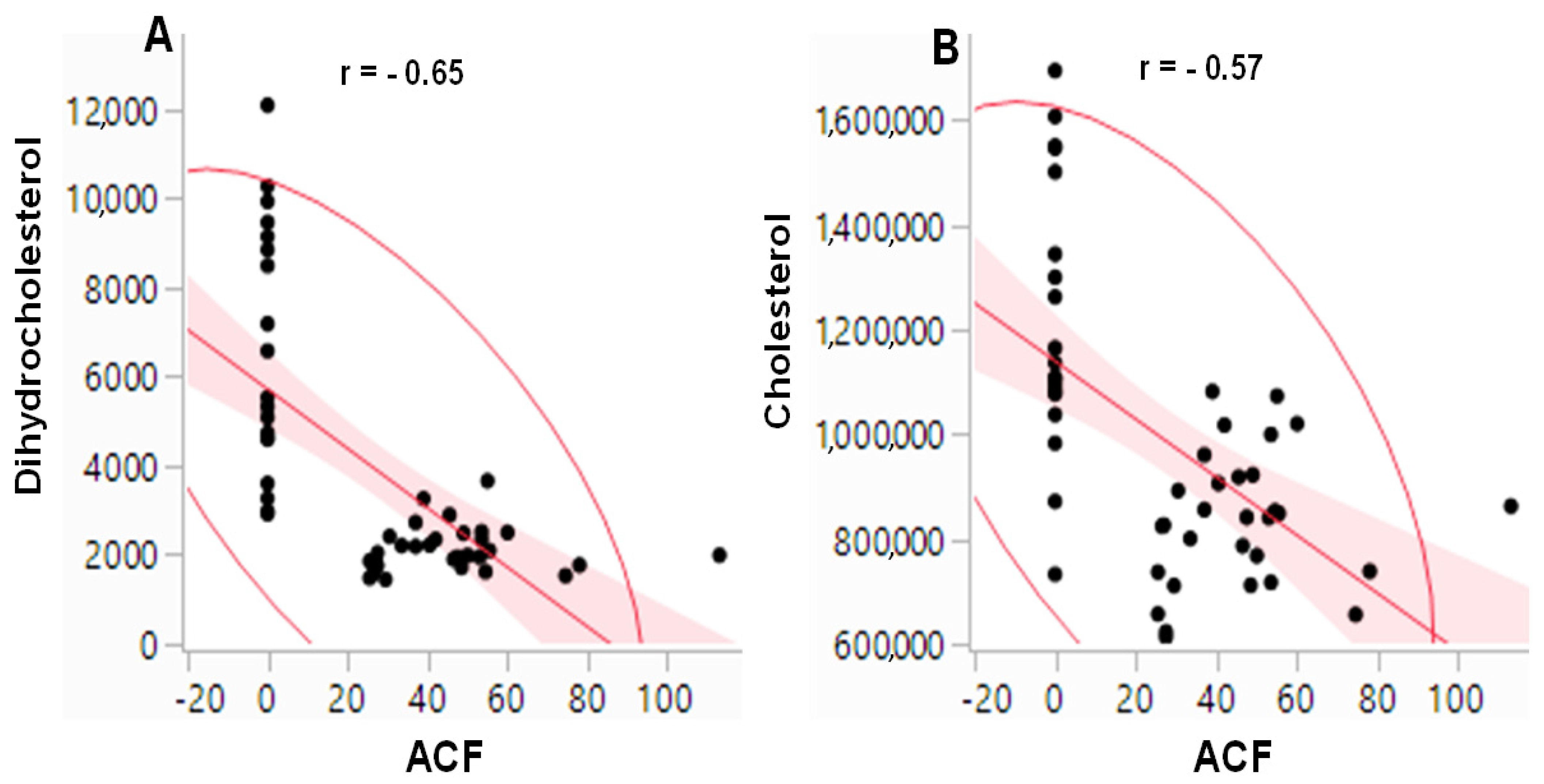
2.4. Correlation between Colonic ACF and Dihydrocholesterol/Cholesterol
3. Discussion
4. Materials and Methods
4.1. Animals, Diets, and AOM Treatment
4.2. Plasma Metabolomics
4.3. Statistical and Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Zeng, H.; Lazarova, D.L. Obesity-related colon cancer: Dietary factors and their mechanisms of anticancer action. Clin. Exp. Pharmacol. Physiol. 2012, 39, 161–167. [Google Scholar] [CrossRef]
- Roslan, N.H.; Makpol, S.; Yusof, Y.A.M. A Review on Dietary Intervention in Obesity Associated Colon Cancer. Asian Pac. J. Cancer Prev. 2019, 20, 1309–1319. [Google Scholar] [CrossRef]
- O’Neill, A.M.; Burrington, C.M.; Gillaspie, E.A.; Lynch, D.T.; Horsman, M.; Greene, M.W. High-fat Western diet–induced obesity contributes to increased tumor growth in mouse models of human colon cancer. Nutr. Res. 2016, 36, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fedewa, S.A.; Ahnen, D.J.; Meester, R.G.S.; Barzi, A.; Jemal, A. Colorectal cancer statistics, 2017. CA Cancer J. Clin. 2017, 67, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Tarasiuk, A.; Mosińska, P.; Fichna, J. The mechanisms linking obesity to colon cancer: An overview. Obes. Res. Clin. Pract. 2018, 12, 251–259. [Google Scholar] [CrossRef]
- Penrose, H.M.; Heller, S.; Cable, C.; Nakhoul, H.; Baddoo, M.; Flemington, E.; Crawford, S.E.; Savkovic, S.D. High-fat diet induced leptin and Wnt expression: RNA-sequencing and pathway analysis of mouse colonic tissue and tumors. Carcinogenesis 2017, 38, 302–311. [Google Scholar] [CrossRef]
- Zeng, H.; Ishaq, S.; Liu, Z.; Bukowski, M. Colonic aberrant crypt formation accompanies an increase of opportunistic pathogenic bacteria in C57BL/6 mice fed a high-fat diet. J. Nutr. Biochem. 2018, 54, 18–27. [Google Scholar] [CrossRef]
- Chen, H.-H.; Tseng, Y.J.; Wang, S.-Y.; Tsai, Y.-S.; Chang, C.-S.; Kuo, T.-C.; Yao, W.-J.; Shieh, C.-C.; Wu, C.-H.; Kuo, P.-H. The metabolome profiling and pathway analysis in metabolic healthy and abnormal obesity. Int. J. Obes. 2015, 39, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Palviainen, M.; Laukkanen, K.; Tavukcuoglu, Z.; Velagapudi, V.; Kärkkäinen, O.; Hanhineva, K.; Auriola, S.; Ranki, A.; Siljander, P. Cancer Alters the Metabolic Fingerprint of Extracellular Vesicles. Cancers 2020, 12, 3292. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, X. Power of metabolomics in biomarker discovery and mining mechanisms of obesity. Obes. Rev. 2012, 14, 344–349. [Google Scholar] [CrossRef]
- Ried, J.S.; Shin, S.-Y.; Krumsiek, J.; Illig, T.; Theis, F.J.; Spector, T.D.; Adamski, J.; Wichmann, H.-E.; Strauch, K.; Soranzo, N.; et al. Novel genetic associations with serum level metabolites identified by phenotype set enrichment analyses. Hum. Mol. Genet. 2014, 23, 5847–5857. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guijas, C.; Montenegro-Burke, J.R.; Warth, B.; Spilker, M.; Siuzdak, G. Metabolomics activity screening for identifying metabolites that modulate phenotype. Nat. Biotechnol. 2018, 36, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Zarei, I.; Oppel, R.C.; Borresen, E.C.; Brown, R.J.; Ryan, E.P. Modulation of plasma and urine metabolome in colorectal cancer survivors consuming rice bran. Integr. Food Nutr. Metab. 2019, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Garvey, W.T.; Newman, J.; Lok, K.H.; Hoppel, C.L.; Adams, S. Plasma Metabolomic Profiles Reflective of Glucose Homeostasis in Non-Diabetic and Type 2 Diabetic Obese African-American Women. PLoS ONE 2010, 5, e15234. [Google Scholar] [CrossRef]
- Piccolo, B.D.; Keim, N.L.; Fiehn, O.; Adams, S.H.; Van Loan, M.D.; Newman, J.W. Habitual physical activity and plasma metabolomic patterns distinguish individuals with low vs. high weight loss during controlled energy restriction. J. Nutr. 2015, 145, 681–690. [Google Scholar] [CrossRef]
- Hong, S.; Moreno-Navarrete, J.M.; Wei, X.; Kikukawa, Y.; Tzameli, I.; Prasad, D.; Lee, Y.; Asara, J.M.; Fernández-Real, J.M.; Maratos-Flier, E.; et al. Nicotinamide N-methyltransferase regulates hepatic nutrient metabolism through Sirt1 protein stabilization. Nat. Med. 2015, 21, 887–894. [Google Scholar] [CrossRef]
- Peh, H.Y.; Tan, W.S.D.; Liao, W.; Wong, W.F. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Nikas, I.P.; Paschou, S.A.; Ryu, H.S. The Role of Nicotinamide in Cancer Chemoprevention and Therapy. Biomolecules 2020, 10, 477. [Google Scholar] [CrossRef]
- Bin Sayeed, M.S.; Ameen, S.S. Beta-Sitosterol: A Promising but Orphan Nutraceutical to Fight against Cancer. Nutr. Cancer 2015, 67, 1214–1220. [Google Scholar] [CrossRef] [PubMed]
- Buqué, A.; Bloy, N.; Kroemer, G.; Galluzzi, L. Possible mechanisms of cancer prevention by nicotinamide. Br. J. Pharmacol. 2021, 178, 2034–2040. [Google Scholar] [CrossRef] [PubMed]
- Fania, L.; Mazzanti, C.; Campione, E.; Candi, E.; Abeni, D.; Dellambra, E. Role of Nicotinamide in Genomic Stability and Skin Cancer Chemoprevention. Int. J. Mol. Sci. 2019, 20, 5946. [Google Scholar] [CrossRef]
- Megaraj, V.; Ding, X.; Fang, C.; Kovalchuk, N.; Zhu, Y.; Zhang, Q.-Y. Role of Hepatic and Intestinal P450 Enzymes in the Metabolic Activation of the Colon Carcinogen Azoxymethane in Mice. Chem. Res. Toxicol. 2014, 27, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Takada, H.; Hirooka, T.; Hiramatsu, Y.; Yamamoto, M. Effect of beta-glucuronidase inhibitor on azoxymethane-induced colonic carcinogenesis in rats. Cancer Res. 1982, 42, 331–334. [Google Scholar]
- Yu, B.; Peng, X.-H.; Wang, L.-Y.; Wang, A.-B.; Su, Y.-Y.; Chen, J.-H.; Zhang, X.-W.; Zhao, D.-Z.; Wang, H.; Pang, D.-X.; et al. Abnormality of intestinal cholesterol absorption in ApcMin/+ mice with colon cancer cachexia. Int. J. Clin. Exp. Pathol. 2019, 12, 759–767. [Google Scholar]
- Broitman, S.; Cerda, S.; Wilkinson, J. Cholesterol metabolism and colon cancer. Prog. Food Nutr. Sci. 1993, 17, 1–40. [Google Scholar]
- Winawer, S.J.; Flehinger, B.J.; Buchalter, J.; Herbert, E.; Shike, M. Declining serum cholesterol levels prior to diagnosis of colon cancer. A time-trend, case-control study. JAMA 1990, 263, 2083–2085. [Google Scholar] [CrossRef]
- Chung, Y.W.; Han, D.S.; Park, Y.K.; Son, B.K.; Paik, C.H.; Lee, H.L.; Jeon, Y.C.; Sohn, J.H. Association of obesity, serum glucose and lipids with the risk of advanced colorectal adenoma and cancer: A case-control study in Korea. Dig. Liver Dis. 2006, 38, 668–672. [Google Scholar] [CrossRef]
- Hamada, K.; Umemoto, A.; Kajikawa, A.; Tanaka, M.; Seraj, M.J.; Nakayama, M.; Kubota, A.; Monden, Y. Mucosa-specific DNA adducts in human small intestine: A comparison with the colon. Carcinogenesis 1994, 15, 2677–2680. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, X.W.; Liu, D.B.; Han, C.Z.; Du, L.L.; Jing, J.X.; Wang, Y. Lipid levels in serum and cancerous tissues of colorectal cancer patients. World J. Gastroenterol. 2014, 20, 8646–8652. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Narita, T.; Komiya, M.; Fujii, G.; Hamoya, T.; Nakanishi, R.; Tamura, S.; Kurokawa, Y.; Takahashi, M.; Mutoh, M. Novel screening system revealed that intracellular cholesterol trafficking can be a good target for colon cancer prevention. Sci. Rep. 2019, 9, 6192. [Google Scholar] [CrossRef] [PubMed]
- Myant, N. The Biology of Cholesterol and Related Steroids; Heinemann Medical Books: London, UK, 1981; p. 910. [Google Scholar]
- Wang, X.; Guan, L.; Zhao, Y.; Lei, L.; Liu, Y.; Ma, K.Y.; Wang, L.; Man, S.W.; Wang, J.; Huang, Y.; et al. Plasma cholesterol-lowering activity of dietary dihydrocholesterol in hypercholesterolemia hamsters. Atherosclerosis 2015, 242, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Skrede, S.; Björkhem, I.; Buchmann, M.S.; Hopen, G.; Fausa, O. A novel pathway for biosynthesis of cholestanol with 7 alpha-hydroxylated C27-steroids as intermediates, and its importance for the accumulation of cholestanol in cerebrotendinous xanthomatosis. J. Clin. Investig. 1985, 75, 448–455. [Google Scholar] [CrossRef]
- Lütjohann, D.; Björkhem, I.; Friedrichs, S.; Kerksiek, A.; Lövgren-Sandblom, A.; Geilenkeuser, W.-J.; Ahrends, R.; Andrade, I.; Ansorena, D.; Astiasarán, I.; et al. First international descriptive and interventional survey for cholesterol and non-cholesterol sterol determination by gas- and liquid-chromatography-Urgent need for harmonisation of analytical methods. J. Steroid Biochem. Mol. Biol. 2019, 190, 115–125. [Google Scholar] [CrossRef]
- Anderson, N.M.; Mucka, P.; Kern, J.G.; Feng, H. The emerging role and targetability of the TCA cycle in cancer metabolism. Protein Cell 2018, 9, 216–237. [Google Scholar] [CrossRef]
- Cho, S.; Song, N.; Choi, J.-Y.; Shin, A. Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank. Cancers 2020, 12, 2939. [Google Scholar] [CrossRef]
- Tan, B.; Qiu, Y.; Zou, X.; Chen, T.; Xie, G.; Cheng, Y.; Dong, T.; Zhao, L.; Feng, B.; Hu, X.; et al. Metabonomics Identifies Serum Metabolite Markers of Colorectal Cancer. J. Proteome Res. 2013, 12, 3000–3009. [Google Scholar] [CrossRef]
- Cui, H.; Yang, X.; Wang, Z.; Li, G.; Li, L.; Huo, S.; Zhang, B.; He, R.; Chen, K.; Xu, B.; et al. Tetrahydropalmatine triggers angiogenesis via regulation of arginine biosynthesis. Pharmacol. Res. 2021, 163, 105242. [Google Scholar] [CrossRef]
- Mirando, A.C.; Abdi, K.; Wo, P.; Lounsbury, K.M. Assessing the effects of threonyl-tRNA synthetase on angiogenesis-related responses. Methods 2017, 113, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Gu, J.; Chang, X.; Liu, F.; Liang, Y.; Yang, X.; Liang, L.; Tang, D. Metabonomics study on orthotopic transplantion mice model of colon cancer treated with Astragalus membranaceus-Curcuma wenyujin in different proportions via UPLC-Q-TOF/MS. J. Pharm. Biomed. Anal. 2021, 193, 113708. [Google Scholar] [CrossRef] [PubMed]
- Reitzer, L. Biosynthesis of Glutamate, Aspartate, Asparagine, L -Alanine, and D -Alanine. EcoSal Plus 2004, 1. [Google Scholar] [CrossRef] [PubMed]
- Padidar, S.; Farquharson, A.J.; Williams, L.M.; Kearney, R.; Arthur, J.R.; Drew, J.E. High-Fat Diet Alters Gene Expression in the Liver and Colon: Links to Increased Development of Aberrant Crypt Foci. Dig. Dis. Sci. 2012, 57, 1866–1874. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; Van Der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | AIN | AIN + AOM | HFD | HFD + AOM |
|---|---|---|---|---|
| Palmitoleic acid | 1.00 ± 0.40 a | 1.11 ± 0.27 a | 0.19 ± 0.05 b | 0.30 ± 0.11 b |
| Myristic acid | 1.00 ± 0.18 a | 1.02 ± 0.10 a | 0.57 ± 0.06 c | 0.74 ± 0.12 b |
| Oleic acid | 1.00 ± 1.46 bc | 0.73 ± 1.32 c | 1.99 ± 0.42 ab | 2.71 ± 0.95 a |
| Malic acid | 1.00 ± 0.26 a | 1.23 ± 0.42 a | 0.63 ± 0.08 b | 0.60 ± 0.12 b |
| Beta-sitosterol | 1.00 ± 0.35 b | 0.46 ± 0.11 c | 2.22 ± 0.65 a | 0.64 ± 0.10 bc |
| Oxoproline | 1.00 ± 0.14 b | 1.08 ± 0.15 b | 1.13 ± 0.17 b | 1.42 ± 0.21 a |
| Alpha-tocopherol | 1.00 ± 0.23 b | 0.67 ± 0.10 c | 1.94 ± 0.46 a | 0.84 ± 0.17 bc |
| Alpha-ketoglutarate | 1.00 ± 0.22 a | 1.15 ± 0.34 a | 0.72 ± 0.09 b | 0.63 ± 0.13 b |
| Fumaric acid | 1.00 ± 0.14 a | 1.17 ± 0.32 a | 0.76 ± 0.05 b | 0.75 ± 0.11 b |
| Citric acid | 1.00 ± 0.24 ab | 1.08 ± 0.14 a | 0.68 ± 0.07 c | 0.88 ± 0.10 b |
| 3-hydroxybutyric acid | 1.00 ± 0.64 c | 1.32 ± 0.62 bc | 2.08 ± 0.50 a | 1.80 ± 0.57 ab |
| Saccharic acid | 1.00 ± 0.18 c | 1.14 ± 0.21 bc | 1.49 ± 0.36 a | 1.33 ± 0.21 ab |
| Citrulline | 1.00 ± 0.21 a | 0.87 ± 0.17 ab | 0.77 ± 0.13 bc | 0.64 ± 0.12 c |
| 2-hydroxyglutaric acid | 1.00 ± 0.13 ab | 1.12 ± 0.17 a | 0.86 ± 0.11 b | 0.87 ± 0.14 b |
| 2-aminobutyric acid | 1.00 ± 0.37 b | 0.87 ± 0.36 b | 0.88 ± 0.24 b | 1.58 ± 0.54 a |
| Palmitic acid | 1.00 ± 0.15 ab | 1.09 ± 0.14 a | 0.86 ± 0.08 b | 0.88 ± 0.13 b |
| 3-ureidopropionate | 1.00 ± 0.29 b | 1.35 ± 0.49 b | 1.19 ± 0.47 b | 2.02 ± 0.55 a |
| Myo-inositol | 1.00 ± 0.24 b | 0.97 ± 0.13 b | 1.37 ± 0.32 a | 1.09 ± 0.27 b |
| Dihydrocholesterol | 1.00 ± 0.26 b | 0.48 ± 0.12 c | 1.89 ± 0.55 a | 0.48 ± 0.12 c |
| Glucuronic acid | 1.00 ± 0.33 c | 1.98 ± 0.51 ab | 1.32 ± 0.37 bc | 2.48 ± 0.96 a |
| Lysine | 1.00 ± 0.37 a | 0.77 ± 0.22 ab | 0.57 ± 0.25 b | 0.58 ± 0.15 b |
| Cholesterol | 1.00 ± 0.14 b | 0.79 ± 0.14 c | 1.32 ± 0.22 a | 0.81 ± 0.12 c |
| Isothreonic acid | 1.00 ± 0.09 b | 1.07 ± 0.08 b | 1.22 ± 0.20 a | 1.03 ± 0.06 b |
| Uracil | 1.00 ± 0.33 b | 1.04 ± 0.15 b | 0.95 ± 0.17 b | 1.29 ± 0.23 a |
| Ornithine | 1.00 ± 0.27 b | 1.39 ± 0.62 b | 1.05 ± 0.46 b | 2.04 ± 0.33 a |
| Succinic acid | 1.00 ± 0.21 ab | 1.27 ± 0.47 a | 0.87 ± 0.11 b | 0.87 ± 0.24 b |
| N-acetyl-d-tryptophan | 1.00 ± 0.22 a | 1.00 ± 0.26 a | 0.68 ± 0.22 b | 0.87 ± 0.18 ab |
| Pyruvic acid | 1.00 ± 0.45 ab | 1.15 ± 0.65 a | 0.58 ± 0.18 b | 0.67 ± 0.34 b |
| 2,3-dihydroxybutanoic acid | 1.00 ± 0.19 b | 1.11 ± 0.12 ab | 1.00 ± 0.12 b | 1.29 ± 0.24 a |
| Linoleic acid | 1.00 ± 0.26 b | 1.10 ± 0.26 ab | 0.99 ± 0.20 b | 1.41 ± 0.50 a |
| Beta-alanine | 1.00 ± 0.32 a | 0.97 ± 0.33 a | 0.60 ± 0.21 b | 0.86 ± 0.23 ab |
| Maleimide | 1.00 ± 0.13 b | 0.88 ± 0.14 b | 1.26 ± 0.14 a | 0.85 ± 0.14 b |
| Isocitric acid | 1.00 ± 0.17 a | 1.15 ± 0.23 a | 0.77 ± 0.11 b | 1.08 ± 0.14 a |
| Tyrosine | 1.00 ± 0.19 a | 1.12 ± 0.20 a | 0.75 ± 0.22 b | 1.06 ± 0.14 a |
| Xylitol | 1.00 ± 0.54 b | 1.58 ± 0.42 a | 0.90 ± 0.23 b | 2.03 ± 0.65 a |
| Nicotinamide | 1.00 ± 0.26 b | 0.92 ± 0.20 bc | 1.58 ± 0.24 a | 0.76 ± 0.15 c |
| Putrescine | 1.00 ± 0.34 a | 0.77 ± 0.25 ab | 0.70 ± 0.10 b | 0.70 ± 0.12 b |
| Threonic acid | 1.00 ± 0.15 b | 1.16 ± 0.24 ab | 1.39 ± 0.22 a | 1.01 ± 0.24 b |
| Methanolphosphate | 1.00 ± 0.12 b | 0.69 ± 0.11 c | 1.18 ± 0.15 a | 0.65 ± 0.09 c |
| Threonine | 1.00 ± 0.23 a | 0.76 ± 0.13 b | 0.74 ± 0.16 b | 0.75 ± 0.12 b |
| Serine | 1.00 ± 0.22 ab | 0.82 ± 0.14 b | 0.80 ± 0.21 b | 1.02 ± 0.14 a |
| Glycine | 1.00 ± 0.32 a | 0.82 ± 0.12 ab | 0.78 ± 0.09 b | 0.80 ± 0.12 b |
| Salicylic acid | 1.00 ± 0.16 ab | 1.15 ± 0.21 ab | 0.95 ± 0.16 b | 1.20 ± 0.19 a |
| Erythritol | 1.00 ± 0.13 b | 1.13 ± 0.07 a | 1.03 ± 0.10 b | 1.08 ± 0.06 ab |
| Sophorose | 1.00 ± 0.32 ab | 0.88 ± 0.29 b | 1.28 ± 0.44 a | 0.78 ± 0.18 b |
| Thymidine | 1.00 ± 0.21 ab | 1.03 ± 0.24 a | 0.79 ± 0.13 b | 1.20 ± 0.20 a |
| Aminomalonate | 1.00 ± 0.34 a | 0.76 ± 0.14 bc | 0.93 ± 0.25 ab | 0.67 ± 0.11 c |
| Arachidonic acid | 1.00 ± 0.11 a | 0.71 ± 0.14 b | 1.06 ± 0.16 a | 0.60 ± 0.10 b |
| Aconitic acid | 1.00 ± 0.35 ab | 1.09 ± 0.32 a | 0.75 ± 0.30 b | 1.14 ± 0.25 a |
| Phenylalanine | 1.00 ± 0.19 ab | 0.97 ± 0.24 ab | 0.81 ± 0.31 b | 1.13 ± 0.20 ab |
| Aspartic acid | 1.00 ± 0.33 ab | 1.01 ± 0.28 ab | 0.72 ± 0.20 b | 1.13 ± 0.27 a |
| Methionine | 1.00 ± 0.24 ab | 0.94 ± 0.26 ab | 0.73 ± 0.30 b | 1.08 ± 0.23 a |
| Glycerol-alpha-phosphate | 1.00 ± 0.29 a | 0.79 ± 0.08 b | 0.89 ± 0.14 ab | 0.79 ± 0.10 b |
| KEGG Pathway | Number of Metabolites Identified | p * | Impact ** |
|---|---|---|---|
| Citrate cycle (TCA cycle) | 7 | <0.0001 | 0.35 |
| Arg biosynthesis | 5 | <0.003 | 0.29 |
| Aminoacyl-tRNA biosynthesis | 8 | <0.005 | 0.17 |
| Ala, Asp, and Glu metabolism | 6 | <0.01 | 0.27 |
| Glyoxylate and dicarboxylate metabolism | 6 | <0.03 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, H.; Umar, S.; Liu, Z.; Bukowski, M.R. Azoxymethane Alters the Plasma Metabolome to a Greater Extent in Mice Fed a High-Fat Diet Compared to an AIN-93 Diet. Metabolites 2021, 11, 448. https://doi.org/10.3390/metabo11070448
Zeng H, Umar S, Liu Z, Bukowski MR. Azoxymethane Alters the Plasma Metabolome to a Greater Extent in Mice Fed a High-Fat Diet Compared to an AIN-93 Diet. Metabolites. 2021; 11(7):448. https://doi.org/10.3390/metabo11070448
Chicago/Turabian StyleZeng, Huawei, Shahid Umar, Zhenhua Liu, and Michael R. Bukowski. 2021. "Azoxymethane Alters the Plasma Metabolome to a Greater Extent in Mice Fed a High-Fat Diet Compared to an AIN-93 Diet" Metabolites 11, no. 7: 448. https://doi.org/10.3390/metabo11070448
APA StyleZeng, H., Umar, S., Liu, Z., & Bukowski, M. R. (2021). Azoxymethane Alters the Plasma Metabolome to a Greater Extent in Mice Fed a High-Fat Diet Compared to an AIN-93 Diet. Metabolites, 11(7), 448. https://doi.org/10.3390/metabo11070448