
Integrated Lipidomic and Metabolomics Analysis Revealing the Effects of Frozen Storage Duration on Pork Lipids


Abstract
1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Preparation of Fatty Acids and Aldehydes Internal Standard Solution (IS)
2.3. Sampling
2.4. Untargeted Lipidomic Analysis
2.5. Fatty Acids Analysis
2.6. Fatty Aldehyde Analysis
2.7. Statistical Analysis
3. Results
3.1. Untargeted Lipidomic Profiling of the Changes in Pork during Freezing Storage
3.2. The Effect of Frozen Storage on Total and Free FAs in Pork
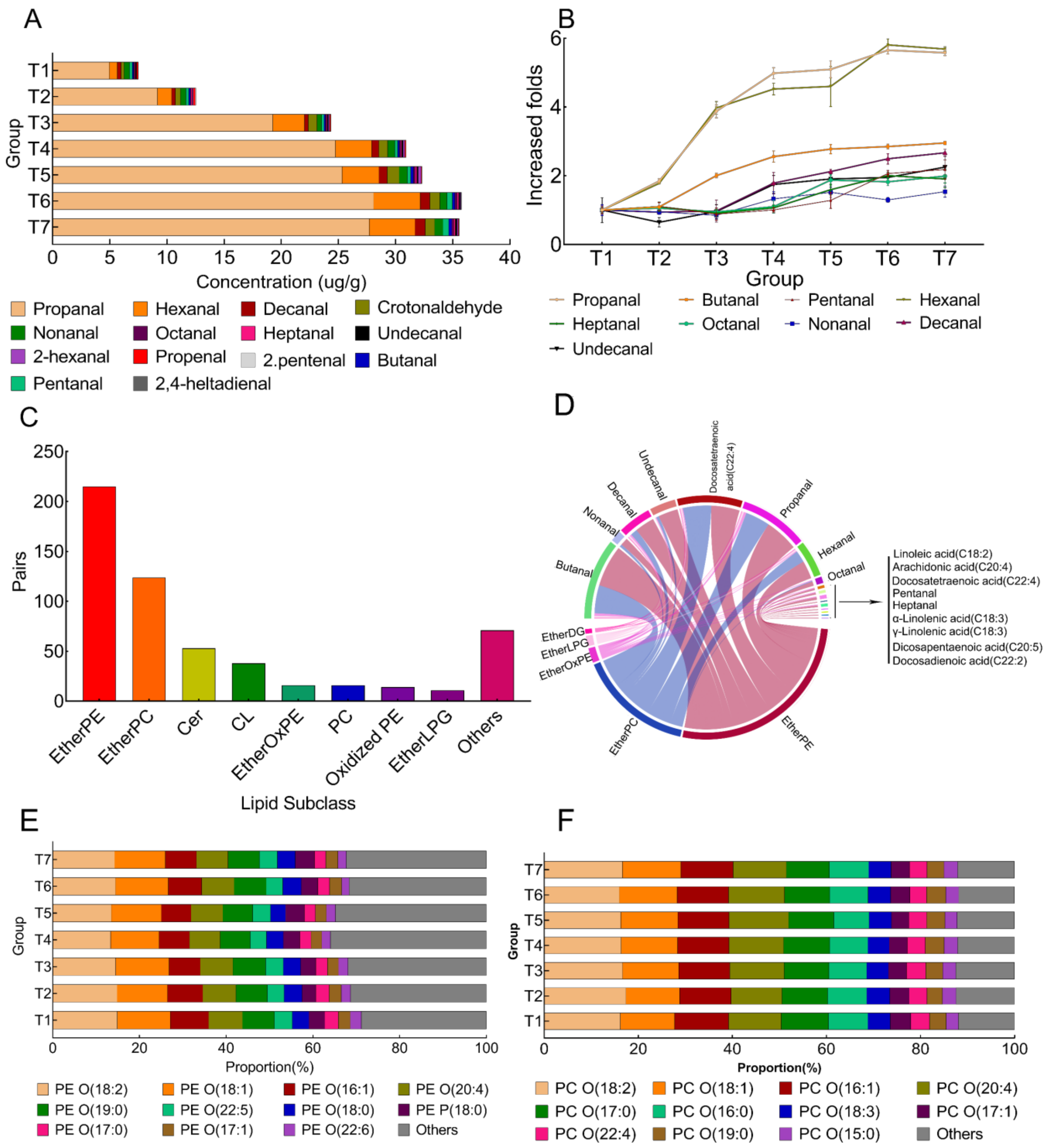
3.3. The Effect of Frozen Storage on Fatty Aldehydes in Pork
3.4. The Correlation in Changed Tendency of Free FAs and Fatty Aldehydes with Lipid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, J.; Zhang, J.Q.; Yang, Y.Y.; Zhu, J.W.; He, W.Z.; Zhao, Q.Y.; Tang, C.H.; Qin, Y.C.; Zhang, J.M. Comparative characterization of lipids and volatile compounds of Beijing Heiliu and Laiwu Chinese Black Pork as markers. Food Res. Int. 2021, 146, 110433–110443. [Google Scholar] [CrossRef] [PubMed]
- Al-Dalali, S.; Li, C.; Xu, B.C. Effect of frozen storage on the lipid oxidation, protein oxidation, and flavor profile of marinated raw beef meat. Food Chem. 2022, 376, 131881. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guo, M.Y.; Liao, E.; Wang, Q.; Peng, L.J.; Jin, W.P.; Wang, H.B. Effects of repeated freezing and thawing on myofibrillar protein and quality characteristics of marinated Enshi black pork. Food Chem. 2022, 378, 131994. [Google Scholar] [CrossRef] [PubMed]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Impact of freezing and thawing on the quality of meat: Review. Meat Sci. 2012, 91, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Medic, H.; Djurkin Kusec, I.; Pleadin, J.; Kozacinski, L.; Njari, B.; Hengl, B.; Kusec, G. The impact of frozen storage duration on physical, chemical and microbiological properties of pork. Meat Sci. 2018, 140, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Farouk, M.M.; Wieliczko, K.J. Effect of diet and fat content on the functional properties of thawed beef. Meat Sci. 2003, 64, 451–458. [Google Scholar] [CrossRef]
- Alonso, V.; Muela, E.; Tenas, J.; Calanche, J.B.; Roncalés, P.; Beltrán, J.A. Changes in physicochemical properties and fatty acid composition of pork following long-term frozen storage. Eur. Food Res. Technol. 2016, 242, 2119–2127. [Google Scholar]
- Holman, B.W.B.; Coombs, C.E.O.; Morris, S.; Kerr, M.J.; Hopkins, D.L. Effect of long term chilled (up to 5 weeks) then frozen (up to 12months) storage at two different sub-zero holding temperatures on beef: 1. meat quality and microbial loads. Meat Sci. 2017, 133, 133–142. [Google Scholar] [CrossRef]
- Coombs, C.E.O.; Holman, B.W.B.; Collins, D.; Friend, M.A.; Hopkins, D.L. Effects of chilled-then-frozen storage (up to 52 weeks) on lamb M. longissimus lumborum quality and safety Parameters. Meat Sci. 2017, 134, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.-M.; Peng, Z.-Y.; Lu, S.; He, H.-J.; Kang, Z.-L.; Ma, H.-J.; Zhao, S.-M.; Wang, Z.-R. Physicochemical properties and protein denaturation of pork longissimus dorsi muscle subjected to six microwave-based thawing methods. Foods 2020, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhang, J.; Jiao, L.; Ma, C.; Kang, Z.; Ma, H. Effects of freezing methods and frozen storage on physicochemical, oxidative properties and protein denaturation of prcine longissimus dorsi. LWT-Food Sci. Technol. 2022, 153, 112529–112536. [Google Scholar] [CrossRef]
- Lee, S.; Kim, E.; Park, D.; Ji, Y.; Kang, G.; Choi, M. Deep freezing to maintain the freshness of pork loin during long-term storage. Food Sci. Biotechnol. 2021, 30, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhong, Q.; Kong, B.; Wang, B.; Pan, N.; Xia, X. Deterioration in quality of quick-frozen pork patties induced by changes in protein structure and lipid and protein oxidation during frozen storage. Food Res. Int. 2020, 133, 109142–109153. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Saha, A.; Xiong, R.; Owens, C.M.; Meullenet, J.F. Changes in broiler breast fillet tenderness, water-holding capacity, and color attributes during long-term frozen storage. J. Food Sci. 2008, 73, E162–E168. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, R.S.B.; Francisco, C.L.; Lino, D.M.; Borba, H. Meat Quality of Santa Inês lamb chilled-then-frozen storage up to 12 months. Meat Sci. 2019, 148, 72–78. [Google Scholar] [CrossRef]
- Xia, X.F.; Kong, B.H.; Liu, Q.; Liu, J. Physicochemical change and protein oxidation in porcine longissimus dorsi as influenced by different freeze–thaw cycles. Meat Sci. 2009, 83, 239–245. [Google Scholar] [CrossRef]
- Huang, L.; Youling, L.X.; Baohua, K.; Xiangang, H.; Jing, L. Influence of storage temperature and duration on lipid and protein oxidation and flavour changes in frozen pork dumpling filler. Meat Sci. 2013, 95, 295–301. [Google Scholar] [CrossRef]
- Coombs, C.E.O.; Holman, B.W.B.; Friend, M.A.; Hopkins, D.L. Long-term red meat preservation using chilled and frozen storage combinations: A review. Meat Sci. 2017, 125, 84–94. [Google Scholar] [CrossRef]
- Lu, N.; Ma, J.; Sun, D.-W. Enhancing physical and chemical quality attributes of frozen meat and meat products: Mechanisms, techniques and applications. Trends Food Sci. Technol. 2022, 124, 63–85. [Google Scholar] [CrossRef]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid-protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef]
- Soyer, A.; Özalp, B.; Dalmış, Ü.; Bilgin, V. Effects of freezing temperature and duration of frozen storage on lipid and protein oxidation in chicken meat. Food Chem. 2010, 120, 1025–1030. [Google Scholar] [CrossRef]
- Tatiyaborworntham, N.; Oz, F.; Richards, M.P.; Wu, H.Z. Paradoxical effects of lipolysis on the lipid oxidation in meat and meat products. Food Chem. X 2022, 14, 100317. [Google Scholar] [CrossRef] [PubMed]
- Schaur, R.J.; Siems, W.; Bresgen, N.; Eckl, P.M. 4-Hydroxy-nonenal-A bioactive lipid peroxidation product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef]
- Ko, C.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V. Mechanisms of oxidative processes in meat and toxicity induced by postprandial degradation products: A review. Compr. Rev. Food Sci. F 2017, 16, 96–123. [Google Scholar] [CrossRef] [PubMed]
- Coombs, C.E.O.; Holman, B.W.B.; Ponnampalam, E.N.; Morris, S.; Friend, M.A.; Hopkins, D.L. Effects of chilled and frozen storage conditions on the lamb M. longissimus lumborum fatty acid and lipid oxidation parameters. Meat Sci. 2018, 136, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Cao, H.; Lin, H.; Deng, S.; Wu, H. Insights into ice-growth inhibition by trehalose and alginate oligosaccharides in peeled pacific white shrimp (litopenaeus vannamei) during frozen storage. Food Chem. 2019, 278, 482–490. [Google Scholar] [CrossRef]
- Liu, Y.; Jiao, J.; Gao, S.; Ning, L.; McHele Limbu, S.; Qiao, F.; Chen, L.; Zhang, M.; Du, Z. Dietary oils modify lipid molecules and nutritional value of fillet in nile tilapia: A deep lipidomics analysis. Food Chem. 2019, 277, 515–523. [Google Scholar] [CrossRef]
- He, C.; Cao, J.; Bao, Y.; Sun, Z.; Liu, Z.; Li, C. Characterization of lipid profiling in three parts (muscle, head and viscera) of Tilapia (oreochromis niloticus) using lipidomics with UPLC-ESI-Q-TOF-MS. Food Chem. 2021, 347, 129057–129066. [Google Scholar] [CrossRef]
- Ai, Z.; Zhang, Y.; Li, X.; Sun, W.; Liu, Y. Widely targeted metabolomics analysis to reveal transformation mechanism of cistanche deserticola active compounds during steaming and drying processes. Front. Nutr. 2021, 8, 742511–742524. [Google Scholar] [CrossRef]
- Roberts, L.D.; Souza, A.L.; Gerszten, R.E.; Clish, C.B. Targeted metabolomics. Curr. Protoc. Mol. Biol. 2012, 30, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.H.; Tang, Z.H.; Chen, B.Y.; Feng, S.N.; Liu, Y.; Meng, Q.S. A high-efficiency quantitation method for fatty aldehyde based on chemical isotope-tagged derivatisation. J. Food Compos. Anal. 2022, 107, 104381–104392. [Google Scholar] [CrossRef]
- Feng, X.H.; Liu, N.; Yang, Y.Y.; Feng, S.N.; Wang, J.; Meng, Q.S. Isotope-coded chemical derivatization method for highly accurately and sensitively quantifying short-chain fatty acids. J. Agric. Food Chem. 2022, 70, 6253–6263. [Google Scholar] [CrossRef]
- Feng, X.H.; Wang, J.; Tang, Z.H.; Chen, B.Y.; Hou, X.H.; Li, J.; Feng, S.N.; Li, P.; Meng, Q.S. A strategy for accurately and sensitively quantifying free and esterified fatty acids using liquid chromatography mass spectrometry. Front. Nutr. 2022, 9, 977076. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash sample-to-extraction solvent ratios for human plasma-based lipidomics studies. Anal. Chim. Acta 2018, 1037, 351–357. [Google Scholar] [CrossRef]
- Tsugawa, H.; Ikeda, K.; Takahashi, M.; Satoh, A.; Mori, Y.; Uchino, H.; Okahashi, N.; Yamada, Y.; Tada, I.; Bonini, P.; et al. A lipidome atlas in MS-DIAL 4. Nat. Biotechnol. 2020, 38, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Kofeler, H.; et al. Update on LlIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Perez-Palacios, T.; Solomando, J.C.; Ruiz-Carrascal, J.; Antequera, T. Improvements in the Methodology for Fatty Acids Analysis in Meat Products: One-Stage Transmethylation and Fast-GC Method. Food Chem. 2022, 371, 130995. [Google Scholar] [CrossRef] [PubMed]
- Scerra, M.; Foti, F.; Caparra, P.; Cilione, C.; Rao, R.; Priolo, A.; Natalello, A.; Luciano, G.; Chies, L. Effect of feeding pigs with bergamot by-product on fatty acid composition and oxidative stability of meat and salami. Meat Sci. 2022, 183, 108662–108671. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Hu, Y.; Liu, Z.; Xie, H.; Zhang, M.; Zheng, R.; Qin, L.; Yin, F.; Zhou, D. Simultaneous quantification of 24 aldehydes and ketones in oysters (crassostrea gigas) with different thermal processing procedures by HPLC-electrospray tandem mass spectrometry. Food Res. Int. 2021, 147, 110559–110567. [Google Scholar] [CrossRef]
- Dominguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2019, 8, 429–460. [Google Scholar]
- Liu, Z.; Zhou, D.; Li, A.; Zhao, M.; Hu, Y.; Li, D.; Xie, H.; Zhao, Q.; Hu, X.; Zhang, J.; et al. Effects of temperature and heating time on the formation of aldehydes during the frying process of clam assessed by an HPLC-MS/MS method. Food Chem. 2020, 308, 125650–125658. [Google Scholar]
- Mi, S.; Shang, K.; Li, X.; Zhang, C.H.; Liu, J.Q.; Huang, D.Q. Characterization and discrimination of selected China’s domestic pork using an LC-MS-based lipidomics approach. Food Control 2019, 100, 305–314. [Google Scholar]
- Zhou, L.; Zhao, M.J.; Bindler, F.; Marchioni, E. Comparison of the volatiles formed by oxidation of phosphatidylcholine to triglyceride in model systems. J. Agric. Food Chem. 2014, 62, 8295–8301. [Google Scholar]
- Wu, N.; Wang, X.C. Identification of important odorants derived from phosphatidylethanolamine species in steamed male rriocheir sinensis hepatopancreas in model systems. Food Chem. 2019, 286, 491–499. [Google Scholar]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar]
- Broniec, A.; Klosinski, R.; Pawlak, A.; Wrona-Krol, M.; Thompson, D.; Sarna, T. Interactions of plasmalogens and their diacyl analogs with singlet oxygen in selected model systems. Free Radical Biol. Med. 2011, 50, 892–898. [Google Scholar] [CrossRef]
- Manful, C.F.; Pham, T.H.; Nadeem, M.; Wheeler, E.; Warren, K.J.T.; Vidal, N.P.; Thomas, R.H. Assessing unfiltered beer-based marinades effects on ether and ester linked phosphatidylcholines and phosphatidylethanolamines in grilled beef and moose meat. Meat Sci. 2021, 171, 108271–108285. [Google Scholar] [CrossRef] [PubMed]
- Broniec, A.; Zadlo, A.; Pawlak, A.; Fuchs, B.; Klosinski, R.; Thompson, D.; Sarna, T. Interaction of plasmenylcholine with free radicals in selected model systems. Free Radical Biol. Med. 2017, 106, 368–378. [Google Scholar] [CrossRef]
- Flakemore, A.R.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O.l. Omega-3 fatty acids, nutrient retention values, and sensory meat eating quality in cooked and raw Australian lamb. Meat Sci. 2017, 123, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ojha, K.S.; Perussello, C.A.; García, C.Á.; Kerry, J.P.; Pando, D.; Tiwari, B.K. Ultrasonic-assisted incorporation of nano-encapsulated omega-3 fatty acids to enhance the fatty acid profile of pork meat. Meat Sci. 2017, 132, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kumar, F.; Tyagi, P.K.; Mir, N.A.; Dev, K.; Begum, J.; Biswas, A.; Sheikh, S.A.; Tyagi, P.K.; Sharma, D.; Sahu, B.; et al. Dietary flaxseed and turmeric is a novel strategy to enrich chicken meat with long chain ω-3 polyunsaturated fatty acids with better oxidative stability and functional properties. Food Chem. 2020, 305, 125458–125466. [Google Scholar] [CrossRef] [PubMed]
- Sohail, A.; Al-Dalali, S.; Wang, J.; Xie, J.; Shakoor, A.; Asimi, S.; Shah, H.; Patil, P. Aroma compounds identified in cooked meat: A review. Food Res. Int. 2022, 157, 111385–111399. [Google Scholar] [CrossRef]
- Ross, C.F.; Smith, D.M. Use of volatiles as indicators of lipid oxidation in muscle foods. Compr. Rev. Food Sci. F 2006, 5, 18–25. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subclass | T1 | T2 | T3 | T4 | T5 | T6 | T7 | p Value |
|---|---|---|---|---|---|---|---|---|
| CAR | 7.41 ± 2.3 b | 11.82 ± 2.71 a | 12.38 ± 1.1 a | 10.02 ± 3.42 ab | 10.06 ± 2.13 ab | 11.66 ± 3.99 a | 10.91 ± 2.34 ab | 1.18 × 10−1 |
| Cer | 38.49 ± 5.51 a | 35.12 ± 2.95 a | 39.94 ± 3.04 a | 27.11 ± 5.92 b | 25.06 ± 3.01 b | 24.83 ± 3.34 b | 23.41 ± 5.03 b | 3.86 × 10−7 |
| CL | 33.05 ± 4.42 bc | 35.02 ± 3.18 b | 44.06 ± 1.87 a | 29.33 ± 4.26 cd | 25 ± 2.13 d | 25.14 ± 6.47 d | 19.29 ± 1.93 e | 1.43 × 10−9 |
| DG | 48.37 ± 3.88 d | 105.29 ± 15.88 ab | 65.86 ± 6.69 cd | 125.3 ± 30.05 ab | 127.2 ± 37.91 a | 128.95 ± 44.27 a | 90.61 ± 11.76 bc | 6.95 × 10−5 |
| EtherDG | 5.42 ± 0.71 c | 6.63 ± 0.25 b | 12.19 ± 0.4 a | 5.88 ± 0.38 c | 3.66 ± 0.13 d | 3.1 ± 0.37 e | 3.15 ± 0.18 e | 1.08 × 10−24 |
| EtherLPC | 4.17 ± 0.92 bc | 3.01 ± 0.89 c | 3.22 ± 0.46 c | 3.77 ± 0.91 bc | 2.95 ± 0.34 c | 6.57 ± 2.01 b | 9.81 ± 5.61 a | 4.46 × 10−4 |
| EtherLPE | 6.35 ± 0.92 ab | 5.27 ± 1.27 ab | 4.54 ± 0.47 b | 5 ± 1.18 ab | 4.88 ± 1.34 ab | 5.84 ± 3.52 ab | 7.5 ± 3 a | 2.66 × 10−1 |
| EtherLPG | 2.99 ± 0.33 b | 3.4 ± 0.38 ab | 2.87 ± 0.34 b | 2.73 ± 0.69 b | 2.81 ± 0.49 b | 4.55 ± 2.29 a | 2.52 ± 0.11 b | 4.01 × 10−2 |
| EtherOxPE | 5.37 ± 0.33 a | 5.42 ± 0.16 a | 5.32 ± 0.25 ab | 4.88 ± 1.81 abc | 3.42 ± 0.3 c | 4.44 ± 1.45 abc | 3.84 ± 1.52 bc | 2.90 × 10−2 |
| EtherPC | 106.6 ± 13.38 a | 94.35 ± 14.43 ab | 84.34 ± 3.68 bc | 69.78 ± 10.28 cd | 66.05 ± 9.64 d | 70.99 ± 17.73 cd | 68.31 ± 11.79 cd | 4.24 × 10−5 |
| EtherPE | 114.18 ± 15.74 a | 112.56 ± 14.01 a | 99.44 ± 7.75 a | 80.49 ± 11.14 b | 73.46 ± 11 b | 74.62 ± 13.74 b | 67.22 ± 6.23 b | 3.29 × 10−7 |
| EtherPG | 4.56 ± 0.13 bc | 5.34 ± 0.39 b | 6.55 ± 0.33 a | 4.66 ± 0.47 bc | 4.42 ± 0.4 c | 4.42 ± 1.28 c | 3.11 ± 0.4 d | 8.89 × 10−8 |
| FA | 23.82 ± 3.76 ab | 27.33 ± 3.6 a | 26.12 ± 2.75 ab | 22.26 ± 4.63 ab | 21.96 ± 2.68 ab | 22.34 ± 6.67 ab | 20.08 ± 6.02 b | 1.96 × 10−1 |
| FAM | 26.7 ± 1.7 d | 30.66 ± 1.62 b | 48.91 ± 3.35 a | 27.45 ± 2.15 cd | 30.06 ± 0.99 bc | 19.26 ± 1.31 e | 21.85 ± 3.14 e | 2.57 × 10−17 |
| LPC | 7.47 ± 0.66 c | 6.18 ± 1.46 bc | 6.54 ± 1.23 bc | 6.47 ± 1.89 bc | 4.94 ± 0.86 c | 7.96 ± 2.34 b | 10.61 ± 3.76 a | 4.91 × 10−3 |
| MG | 7.37 ± 0.34 c | 7.82 ± 0.27 c | 10.89 ± 0.23 a | 8.11 ± 0.84 c | 9.44 ± 0.6 b | 7.76 ± 1.9 c | 7.34 ± 0.44 c | 1.51 × 10−6 |
| Others | 23.19 ± 3.33 ab | 23.47 ± 3.84 ab | 25.42 ± 1.13 a | 19.71 ± 3.61 abc | 18.93 ± 7.47 bc | 19.74 ± 3.85 abc | 15.8 ± 2.14 c | 1.42 × 10−2 |
| OxFA | 15.09 ± 2.53 a | 16.33 ± 6.48 ab | 16.97 ± 0.86 ab | 20.64 ± 8.6 ab | 12.23 ± 4.65 b | 16.42 ± 8.67 ab | 9.95 ± 0.6 b | 1.16 × 10−1 |
| OxPC | 5.85 ± 0.63 a | 5.38 ± 0.84 ab | 4.95 ± 0.21 bc | 4.01 ± 0.52 d | 3.58 ± 0.47 d | 4.22 ± 0.9 c | 3.85 ± 0.46 d | 9.38 × 10−6 |
| OxPE | 12.63 ± 2.01 a | 13.18 ± 1.56 a | 12.92 ± 1.24 a | 9.51 ± 1.55 c | 9.59 ± 1.3 bc | 11.47 ± 1.85 ab | 9.04 ± 1.14 bc | 1.95 × 10−4 |
| OxTG | 4.29 ± 0.85 | 3.86 ± 0.24 | 4.51 ± 1.31 | 3.93 ± 1.76 | 3.45 ± 0.59 | 4.32 ± 2.27 | 3.57 ± 0.27 | 8.02 × 10−1 |
| PC | 43.96 ± 4.47 a | 35.01 ± 4.35 b | 34.54 ± 2.09 b | 32.41 ± 5.08 b | 30.96 ± 4.82 b | 30.13 ± 5.6 b | 32.53 ± 4.55 b | 1.04 × 10−3 |
| PE | 10.15 ± 0.54 a | 8.91 ± 1.05 ab | 8.62 ± 0.94 b | 8.03 ± 1.13 b | 8.46 ± 1.45 b | 8.11 ± 1.04 b | 7.9 ± 0.62 b | 2.39 × 10−2 |
| PG | 3.74 ± 0.33 b | 4.29 ± 0.71 b | 5.51 ± 0.2 a | 3.73 ± 0.71 b | 3.98 ± 1.09 b | 3.51 ± 0.63 b | 2.64 ± 0.38 c | 1.36 × 10−5 |
| PS | 3.05 ± 0.79 | 2.41 ± 0.94 | 3.3 ± 0.47 | 2.53 ± 0.76 | 3.74 ± 4.18 | 3.68 ± 1.46 | 1.96 ± 0.55 | 6.37 × 10−1 |
| SM | 18.59 ± 3.76 a | 16.66 ± 5.5 ab | 12.99 ± 1.2 bc | 12.56 ± 2.53 bc | 9.12 ± 2.84 c | 15.7 ± 4.57 ab | 15.6 ± 3.9 ab | 9.35 × 10−3 |
| TG | 34.85 ± 7.32 a | 21.85 ± 3.85 b | 40.4 ± 14.65 ab | 44.74 ± 20.45 ab | 38.37 ± 10.8 a | 42.31 ± 28.63 ab | 20.39 ± 3.24 b | 9.42 × 10−2 |
| VAE | 8.67 ± 0.5 abcd | 7.87 ± 1.46 cd | 8.22 ± 0.12 bc | 9.47 ± 1.16 abc | 9.84 ± 1.82 ab | 7.24 ± 0.74 d | 10.1 ± 1.91 a | 9.68 × 10−3 |
| Compounds | T1 | T2 | T3 | T4 | T5 | T6 | T7 | p Value |
|---|---|---|---|---|---|---|---|---|
| Hexanoic acid (C6:0) | 41.95 ± 12.10 b | 45.95 ± 8.53 b | 45.01 ± 9.66 b | 36.98 ± 10.53 b | 50.87 ± 15.91 b | 52.79 ± 18.52 b | 92.25 ± 22.24 a | 5.20 × 10−5 |
| Heptanoic acid (C7:0) | 8.49 ± 0.78 b | 6.47 ± 0.83 b | 8.78 ± 2.01 b | 9.61 ± 2.28 b | 9.46 ± 3.16 b | 8.41 ± 3.28 b | 16.00 ± 2.45 a | 2.77 × 10−5 |
| Octanoic acid (C8:0) | 12.14 ± 3.90 b | 12.94 ± 1.35 b | 19.66 ± 5.56 b | 16.97 ± 4.77 b | 18.72 ± 7.34 b | 20.82 ± 7.64 b | 35.23 ± 5.86 a | 1.04 × 10−5 |
| Nonanoic acid (C9:0) | 67.85 ± 8.54 b | 66.69 ± 8.96 b | 89.75 ± 12.48 ab | 90.74 ± 21.71 ab | 72.24 ± 14.49 b | 66.89 ± 22.09 b | 125.5 ± 40.09 a | 9.76 × 10−4 |
| Decanoic acid (C10:0) | 7.18 ± 1.19 b | 7.91 ± 2.77 b | 8.08 ± 1.21 b | 10.42 ± 2.40 ab | 7.37 ± 1.63 b | 8.40 ± 1.31 ab | 12.31 ± 2.47 a | 2.35 × 10−3 |
| Undecanoic acid (C11:0) | 2.24 ± 0.23 b | 2.47 ± 0.70 b | 2.05 ± 0.14 b | 2.31 ± 0.97 b | 2.06 ± 0.42 b | 3.02 ± 1.56 ab | 4.17 ± 0.86 a | 3.98 × 10−3 |
| Dodecanoic acid (C12:0) | 40.8 ± 3.21 c | 42.88 ± 5.67 c | 43.67 ± 8.91 c | 66.35 ± 5.15 ac | 46.98 ± 8.69 bc | 55.1 ± 12.27 abc | 60.44 ± 8.66 ab | 7.05 × 10−5 |
| Tridecanoic acid (C13:0) | 9.91 ± 1.44 a | 3.95 ± 0.32 c | 4.88 ± 0.69 bc | 3.80 ± 0.61 c | 2.75 ± 0.75 c | 4.60 ± 1.05 bc | 6.61 ± 1.83 b | 5.40 × 10−10 |
| Myristic acid (C14:0) | 215.64 ± 14.77 b | 190.81 ± 76.93 b | 190.73 ± 28.7 b | 251.99 ± 31.85 ab | 190.77 ± 31.12 b | 267.99 ± 49.63 ab | 328.75 ± 69.32 a | 4.46 × 10−4 |
| Pentadecanoic acid (C15:0) | 59.82 ± 7.48 a | 28.80 ± 3.94 c | 46.21 ± 6.7 b | 53.51 ± 6.58 ab | 41.49 ± 3.93 b | 46.32 ± 7.16 b | 61.44 ± 7.16 a | 6.19 × 10−8 |
| Palmitic acid (C16:0) | 2285.23 ± 151.17 cd | 2145.11 ± 427.41 d | 2288.36 ± 210.11 cd | 2702.32 ± 276.34 bc | 2276.38 ± 72.14 cd | 2904.48 ± 227.19 b | 3792.98 ± 283.38 a | 3.86 × 10−10 |
| cis-9-Hexadecenoic acid (C16:1) | 190.44 ± 12.6 cd | 178.76 ± 35.62 d | 190.7 ± 17.51 cd | 225.19 ± 23.03 bc | 189.7 ± 6.01 cd | 242.04 ± 18.93 b | 316.08 ± 23.61 a | 3.86 × 10−10 |
| Heptadecanoic acid (C17:0) | 25.74 ± 3.07 cd | 24.69 ± 1.47 d | 36.20 ± 3.18 bc | 38.16 ± 6.11 b | 36.26 ± 4.96 bc | 42.02 ± 8.06 b | 80.19 ± 8.56 a | 3.38 × 10−14 |
| Stearic acid (C18:0) | 1493.30 ± 134.63 c | 1404.80 ± 169.27 c | 1624.05 ± 35.98 bc | 1863.03 ± 217.15 b | 1509.55 ± 110.36 c | 1862.05 ± 151.18 b | 2321.65 ± 161.52 a | 1.54 × 10−9 |
| Elaidic acid (C18:1) | 64.31 ± 4.25 cd | 51.33 ± 10.78 d | 61.39 ± 11.00 cd | 77.7 ± 10.49 b | 63.13 ± 11.16 cd | 86.84 ± 14.30 b | 125.94 ± 10.44 a | 1.92 × 10−10 |
| cis-9-Octadecenoic acid (C18:1) | 990.02 ± 111.94 d | 1283.14 ± 216.31 cd | 1412.77 ± 180.39 bcd | 1623.19 ± 370.00 bc | 1654.44 ± 202.12 bc | 1837.89 ± 171.57 b | 2939.24 ± 414.91 a | 9.89 × 10−11 |
| Linoleic acid (C18:2) | 4651.50 ± 1994.05 cd | 4525.17 ± 516.76 d | 4937.29 ± 1073.83 cd | 5858.23 ± 1474.05 cd | 7379.44 ± 1988.49 bc | 8856.42 ± 951.04 ab | 10,323.84 ± 1177.47 a | 5.18 × 10−7 |
| α-Linolenic acid (C18:3) | 93.44 ± 31.32 c | 91.02 ± 14.83 c | 93.66 ± 16.33 c | 112.52 ± 20.26 bc | 128.92 ± 25.74 bc | 155.86 ± 2.97 ab | 191.37 ± 28.10 a | 1.21 × 10−7 |
| γ-Linolenic acid (C18:3) | 32.11 ± 14.81 c | 34.39 ± 6.47 c | 37.26 ± 5.30 c | 42.71 ± 11.99 c | 51.08 ± 11.39 bc | 70.27 ± 12.73 ab | 84.4 ± 15.18 a | 1.86 × 10−7 |
| Nonadecanoic acid (C19:0) | 17.94 ± 3.52 b | 14.87 ± 1.25 b | 21.68 ± 2.71 b | 21.68 ± 0.90 b | 22.73 ± 4.38 b | 21.73 ± 3.62 b | 43.12 ± 8.27 a | 8.89 × 10−10 |
| Arachidic acid (C20:0) | 13.21 ± 1.79 c | 10.38 ± 3.18 c | 14.28 ± 0.66 b | 19.86 ± 3.28 ab | 13.32 ± 4.12 c | 13.8 ± 2.15 c | 23.48 ± 3.34 a | 7.65 × 10−7 |
| cis-11-Eicosenoic acid (C20:1) | 49.34 ± 9.78 b | 59.72 ± 24.38 b | 58.7 ± 10.20 b | 69.85 ± 21.63 b | 70.46 ± 14.56 b | 80.04 ± 12.7 b | 120.94 ± 11.93 a | 3.09 × 10−6 |
| cis-11,14-Eicosenoic acid (C20:2) | 31.03 ± 2.94 d | 41.47 ± 5.01 cd | 46.85 ± 5.68 c | 54.14 ± 9.29 bc | 56.34 ± 6.47 bc | 64.19 ± 4.83 b | 94.20 ± 13.21 a | 5.73 × 10−12 |
| cis-8,11,14-Eicosenoic acid (C20:3) | 13.99 ± 4.18 b | 12.41 ± 2.72 b | 12.46 ± 2.96 b | 13.09 ± 1.43 b | 13.41 ± 1.14 b | 12.22 ± 2.68 b | 28.95 ± 5.19 a | 1.67 × 10−8 |
| cis-11,14,17-Eicosenoic acid (C20:3) | 180.19 ± 39.77 bcd | 148.48 ± 14.84 cd | 137.84 ± 6.01 d | 190.01 ± 43.23 bcd | 235.35 ± 70.14 bc | 265.19 ± 35.19 b | 378.32 ± 74.27 a | 7.58 × 10−8 |
| Arachidonic acid (C20:4) | 1432.37 ± 339.35 d | 1511.11 ± 154.33 d | 1515.61 ± 175.7 d | 1952.3 ± 383.42 cd | 2590.8 ± 522.5 bc | 3007.3 ± 235.77 ab | 3723.1 ± 820.65 a | 3.98 × 10−9 |
| cis-5,8,11,14,17-Dicosapentaenoic acid (C20:5) | 59.82 ± 13.49 d | 70.2 ± 8.95 d | 71.98 ± 16.64 d | 93.55 ± 27.53 cd | 135.13 ± 32.90 bc | 173.17 ± 30.49 ab | 217.17 ± 27.59 a | 4.16 × 10−11 |
| Heneicosanoic acid (C21:0) | 2.62 ± 1.41 bc | 0.68 ± 0.09 c | 1.17 ± 0.11 bc | 3.16 ± 2.47 bc | 3.88 ± 1.90 b | 1.06 ± 0.36 c | 7.66 ± 1.38 a | 1.42 × 10−7 |
| Behenic acid (C22:0) | 3.08 ± 1.14 c | 2.01 ± 0.47 c | 3.43 ± 0.35 bc | 3.63 ± 0.59 b | 3.26 ± 0.54 bc | 3.68 ± 0.60 b | 8.38 ± 1.16 a | 5.28 × 10−12 |
| cis-13-Docosenoic acid (C22:1) | 14.05 ± 3.84 b | 7.99 ± 2.67 b | 10.13 ± 2.62 b | 11.1 ± 4.91 b | 9.69 ± 3.75 b | 11.79 ± 1.98 b | 31.16 ± 4.34 a | 6.29 × 10−10 |
| cis-13,16-Docosadienoic acid (C22:2) | 3.26 ± 0.87 b | 5.95 ± 2.21 b | 4.16 ± 1.20 b | 3.75 ± 1.37 b | 5.72 ± 1.53 b | 5.33 ± 1.78 b | 12.72 ± 1.80 a | 5.23 × 10−9 |
| cis-13,16,19-Docosatrienoic acid (C22:3) | 6.15 ± 1.57 d | 11.03 ± 1.48 c | 11.12 ± 1.26 c | 8.69 ± 3.54 cd | 22.45 ± 2.75 a | 16.88 ± 1.50 b | 23.51 ± 2.63 a | 3.65 × 10−13 |
| cis-7,10,13,16-Docosatetraenoic acid (C22:4) | 120.77 ± 31.41 e | 175.54 ± 9.91 cde | 163.89 ± 10.47 de | 224.38 ± 46.00 bcd | 259.61 ± 39.24 abc | 332.34 ± 42.64 a | 279.61 ± 78.75 ab | 1.48 × 10−7 |
| cis-7,10,13,16,19-Docosatetraenoic acid (C22:5) | 23.36 ± 10.40 c | 22.46 ± 3.62 c | 24.21 ± 4.66 c | 32.45 ± 9.60 bc | 42.84 ± 15.37 bc | 50.74 ± 7.99 ab | 71.71 ± 17.40 a | 2.79 × 10−7 |
| cis-4,7,10,13,16,19-Docosatetraenoic acid (C22:6) | 90.5 ± 37.1 c | 75.54 ± 8.10 c | 88.9 ± 27.58 c | 111.24 ± 35.63 bc | 153.63 ± 17.95 ab | 175.62 ± 68.84 ab | 210.97 ± 56.63 a | 5.22 × 10−5 |
| Lignoceric acid (C24:0) | 15.49 ± 4.85 b | 7.49 ± 1.15 c | 12.31 ± 1.41 bc | 14.4 ± 3.19 b | 13.78 ± 3.78 bc | 16.35 ± 3.99 b | 28.09 ± 3.45 a | 2.55 × 10−8 |
| cis-15-Tetracosenoic acid (C24:1) | 7.13 ± 1.20 c | 8.15 ± 0.80 c | 9.71 ± 1.57 bc | 9.34 ± 2.00 bc | 12.21 ± 2.58 b | 12.7 ± 2.01 b | 25.21 ± 1.71 a | 1.21 × 10−14 |
| Hexacosanoic acid (C26:0) | 14.3 ± 3.04 b | 7.11 ± 1.55 b | 10.05 ± 1.56 b | 12.19 ± 3.03 b | 9.92 ± 2.55 b | 13.28 ± 5.57 b | 28.91 ± 6.96 a | 5.52 × 10−8 |
| Octacosanoic acid (C28:0) | 3.69 ± 1.24 b | 1.68 ± 0.25 b | 3.11 ± 0.52 b | 2.59 ± 0.81 b | 2.94 ± 2.20 b | 3.74 ± 0.98 b | 16.83 ± 4.61 a | 1.79 × 10−11 |
| Melissic acid (C30:0) | 6.22 ± 3.50 b | 1.41 ± 0.19 b | 3.64 ± 1.50 b | 1.62 ± 0.42 b | 2.78 ± 2.71 b | 1.55 ± 1.07 b | 22.54 ± 4.63 a | 3.58 × 10−13 |
| SFA | 4346.85 ± 296.53 d | 4029.1 ± 605.20 d | 4477.09 ± 294.29 cd | 5225.32 ± 378.14 bc | 4337.51 ± 187.17 d | 5418.07 ± 476.1 b | 7116.53 ± 554.60 a | 3.18 × 10−11 |
| PUFA | 6738.5 ± 2465.51 d | 6724.76 ± 609.61 d | 7145.23 ± 1262.16 d | 8697.06 ± 1909.39 cd | 11,074.72 ± 2646.61 bc | 13,185.52 ± 1300.82 ab | 15,639.87 ± 1737.61 a | 1.43 × 10−8 |
| MUFA | 3410.08 ± 193.36 d | 3555.44 ± 640.99 d | 3841.04 ± 338.74 cd | 4493.5 ± 217.81 bc | 4086.3 ± 230.95 bcd | 4933.73 ± 393.69 b | 7035.46 ± 688.71 a | 1.00 × 10−12 |
| UFA | 10,148.57 ± 2540.65 d | 10,280.20 ± 1057.40 d | 10,986.27 ± 1300.68 d | 13,190.56 ± 1969.59 cd | 15,161.02 ± 2644.33 bc | 18,119.25 ± 1052.51 b | 22,675.33 ± 1517.95 a | 2.30 × 10−11 |
| Total FA | 14,495.42 ± 2602.75 d | 14,309.30 ± 1652.90 d | 15,463.37 ± 1512.93 d | 18,415.88 ± 2090.33 cd | 19,498.54 ± 2778.78 c | 23,537.32 ± 923.82 b | 29,791.87 ± 1297.66 a | 9.07 × 10−13 |
| ω3/ω6 | 0.08 ± 0.02 | 0.07 ± 0.00 | 0.06 ± 0.00 | 0.07 ± 0.00 | 0.07 ± 0.01 | 0.07 ± 0.00 | 0.08 ± 0.00 | 1.95 × 10−1 |
| ω7/ω6 | 0.42 ± 0.02 | 0.34 ± 0.06 | 0.35 ± 0.05 | 0.34 ± 0.07 | 0.23 ± 0.05 | 0.24 ± 0.04 | 0.26 ± 0.05 | 2.23 × 10−2 |
| ω9/ω6 | 0.21 ± 0.12 | 0.22 ± 0.04 | 0.24 ± 0.07 | 0.23 ± 0.08 | 0.19 ± 0.07 | 0.17 ± 0.03 | 0.23 ± 0.05 | 6.87 × 10−1 |
| Compounds | T1 | T2 | T3 | T4 | T5 | T6 | T7 | p Value |
|---|---|---|---|---|---|---|---|---|
| Propanal | 49.69 ± 1.22 c | 91.8 ± 3.30 c | 192.49 ± 9.21 c | 247.52 ± 7.98 b | 253.28 ± 1.23 b | 280.91 ± 5.56 a | 277.19 ± 4.13 a | 2.85 × 10−30 |
| Butanal | 0.59 ± 0.01 d | 0.64 ± 0.02 d | 1.18 ± 0.04 c | 1.50 ± 0.10 b | 1.63 ± 0.01 a | 1.68 ± 0.04 a | 1.74 ± 0.03 a | 2.74 × 10−24 |
| Pentanal | 2.37 ± 0.24 bc | 2.62 ± 0.27 bc | 2.09 ± 0.41 c | 2.38 ± 0.16 bc | 3.04 ± 0.05 b | 4.89 ± 0.21 a | 5.18 ± 0.67 a | 1.73 × 10−13 |
| Hexanal | 7.01 ± 0.26 e | 12.49 ± 0.15 d | 27.86 ± 1.29 c | 31.72 ± 1.17 b | 32.27 ± 0.41 b | 40.73 ± 1.20 a | 39.89 ± 0.49 a | 1.57 × 10−23 |
| Heptanal | 0.78 ± 0.06 c | 0.86 ± 0.11 c | 0.71 ± 0.08 c | 0.83 ± 0.06 c | 1.25 ± 0.01 b | 1.57 ± 0.07 a | 1.49 ± 0.20 ab | 8.92 × 10−13 |
| Octanal | 1.28 ± 0.05 b | 1.36 ± 0.05 b | 1.22 ± 0.1 b | 1.41 ± 0.24 b | 2.4 ± 0.02 a | 2.34 ± 0.14 a | 2.55 ± 0.25 a | 1.15 × 10−14 |
| Nonanal | 4.73 ± 0.75 b | 4.48 ± 0.32 b | 4.02 ± 0.49 b | 6.27 ± 0.55 a | 7.19 ± 0.07 a | 6.13 ± 0.33 a | 7.28 ± 0.76 a | 9.60 × 10−9 |
| Decanal | 3.37 ± 0.43 d | 3.17 ± 0.10 d | 3.29 ± 1.09 d | 6.02 ± 1.06 c | 7.16 ± 0.02 bc | 8.41 ± 0.52 ab | 9.00 ± 0.34 a | 4.80 × 10−16 |
| Undecanal | 0.59 ± 0.21 a | 0.38 ± 0.08 a | 0.57 ± 0.12 a | 1.04 ± 0.16 b | 1.13 ± 0.01 b | 1.16 ± 0.09 b | 1.34 ± 0.19 b | 1.55 × 10−11 |
| Propenal | 1.35 ± 0.29 b | 1.88 ± 0.16 a | 1.16 ± 0.05 b | 0.95 ± 0.11 bc | 0.67 ± 0.04 cd | 0.45 ± 0.08 d | 0.7 ± 0.05 cd | 1.34 × 10−10 |
| Crotonaldehyde | 2.59 ± 0.08 e | 4.56 ± 0.22 d | 7.65 ± 0.31 c | 7.86 ± 0.47 c | 10.41 ± 0.02 a | 8.66 ± 0.18 b | 8.13 ± 0.20 bc | 8.71 × 10−27 |
| 2-Pentenal | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.04 ± 0.00 | 0.06 ± 0.00 | 0.08 ± 0.00 | 0.07 ± 0.00 | 0.07 ± 0.00 | 8.30 × 10−14 |
| 2-Hexanal | 0.5 ± 0.03 e | 0.66 ± 0.02 e | 1.07 ± 0.05 d | 1.53 ± 0.12 b | 2.55 ± 0.02 a | 0.64 ± 0.01 e | 1.28 ± 0.05 c | 3.28 × 10−23 |
| 2,4-Heltadienal | 0.03 ± 0.01 c | 0.16 ± 0.00 b | 0.10 ± 0.01 b | 0.05 ± 0.00 b | 0.12 ± 0.00 b | 0.03 ± 0.00 c | 0.06 ± 0.00 b | 2.63 × 10−14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Li, J.; Zhang, L.; Rao, Z.; Feng, S.; Wang, Y.; Liu, H.; Meng, Q. Integrated Lipidomic and Metabolomics Analysis Revealing the Effects of Frozen Storage Duration on Pork Lipids. Metabolites 2022, 12, 977. https://doi.org/10.3390/metabo12100977
Feng X, Li J, Zhang L, Rao Z, Feng S, Wang Y, Liu H, Meng Q. Integrated Lipidomic and Metabolomics Analysis Revealing the Effects of Frozen Storage Duration on Pork Lipids. Metabolites. 2022; 12(10):977. https://doi.org/10.3390/metabo12100977
Chicago/Turabian StyleFeng, Xiaohui, Jing Li, Longchao Zhang, Zhenghua Rao, Shengnan Feng, Yujiao Wang, Hai Liu, and Qingshi Meng. 2022. "Integrated Lipidomic and Metabolomics Analysis Revealing the Effects of Frozen Storage Duration on Pork Lipids" Metabolites 12, no. 10: 977. https://doi.org/10.3390/metabo12100977
APA StyleFeng, X., Li, J., Zhang, L., Rao, Z., Feng, S., Wang, Y., Liu, H., & Meng, Q. (2022). Integrated Lipidomic and Metabolomics Analysis Revealing the Effects of Frozen Storage Duration on Pork Lipids. Metabolites, 12(10), 977. https://doi.org/10.3390/metabo12100977