
Untargeted Metabolomics Uncovers the Essential Lysine Transporter in Toxoplasma gondii

 ,
,  ,
, 
{kind=link}
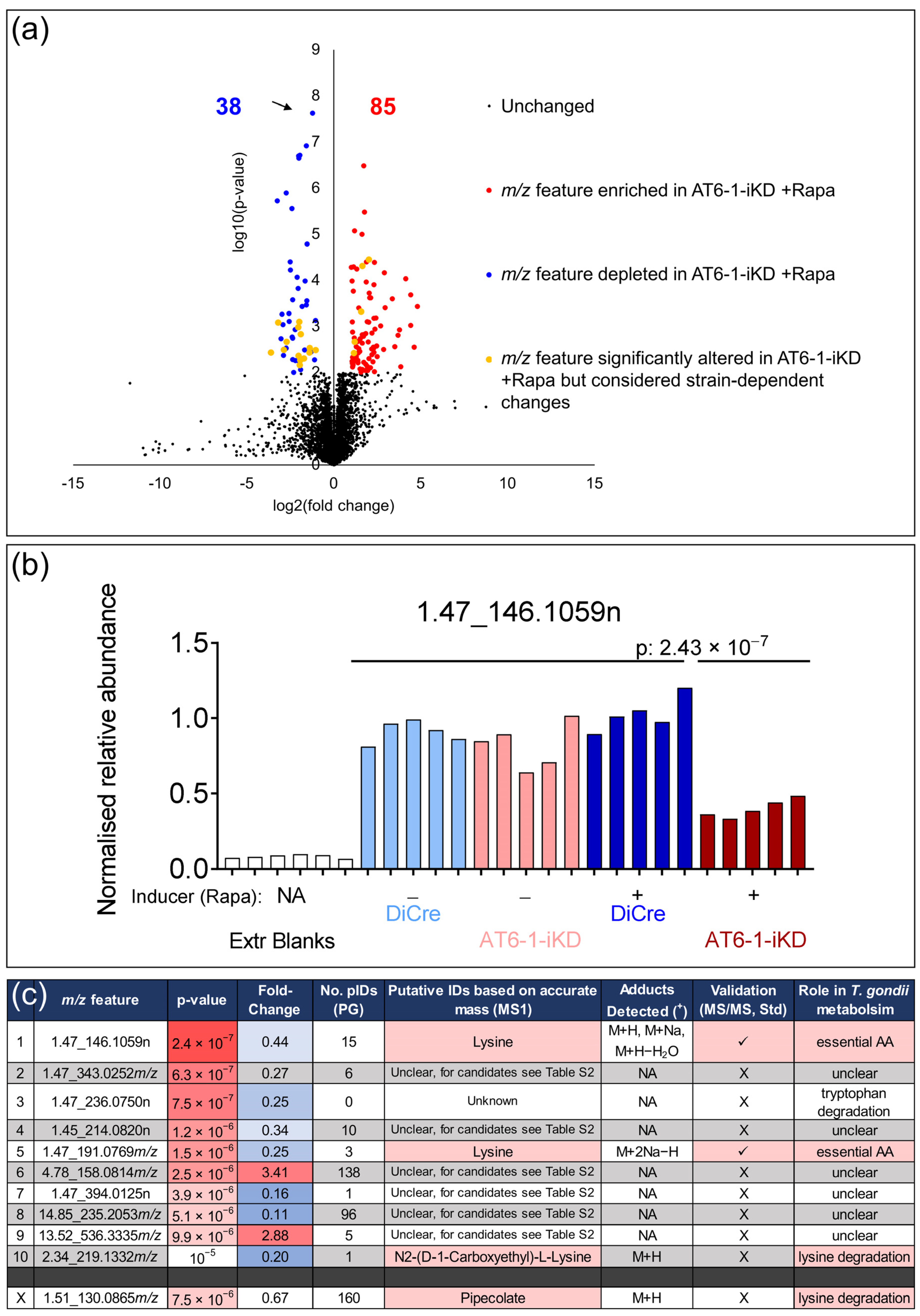{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results

3. Discussion
4. Materials and Methods
4.1. Parasite Culture and Maintenance
4.2. Cloning of DNA Constructs
4.3. Genomic DNA Extraction and Integration PCR
4.4. Plaque Assays and Intracellular Growth Assays
4.5. Immunofluorescence Assays
4.6. Western Blots
4.7. UHPLC-MS/MS Sample Preparation
U-13C Amino Acid Labeling and Sample Preparation
4.8. UHPLC-MS/MS Analysis
4.9. MS Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flegr, J.; Prandota, J.; Sovickova, M.; Israili, Z.H. Toxoplasmosis—A global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS ONE 2014, 9, e90203. [Google Scholar] [CrossRef] [Green Version]
- Furtado, J.M.; Smith, J.R.; Belfort, R., Jr.; Gattey, D.; Winthrop, K.L. Toxoplasmosis: A global threat. J. Glob. Infect. Dis. 2011, 3, 281–284. [Google Scholar] [CrossRef]
- Nissapatorn, V. Toxoplasma gondii and HIV: A never-ending story. Lancet HIV 2017, 4, e146–e147. [Google Scholar] [CrossRef]
- McAuley, J.B. Congenital toxoplasmosis. J. Pediatric Infect. Dis. Soc. 2014, 3, S30–S35. [Google Scholar] [CrossRef]
- Pittman, K.J.; Knoll, L.J. Long-Term relationships: The complicated interplay between the host and the developmental stages of Toxoplasma gondii during acute and chronic infections. Microbiol. Mol. Biol. Rev. 2015, 79, 387–401. [Google Scholar] [CrossRef] [Green Version]
- Blume, M.; Seeber, F. Metabolic interactions between toxoplasma gondii and its host. F1000Res. 2018, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, P.; Major, P.; Nakjang, S.; Hirt, R.P.; Embley, T.M. Transport proteins of parasitic protists and their role in nutrient salvage. Front. Plant Sci. 2014, 5, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.E. The transportome of the malaria parasite. Biol. Rev. Camb. Philos. Soc. 2020, 95, 305–332. [Google Scholar] [CrossRef] [PubMed]
- Blume, M.; Rodriguez-Contreras, D.; Landfear, S.; Fleige, T.; Soldati-Favre, D.; Lucius, R.; Gupta, N. Host-derived glucose and its transporter in the obligate intracellular pathogen Toxoplasma gondii are dispensable by glutaminolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 12998–13003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, J.C.; Afifi Afifi, M.; Pizzorno, G.; Handschumacher, R.E.; Joiner, K.A. Toxoplasma gondii tachyzoites possess an unusual plasma membrane adenosine transporter. Mol. Biochem. Parasitol. 1995, 70, 59–69. [Google Scholar] [CrossRef]
- Parker, K.E.R.; Fairweather, S.J.; Rajendran, E.; Blume, M.; McConville, M.J.; Broer, S.; Kirk, K.; van Dooren, G.G. The tyrosine transporter of Toxoplasma gondii is a member of the newly defined apicomplexan amino acid transporter (ApiAT) family. PLoS Pathog. 2019, 15, e1007577. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, E.; Hapuarachchi, S.V.; Miller, C.M.; Fairweather, S.J.; Cai, Y.; Smith, N.C.; Cockburn, I.A.; Broer, S.; Kirk, K.; van Dooren, G.G. Cationic amino acid transporters play key roles in the survival and transmission of apicomplexan parasites. Nat. Commun. 2017, 8, 14455. [Google Scholar] [CrossRef] [Green Version]
- Massimine, K.M.; Doan, L.T.; Atreya, C.A.; Stedman, T.T.; Anderson, K.S.; Joiner, K.A.; Coppens, I. Toxoplasma gondii is capable of exogenous folate transport. A likely expansion of the BT1 family of transmembrane proteins. Mol. Biochem. Parasitol. 2005, 144, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Quistgaard, E.M.; Low, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi, P.M.; Maco, B.; Dogga, S.K.; Frenal, K.; Soldati-Favre, D. Toxoplasma gondii TFP1 is an essential transporter family protein critical for microneme maturation and exocytosis. Mol. Microbiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A genome-wide CRISPR Screen in Toxoplasma identifies essential apicomplexan genes. Cell 2016, 166, 1423–1435. [Google Scholar] [CrossRef] [Green Version]
- Barylyuk, K.; Koreny, L.; Ke, H.; Butterworth, S.; Crook, O.M.; Lassadi, I.; Gupta, V.; Tromer, E.; Mourier, T.; Stevens, T.J.; et al. A comprehensive subcellular atlas of the Toxoplasma proteome via hyperlopit provides spatial context for protein functions. Cell Host Microbe 2020, 28, 752–766.e9. [Google Scholar] [CrossRef]
- Sangare, L.O.; Alayi, T.D.; Westermann, B.; Hovasse, A.; Sindikubwabo, F.; Callebaut, I.; Werkmeister, E.; Lafont, F.; Slomianny, C.; Hakimi, M.A.; et al. Unconventional endosome-like compartment and retromer complex in Toxoplasma gondii govern parasite integrity and host infection. Nat. Commun. 2016, 7, 11191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erler, H.; Ren, B.; Gupta, N.; Beitz, E. The intracellular parasite Toxoplasma gondii harbors three druggable FNT-type formate and l-lactate transporters in the plasma membrane. J. Biol. Chem. 2018, 293, 17622–17630. [Google Scholar] [CrossRef] [Green Version]
- Kloehn, J.; Oppenheim, R.D.; Siddiqui, G.; De Bock, P.J.; Kumar Dogga, S.; Coute, Y.; Hakimi, M.A.; Creek, D.J.; Soldati-Favre, D. Multi-omics analysis delineates the distinct functions of sub-cellular acetyl-CoA pools in Toxoplasma gondii. BMC Biol. 2020, 18, 67. [Google Scholar] [CrossRef] [PubMed]
- MacRae, J.I.; Sheiner, L.; Nahid, A.; Tonkin, C.; Striepen, B.; McConville, M.J. Mitochondrial metabolism of glucose and glutamine is required for intracellular growth of Toxoplasma gondii. Cell Host Microbe 2012, 12, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Hunt, A.; Russell, M.R.G.; Wagener, J.; Kent, R.; Carmeille, R.; Peddie, C.J.; Collinson, L.; Heaslip, A.; Ward, G.E.; Treeck, M.; et al. Differential requirements for cyclase-associated protein (CAP) in actin-dependent processes of Toxoplasma gondii. Elife 2019, 8. [Google Scholar] [CrossRef]
- Bisio, H.; Chaabene, R.B.; Sabitzki, R.; Maco, B.; Marq, J.B.; Gilberger, T.W.; Spielmann, T.; Soldati-Favre, D. The ZIP code of vesicle trafficking in Apicomplexa: SEC1/Munc18 and SNARE proteins. mBio 2020, 11. [Google Scholar] [CrossRef]
- Pieperhoff, M.S.; Pall, G.S.; Jimenez-Ruiz, E.; Das, S.; Melatti, C.; Gow, M.; Wong, E.H.; Heng, J.; Muller, S.; Blackman, M.J.; et al. Conditional U1 gene silencing in Toxoplasma gondii. PLoS ONE 2015, 10, e0130356. [Google Scholar] [CrossRef] [Green Version]
- Fairweather, S.J.; Rajendran, E.; Blume, M.; Javed, K.; Steinhofel, B.; McConville, M.J.; Kirk, K.; Broer, S.; Van Dooren, G. Coordinated action of multiple transporters in the acquisition of essential cationic amino acids by the intracellular parasite Toxoplasma gondii. BioRxiv 2021. [Google Scholar] [CrossRef]
- Rajendran, E.; Morgan, C.; Goulart, C.; Steinhofel, B.; Tjhin, E.T.; Smith, N.C.; Kirk, K.; Van Dooren, G. Substrate-mediated regulation of the arginine transporter of Toxoplasma gondii. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.; Soldati-Favre, D. Amino acid metabolism in apicomplexan parasites. Metabolites 2021, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Beri, D.; Ramdani, G.; Balan, B.; Gadara, D.; Poojary, M.; Momeux, L.; Tatu, U.; Langsley, G. Insights into physiological roles of unique metabolites released from Plasmodium-infected RBCs and their potential as clinical biomarkers for malaria. Sci. Rep. 2019, 9, 2875. [Google Scholar] [CrossRef]
- Creek, D.J.; Chua, H.H.; Cobbold, S.A.; Nijagal, B.; MacRae, J.I.; Dickerman, B.K.; Gilson, P.R.; Ralph, S.A.; McConville, M.J. Metabolomics-based screening of the malaria box reveals both novel and established mechanisms of action. Antimicrob. Agents Chemother. 2016, 60, 6650–6663. [Google Scholar] [CrossRef] [Green Version]
- Li, J.V.; Wang, Y.; Saric, J.; Nicholson, J.K.; Dirnhofer, S.; Singer, B.H.; Tanner, M.; Wittlin, S.; Holmes, E.; Utzinger, J.; et al. Global metabolic responses of NMRI mice to an experimental Plasmodium berghei infection. J. Proteome Res. 2008, 7, 3948–3956. [Google Scholar] [CrossRef]
- Naemat, A.; Elsheikha, H.M.; Boitor, R.A.; Notingher, I. Tracing amino acid exchange during host-pathogen interaction by combined stable-isotope time-Resolved Raman spectral imaging. Sci. Rep. 2016, 6, 20811. [Google Scholar] [CrossRef] [Green Version]
- Meier, A.; Erler, H.; Beitz, E. Targeting Channels and Transporters in Protozoan Parasite Infections. Front Chem 2018, 6, 88. [Google Scholar] [CrossRef] [Green Version]
- Staines, H.M.; Moore, C.M.; Slavic, K.; Krishna, S. Transmembrane solute transport in the apicomplexan parasite Plasmodium. Emerg. Top. Life Sci. 2017, 1, 553–561. [Google Scholar] [CrossRef]
- Cerutti, A.; Blanchard, N.; Besteiro, S. The bradyzoite: A key developmental stage for the persistence and pathogenesis of toxoplasmosis. Pathogens 2020, 9, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B.; Brown, K.M.; Lee, T.D.; Sibley, L.D. Efficient gene disruption in diverse strains of Toxoplasma gondii using CRISPR/CAS9. MBIO 2014, 5, e01114-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donald, R.G.; Carter, D.; Ullman, B.; Roos, D.S. Insertional tagging, cloning, and expression of the Toxoplasma gondii hypoxanthine-xanthine-guanine phosphoribosyltransferase gene. Use as a selectable marker for stable transformation. J. Biol. Chem. 1996, 271, 14010–14019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, A.; Kloehn, J.; Lunghi, M.; Chiappino-Pepe, A.; Waldman, B.S.; Nicolas, D.; Varesio, E.; Hehl, A.; Lourido, S.; Hatzimanikatis, V.; et al. Functional and computational genomics reveal unprecedented flexibility in stage-specific Toxoplasma metabolism. Cell Host Microbe 2020, 27, 290–306. [Google Scholar] [CrossRef] [PubMed]
- Plattner, F.; Yarovinsky, F.; Romero, S.; Didry, D.; Carlier, M.F.; Sher, A.; Soldati-Favre, D. Toxoplasma profilin is essential for host cell invasion and TLR11-dependent induction of an interleukin-12 response. Cell Host Microbe 2008, 3, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herm-Gotz, A.; Weiss, S.; Stratmann, R.; Fujita-Becker, S.; Ruff, C.; Meyhofer, E.; Soldati, T.; Manstein, D.J.; Geeves, M.A.; Soldati, D.; et al. Toxoplasma gondii myosin A and its light chain: A fast, single-headed, plus-end-directed motor. EMBO J. 2002, 21, 2149–2158. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; van Dooren, G.G.; Beatty, W.L.; Striepen, B. Genetic evidence that an endosymbiont-derived endoplasmic reticulum-associated protein degradation (ERAD) system functions in import of apicoplast proteins. J. Biol. Chem. 2009, 284, 33683–33691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pino, P.; Aeby, E.; Foth, B.J.; Sheiner, L.; Soldati, T.; Schneider, A.; Soldati-Favre, D. Mitochondrial translation in absence of local tRNA aminoacylation and methionyl tRNA met formylation in Apicomplexa. Mol. Microbiol. 2010, 76, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Prinsen, H.; Schiebergen-Bronkhorst, B.G.M.; Roeleveld, M.W.; Jans, J.J.M.; de Sain-van der Velden, M.G.M.; Visser, G.; van Hasselt, P.M.; Verhoeven-Duif, N.M. Rapid quantification of underivatized amino acids in plasma by hydrophilic interaction liquid chromatography (HILIC) coupled with tandem mass-spectrometry. J. Inherit. Metab. Dis. 2016, 39, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, K.J.; Pratt, B.; Bose, N.; Dubois, L.G.; St John-Williams, L.; Perrott, K.M.; Ky, K.; Kapahi, P.; Sharma, V.; MacCoss, M.J.; et al. Skyline for small molecules: A unifying software package for quantitative metabolomics. J. Proteome Res. 2020, 19, 1447–1458. [Google Scholar] [CrossRef]
- Clasquin, M.F.; Melamud, E.; Rabinowitz, J.D. LC-MS data processing with MAVEN: A metabolomic analysis and visualization engine. Curr. Protoc. Bioinform. 2012, Chapter 14, Unit14.11. [Google Scholar] [CrossRef] [Green Version]
- Zamboni, N.; Fendt, S.M.; Ruhl, M.; Sauer, U. (13)C-based metabolic flux analysis. Nat. Protoc. 2009, 4, 878–892. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kloehn, J.; Lunghi, M.; Varesio, E.; Dubois, D.; Soldati-Favre, D. Untargeted Metabolomics Uncovers the Essential Lysine Transporter in Toxoplasma gondii. Metabolites 2021, 11, 476. https://doi.org/10.3390/metabo11080476
Kloehn J, Lunghi M, Varesio E, Dubois D, Soldati-Favre D. Untargeted Metabolomics Uncovers the Essential Lysine Transporter in Toxoplasma gondii. Metabolites. 2021; 11(8):476. https://doi.org/10.3390/metabo11080476
Chicago/Turabian StyleKloehn, Joachim, Matteo Lunghi, Emmanuel Varesio, David Dubois, and Dominique Soldati-Favre. 2021. "Untargeted Metabolomics Uncovers the Essential Lysine Transporter in Toxoplasma gondii" Metabolites 11, no. 8: 476. https://doi.org/10.3390/metabo11080476
APA StyleKloehn, J., Lunghi, M., Varesio, E., Dubois, D., & Soldati-Favre, D. (2021). Untargeted Metabolomics Uncovers the Essential Lysine Transporter in Toxoplasma gondii. Metabolites, 11(8), 476. https://doi.org/10.3390/metabo11080476