
Integrated Analysis of Widely Targeted Metabolomics and Transcriptomics Reveals the Effects of Transcription Factor NOR-like1 on Alkaloids, Phenolic Acids, and Flavonoids in Tomato at Different Ripening Stages

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sample Preparation
2.2. Widely Targeted Metabolic Analysis
2.3. Differential-Metabolite (DEM) Analysis
2.4. Transcriptomic Analysis
2.5. Quantification of Gene-Expression Levels
2.6. Differential Analysis and Differential Gene-Enrichment Analysis
3. Results
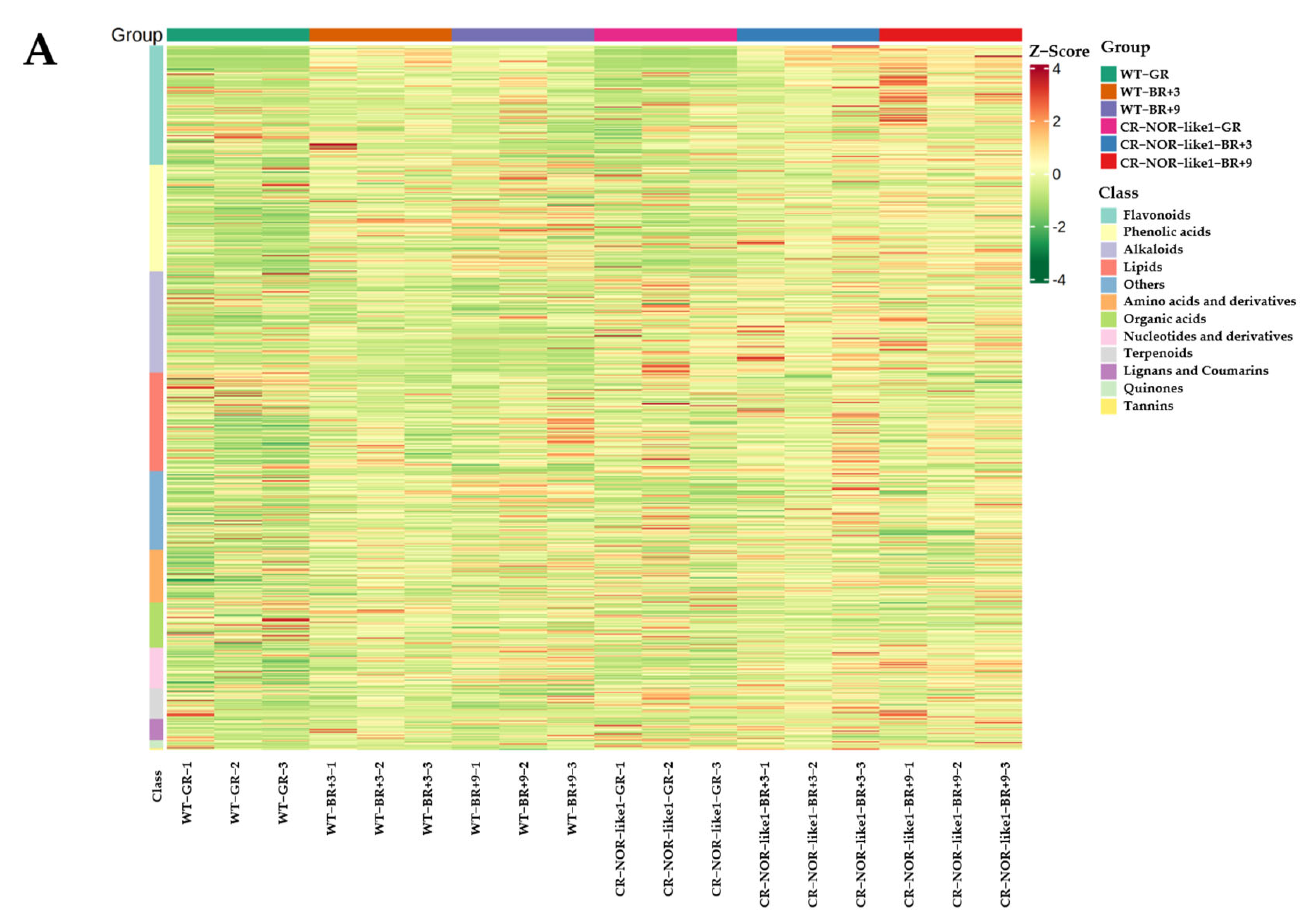
3.1. Widely Targeted Metabolomic Differential Analysis
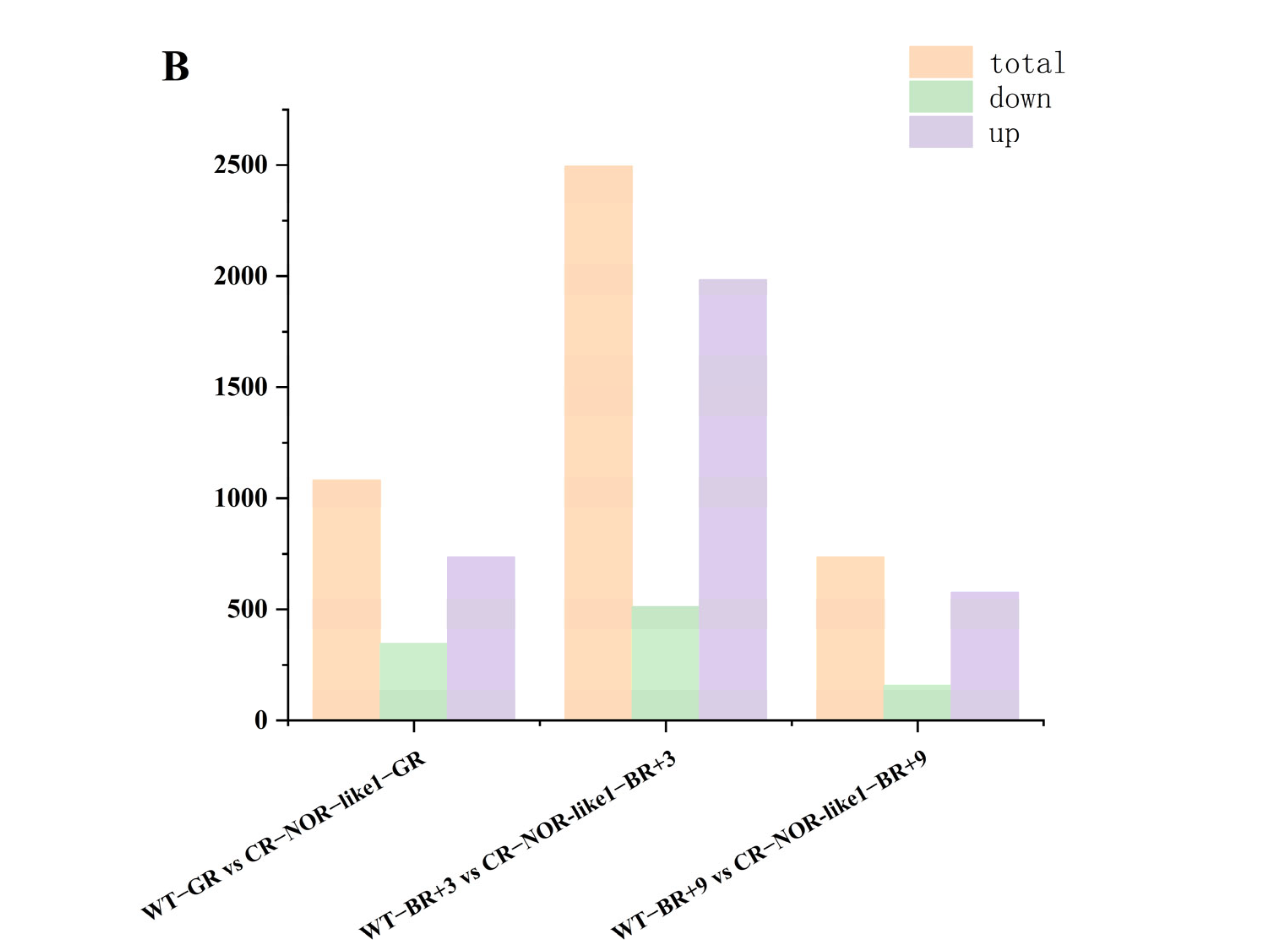
3.2. Differential-Metabolite (DEM) Identification
3.3. An Overview of RNA-Seq Data
3.4. Differentially Expressed Gene (DEG) Identification
3.5. Effect of NOR-like1 on Alkaloids
3.6. Effect of NOR-like1 on Phenolic Acids
3.7. Effect of NOR-like1 on Flavonoids
3.8. Correlation Network Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Mun, H.I.; Kwon, M.C.; Lee, N.R.; Son, S.Y.; Song, D.H.; Lee, C.H. Comparing Metabolites and Functional Properties of Various Tomatoes Using Mass Spectrometry-Based Metabolomics Approach. Front. Nutr. 2021, 8, 659646. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.H.P.; Kupski, L.; de Souza, T.D.; Arias, J.L.D.O.; D’Oca, M.M.; Furlong, E.B. Relations between nutrients and bioactive compounds of commercial tomato varieties by the Principal Component Analysis. Food Sci. Technol. 2022, 42, 60020. [Google Scholar] [CrossRef]
- Min, D.; Li, Z.; Fu, X.; Wang, J.; Li, F.; Li, X.; Zhang, X. Integration of transcriptomic and metabonomic reveals molecular differences of sweetness and aroma between postharvest and vine ripened tomato fruit. Food Control 2022, 139, 109102. [Google Scholar] [CrossRef]
- Bailly, C. The steroidal alkaloids alpha-tomatine and tomatidine: Panorama of their mode of action and pharmacological properties. Steroids 2021, 176, 108933. [Google Scholar] [CrossRef] [PubMed]
- Dzakovich, M.P.; Hartman, J.L.; Cooperstone, J.L. A High-Throughput Extraction and Analysis Method for Steroidal Glycoalkaloids in Tomato. Front. Plant Sci. 2020, 11, 767. [Google Scholar] [CrossRef]
- Nakayasu, M.; Akiyama, R.; Kobayashi, M.; Lee, H.J.; Kawasaki, T.; Watanabe, B.; Urakawa, S.; Kato, J.; Sugimoto, Y.; Iijima, Y.; et al. Identification of alpha-Tomatine 23-Hydroxylase Involved in the Detoxincation of a Bitter Glycoalkaloid. Plant Cell Physiol. 2020, 61, 21–28. [Google Scholar] [CrossRef]
- Pardini, A.; Consumi, M.; Leone, G.; Bonechi, C.; Tamasi, G.; Sangiorgio, P.; Verardi, A.; Rossi, C.; Magnani, A. Effect of different post-harvest storage conditions and heat treatment on tomatine content in commercial varieties of green tomatoes. J. Food Compos. Anal. 2021, 96, 103735. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of postharvest UV-C irradiation on phenolic acids, flavonoids, and key phenylpropanoid pathway genes in tomato fruit. Sci. Hortic. 2018, 241, 107–114. [Google Scholar] [CrossRef]
- Slimestad, R.; Verheul, M. Review of flavonoids and other phenolics from fruits of different tomato (Lycopersicon esculentum Mill.) cultivars. J. Sci. Food Agric. 2009, 89, 1255–1270. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialecka-Florjanczyk, E.; Fabiszewska, A.; Zieniuk, B. Phenolic Acids Derivatives—Biotechnological Methods of Synthesis and Bioactivity. Curr. Pharm. Biotechnol. 2018, 19, 1098–1113. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S. Tomato Fruit Development and Metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [Green Version]
- Ballester, A.-R.; Tikunov, Y.; Molthoff, J.; Grandillo, S.; Viquez-Zamora, M.; de Vos, R.; de Maagd, R.A.; van Heusden, S.; Bovy, A.G. Identification of Loci Affecting Accumulation of Secondary Metabolites in Tomato Fruit of a Solanum lycopersicum × Solanum chmielewskii Introgression Line Population. Front. Plant Sci. 2016, 7, 1428. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Xu, X.; Hu, X.; Liu, Y.; Cao, H.; Chan, H.; Gong, Z.; Yuan, Y.; Luo, Y.; Feng, B.; et al. SlMYB72 Regulates the Metabolism of Chlorophylls, Carotenoids, and Flavonoids in Tomato Fruit. Plant Physiol. 2020, 183, 854–868. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Cao, H.; Yuan, S.; Liu, Y.; Lu, J.; Lu, W.; Li, N.; Wang, J.; Zou, J.; Tang, N.; et al. SlMYB75, an MYB-type transcription factor, promotes anthocyanin accumulation and enhances volatile aroma production in tomato fruits. Hortic. Res. 2019, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Koul, A.; Kaul, S.; Dhar, M.K. Tissue-specific transcriptional regulation and metabolite accumulation in tomato (Solanum lycopersicum L.). Protoplasma 2020, 257, 1093–1108. [Google Scholar] [CrossRef]
- Bineau, E.; Rambla, J.L.; Duboscq, R.; Corre, M.N.; Bitton, F.; Lugan, R.; Granell, A.; Plissonneau, C.; Causse, M. Inheritance of Secondary Metabolites and Gene Expression Related to Tomato Fruit Quality. Int. J. Mol. Sci. 2022, 23, 6163. [Google Scholar] [CrossRef]
- Sacco, A.; Raiola, A.; Calafiore, R.; Barone, A.; Rigano, M.M. New insights in the control of antioxidants accumulation in tomato by transcriptomic analyses of genotypes exhibiting contrasting levels of fruit metabolites. BMC Genom. 2019, 20, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Fan, Z.-Q.; Zhang, Q.; Li, H.-L.; Liu, G.-S.; Jing, Y.; Zhang, Y.-P.; Zhu, B.-Z.; Zhu, H.-L.; Chen, J.-Y.; et al. A tomato NAC transcription factor, SlNAM1, positively regulates ethylene biosynthesis and the onset of tomato fruit ripening. Plant J. 2021, 108, 1317–1331. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hou, X.; Qi, N.; Liu, H.; Li, Y.; Huang, D.; Wang, C.; Liao, W. Insight into ripening-associated transcription factors in tomato: A review. Sci. Hortic. 2021, 288, 110363. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, W.; Zhao, X.; Tan, X.; Fan, Z.; Zhang, Y.; Jing, Y.; Meng, L.; Zhu, B.; Zhu, H.; et al. A NAC transcription factor, NOR-like1, is a new positive regulator of tomato fruit ripening. Hortic. Res. 2018, 5, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wei, W.; Fan, Z.; Zhao, X.; Zhang, Y.; Jing, Y.; Zhu, B.; Zhu, H.; Shan, W.; Chen, J.; et al. Re-evaluation of the nor mutation and the role of the NAC-NOR transcription factor in tomato fruit ripening. J. Exp. Bot. 2020, 71, 3560–3574. [Google Scholar] [CrossRef]
- Brumos, J. Gene regulation in climacteric fruit ripening. Curr. Opin. Plant Biol. 2021, 63, 102042. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, N.; Zhu, X.; Wu, M.; Jiang, C.-Z.; Grierson, D.; Luo, Y.; Shen, W.; Zhong, S.; Fu, D.-Q.; et al. Diversity and redundancy of the ripening regulatory networks revealed by the fruitENCODE and the new CRISPR/Cas9 CNR and NOR mutants. Hortic. Res. 2019, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Feng, H.; Meng, X.; Li, D.; Yang, D.; Wu, C.; Meng, Q. Overexpression of tomato SlNAC1 transcription factor alters fruit pigmentation and softening. BMC Plant Biol. 2014, 14, 351. [Google Scholar] [CrossRef] [Green Version]
- Meng, C.; Yang, D.; Ma, X.; Zhao, W.; Liang, X.; Ma, N.; Meng, Q. Suppression of tomato SlNAC1 transcription factor delays fruit ripening. J. Plant Physiol. 2016, 193, 88–96. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A new tomato NAC (NAM/ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2014, 55, 119–135. [Google Scholar] [CrossRef]
- Kou, X.; Liu, C.; Han, L.; Wang, S.; Xue, Z. NAC transcription factors play an important role in ethylene biosynthesis, reception and signaling of tomato fruit ripening. Mol. Genet. Genom. 2016, 291, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Huang, S.; Zhang, Q.; Zhu, S.; Dong, X. Changes in the Primary Metabolites of ‘Fengtang’ Plums during Storage Detected by Widely Targeted Metabolomics. Foods 2022, 11, 2830. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Qin, Y.; Zhong, C.; Zhou, R.; Xie, J.; Liu, H.; Xiao, J.; Cai, P.; Zhang, S.; Qin, Y. Differential gene expression and associated metabolite accumulation in fungus Ophiocordyceps xuefengensis cultivated under breathable and airtight conditions. Mycoscience 2019, 60, 281–286. [Google Scholar] [CrossRef]
- Leng, F.; Cao, J.; Ge, Z.; Wang, Y.; Zhao, C.; Wang, S.; Li, X.; Zhang, Y.; Sun, C. Transcriptomic Analysis of Root Restriction Effects on Phenolic Metabolites during Grape Berry Development and Ripening. J. Agric. Food Chem. 2020, 68, 9090–9099. [Google Scholar] [CrossRef]
- Kim, J.I.; Hidalgo-Shrestha, C.; Bonawitz, N.D.; Franke, R.B.; Chapple, C. Spatio-temporal control of phenylpropanoid biosynthesis by inducible complementation of a cinnamate 4-hydroxylase mutant. J. Exp. Bot. 2021, 72, 3061–3073. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Cao, H.; Hao, X.; Zeng, J.; Yang, Y.; Wang, X. Transcriptome Analysis of an Anthracnose-Resistant Tea Plant Cultivar Reveals Genes Associated with Resistance to Colletotrichum camelliae. PLoS ONE 2016, 11, e0148535. [Google Scholar] [CrossRef]
- Li, X.; Ma, D.; Chen, J.; Pu, G.; Ji, Y.; Lei, C.; Du, Z.; Liu, B.; Ye, H.; Wang, H. Biochemical characterization and identification of a cinnamyl alcohol dehydrogenase from Artemisia annua. Plant Sci. 2012, 193, 85–95. [Google Scholar] [CrossRef]
- Chaudhary, A.; Jaswal, V.S.; Choudhary, S.; Sonika; Sharma, A.; Beniwal, V.; Tuli, H.S.; Sharma, S. Ferulic Acid: A Promising Therapeutic Phytochemical and Recent Patents Advances. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 115–123. [Google Scholar] [CrossRef]
- Otero, C.; Miranda-Rojas, S.; Llancalahuen, F.M.; Fuentes, J.A.; Atala, C.; Gonzalez-Silva, G.; Verdugo, D.; Sierra-Rosales, P.; Moreno, A.; Gordillo-Fuenzalida, F. Biochemical characterization of Peumus boldus fruits: Insights of its antioxidant properties through a theoretical approach. Food Chem. 2022, 370, 131012. [Google Scholar] [CrossRef]
- Yang, M.; Zhou, P.; Gui, C.; Da, G.; Gong, L.; Zhang, X. Comparative Transcriptome Analysis of Ampelopsis megalophylla for Identifying Genes Involved in Flavonoid Biosynthesis and Accumulation during Different Seasons. Molecules 2019, 24, 1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.; Shen, C.; Feng, X.; Gao, M.; Zhang, Y.; Wei, X.; Chen, Y.; Cheng, C.; Wu, R. Integrated Metabolomic and Transcriptomic Analysis Reveals Differential Flavonoid Accumulation and Its Underlying Mechanism in Fruits of Distinct Canarium album Cultivars. Foods 2022, 11, 2527. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, L.; Cai, Z.; Chen, C.; Liu, Z.; Liu, S.; Zou, L.; Tan, M.; Chen, J.; Liu, X.; et al. Metabolite Profiling and Transcriptome Analysis Explains Difference in Accumulation of Bioactive Constituents in Licorice (Glycyrrhiza uralensis) Under Salt Stress. Front. Plant Sci. 2021, 12, 727882. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zheng, W.; Wang, Y.; Wu, Y.; Yu, J.; Xia, P. Integrative Metabolome and Transcriptome Analysis Reveals the Regulatory Network of Flavonoid Biosynthesis in Response to MeJA in Camellia vietnamensis Huang. Int. J. Mol. Sci. 2022, 23, 9370. [Google Scholar] [CrossRef]
- Zhang, X.; Abrahan, C.; Colquhoun, T.A.; Liu, C.-J. A Proteolytic Regulator Controlling Chalcone Synthase Stability and Flavonoid Biosynthesis in Arabidopsis. Plant Cell 2017, 29, 1157–1174. [Google Scholar] [CrossRef] [Green Version]
- Verhoeyen, M.E.; Bovy, A.; Collins, G.; Muir, S.; Robinson, S.; de Vos, C.H.R.; Colliver, S. Increasing antioxidant levels in tomatoes through modification of the flavonoid biosynthetic pathway. J. Exp. Bot. 2002, 53, 2099–2106. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.-c.; Zhang, X.-d.; Gao, Z.-q.; Hu, T.; Liu, Y. The Research Progress of Chalcone Isomerase (CHI) in Plants. Mol. Biotechnol. 2019, 61, 32–52. [Google Scholar] [CrossRef]
- Zhang, Y.; de Stefano, R.; Robine, M.; Butelli, E.; Bulling, K.; Hill, L.; Rejzek, M.; Martin, C.; Schoonbeek, H.-j. Different ROS-Scavenging Properties of Flavonoids Determine Their Abilities to Extend Shelf Life of Tomato. Plant Physiol. 2015, 169, 1568–1583. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | WT-GR vs. CR-NOR-like1-GR | WT-BR+3 vs. CR-NOR-like1-BR+3 | WT-BR+9 vs. CR-NOR-like1-BR+9 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Log2FC | p-Value | Type | Log2FC | p-Value | Type | Log2FC | p-Value | Type | ||
| Aldehyde dehydrogenase (ALDH) | LOC104645747 | −1.03 | 3.59 × 10−3 | Down | 2.50 | 9.62 × 10−13 | Up | 2.58 | 1.38 × 10−4 | Up |
| SlADH2B7d | −1.64 | 2.12 × 10−26 | Down | -- | -- | -- | -- | -- | -- | |
| LOC101250474 | -- | -- | -- | −2.10 | 8.19 × 10−10 | Down | −1.77 | 5.66 × 10−6 | Down | |
| SlADH2B7d | -- | -- | -- | 2.35 | 5.23 × 10−13 | Up | -- | -- | -- | |
| SlADH3F1a | -- | -- | -- | −1.57 | 1.62 × 10−13 | Down | -- | -- | -- | |
| SlALDH2B7a | -- | -- | -- | 3.03 | 4.59 × 10−14 | Up | 1.45 | 5.86 × 10−6 | Up | |
| SlALD3H1 | -- | -- | -- | -- | -- | -- | −1.11 | 4.09 × 10−7 | Down | |
| Amidase (Ami) | LOC101257218 | −1.57 | 5.71 × 10−4 | Down | -- | -- | -- | -- | -- | -- |
| LOC101260379 | 1.97 | 1.72 × 10−3 | Up | -- | -- | -- | -- | -- | -- | |
| Arginase (ARG2) | ARG2 | 2.27 | 4.39 × 10−4 | Up | 3.75 | 1.25 × 10−3 | Up | -- | -- | -- |
| Ornithine decarboxylase (ODC) | ODC | 1.90 | 1.19 × 10−3 | Up | -- | -- | -- | -- | -- | -- |
| Arginine decarboxylase (adc1) | adc1 | 1.98 | 5.37 × 10−6 | Up | -- | -- | -- | -- | -- | -- |
| N-carbamoylputrescine amidase (CPA) | LOC101268110 | 9.02 | 1.53 × 10−12 | Up | 9.39 | 4.97 × 10−14 | Up | 7.65 | 1.61 × 10−9 | Up |
| Delta-1-pyrroline-5-carboxylate synthetase (P5CS) | LOC101244293 | −1.16 | 2.50 × 10−10 | Down | -- | -- | -- | -- | -- | -- |
| Proline dehydrogenase (PDH) | PDH | -- | -- | -- | 2.33 | 8.58 × 10−15 | Up | -- | -- | -- |
| Aspartate aminotransferase (AST) | LOC101244012 | -- | -- | -- | -- | -- | -- | −1.15 | 4.13 × 10−6 | Down |
| Gene Name | Gene ID | WT-GR vs. CR-NOR-like1-GR | WT-BR+3 vs. CR-NOR-like1-BR+3 | WT-BR+9 vs. CR-NOR-like1-BR+9 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Log2FC | p-Value | Type | Log2FC | p-Value | Type | Log2FC | p-Value | Type | ||
| Phenylalanine ammonia-lyase (PAL) | PAL5 | 2.71 | 2.75 × 10−4 | Up | -- | -- | -- | 2.52 | 1.50 × 10−6 | Up |
| LOC101243631 | -- | -- | -- | -- | -- | -- | 1.18 | 1.62 × 10−4 | Up | |
| PAL3 | -- | -- | -- | -- | -- | -- | 1.27 | 2.20 × 10−6 | Up | |
| Trans-cinnamate 4-monooxygenase (C4H) | LOC101262919 | 3.28 | 2.83 × 10−6 | Up | -- | -- | -- | −2.45 | 1.56 × 10−6 | Down |
| LOC101244496 | -- | -- | -- | -- | -- | -- | 1.15 | 1.52 × 10−3 | Up | |
| 4-coumarate—CoA ligase (4CL) | LOC101251197 | −4.24 | 1.46 × 10−4 | Down | -- | -- | -- | -- | -- | -- |
| LOC101251363 | -- | -- | -- | 1.06 | 1.75 × 10−3 | Up | 1.71 | 1.65 × 10−3 | Up | |
| Cinnamoyl-CoA reductase (CCR) | CCR2 | −1.86 | 1.39 × 10−6 | Down | -- | -- | -- | -- | -- | -- |
| LOC101246651 | −1.59 | 1.55 × 10−5 | Down | 1.47 | 6.13 × 10−5 | Up | 2.11 | 4.65 × 10−5 | Up | |
| LOC101264879 | −2.04 | 1.23 × 10−4 | Down | -- | -- | -- | 1.71 | 2.82 × 10−6 | Up | |
| LOC101250958 | -- | -- | -- | 1.16 | 3.84 × 10−3 | Up | -- | -- | -- | |
| LOC101262601 | -- | -- | -- | 2.71 | 8.51 × 10−14 | Up | -- | -- | -- | |
| LOC101265652 | -- | -- | -- | 1.32 | 9.78 × 10−5 | Up | -- | -- | -- | |
| PAR2 | -- | -- | -- | 1.17 | 2.15 × 10−3 | Up | -- | -- | -- | |
| Cinnamyl-alcohol dehydrogenase (CAD) | LOC112940682 | 6.67 | 7.36 × 10−5 | Up | -- | -- | -- | -- | -- | -- |
| LOC101253340 | −3.12 | 1.84 × 10−3 | Down | 2.51 | 4.30 × 10−3 | Up | -- | -- | -- | |
| LOC101250635 | -- | -- | -- | 1.45 | 4.30 × 10−4 | Up | -- | -- | -- | |
| LOC101265606 | -- | -- | -- | -- | -- | -- | −2.27 | 3.28 × 10−3 | Down | |
| Peroxidase (POD) | LOC101244376 | −7.59 | 8.55 × 10−7 | Down | -- | -- | -- | -- | -- | -- |
| LOC101251503 | 5.18 | 4.85 × 10−13 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101253377 | 1.34 | 2.53 × 10−3 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101257228 | 4.96 | 5.63 × 10−11 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101263035 | −1.25 | 3.36 × 10−3 | Down | -- | -- | -- | -- | -- | -- | |
| LOC101258529 | -- | -- | -- | 2.72 | 1.18 × 10−3 | Up | -- | -- | -- | |
| LOC101267754 | -- | -- | -- | 2.18 | 3.45 × 10−3 | Up | -- | -- | -- | |
| LOC101268153 | -- | -- | -- | 2.58 | 4.52 × 10−4 | Up | -- | -- | -- | |
| TAP2 | -- | -- | -- | 1.82 | 2.65 × 10−3 | Up | -- | -- | -- | |
| TMP1 | -- | -- | -- | 2.76 | 4.28 × 10−18 | Up | -- | -- | -- | |
| LOC101253648 | -- | -- | -- | -- | -- | -- | −1.45 | 6.78 × 10−5 | Down | |
| Caffeoyl-CoA O-methyltransferase (CCoAOMT) | LOC101253032 | 2.12 | 8.52 × 10−10 | Up | -- | -- | -- | -- | -- | -- |
| LOC101252203 | -- | -- | -- | 2.70 | 1.47 × 10−6 | Up | -- | -- | -- | |
| Beta-glucosidase (BGL) | LOC101249847 | 3.83 | 4.29 × 10−7 | Up | -- | -- | -- | -- | -- | -- |
| LOC101246223 | −2.81 | 2.96 × 10−4 | Down | -- | -- | -- | -- | -- | -- | |
| LOC101251735 | -- | -- | -- | −1.51 | 2.27 × 10−5 | Down | -- | -- | -- | |
| LOC101256510 | -- | -- | -- | 1.66 | 3.21 × 10−4 | Up | -- | -- | -- | |
| LOC101256717 | -- | -- | -- | 1.37 | 1.00 × 10−4 | Up | -- | -- | -- | |
| LOC101265077 | -- | -- | -- | 1.24 | 2.71 × 10−8 | Up | -- | -- | -- | |
| LOC101260057 | -- | -- | -- | 2.59 | 1.60 × 10−6 | Up | -- | -- | -- | |
| LOC101266643 | -- | -- | -- | 2.57 | 1.65 × 10−6 | Up | -- | -- | -- | |
| Feruloyl-CoA ortho-hydroxylase (FC2′H) | LOC101252918 | 2.75 | 8.03 × 10−6 | Up | 2.05 | 1.39 × 10−3 | Up | -- | -- | -- |
| Coumaroylquinate(coumaroylshikimate) 3′-monooxygenase (C3′H) | LOC101246092 | 1.77 | 3.55 × 10−3 | Up | 1.17 | 3.53 × 10−3 | Up | -- | -- | -- |
| Coniferyl-aldehyde dehydrogenase (REF1) | LOC101247788 | 1.27 | 1.63 × 10−3 | Up | -- | -- | -- | -- | -- | -- |
| Shikimate O-hydroxycinnamoyltransferase (HCT) | LOC101245886 | −2.20 | 3.09 × 10−6 | Down | 2.30 | 3.42 × 10−10 | Up | 2.18 | 3.96 × 10−5 | Up |
| LOC101247305 | 7.39 | 1.06 × 10−4 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101248087 | 5.05 | 3.14 × 10−8 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101252161 | −2.31 | 2.14 × 10−3 | Down | -- | -- | -- | 2.28 | 7.77 × 10−4 | Up | |
| LOC101253556 | 5.90 | 5.02 × 10−12 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101256271 | 1.60 | 1.93 × 10−3 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101260610 | 5.16 | 1.78 × 10−4 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101244961 | -- | -- | -- | −1.58 | 1.04 × 10−3 | Down | -- | -- | -- | |
| LOC101246106 | -- | -- | -- | −1.24 | 3.17 × 10−3 | Down | -- | -- | -- | |
| LOC101266953 | -- | -- | -- | 1.62 | 5.32 × 10−3 | Up | -- | -- | -- | |
| Caffeic acid 3-O-methyltransferase (COMT) | LOC101251452 | 1.49 | 5.89 × 10−4 | Up | -- | -- | -- | -- | -- | -- |
| Scopoletin glucosyltransferase (TOGT1) | LOC101253350 | 3.07 | 6.84 × 10−8 | Up | -- | -- | -- | -- | -- | -- |
| LOC101259704 | −1.92 | 1.37 × 10−16 | Down | -- | -- | -- | -- | -- | -- | |
| twi1 | 1.31 | 2.54 × 10−3 | Up | -- | -- | -- | -- | -- | -- | |
| GAME1 | -- | -- | -- | 2.32 | 1.34 × 10−2 | Up | -- | -- | -- | |
| LOC101258702 | -- | -- | -- | −1.36 | 3.72 × 10−4 | Down | -- | -- | -- | |
| LOC101260915 | -- | -- | -- | 5.63 | 1.64 × 10−10 | Up | -- | -- | -- | |
| Coniferyl-alcohol glucosyltransferase (CAGT) | LOC101256157 | -- | -- | -- | −3.10 | 2.92 × 10−17 | Down | -- | -- | -- |
| Gene Name | Gene ID | WT-GR vs. CR-NOR-like1-GR | WT-BR3 vs. CR-NOR-like1-BR+3 | WT-BR9 vs. CR-NOR-like1-BR+9 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Log2FC | p-Value | Type | Log2FC | p-Value | Type | Log2FC | p-Value | Type | ||
| Shikimate O-hydroxycinnamoyltransferase (HCT) | LOC101253556 | 5.90 | 5.02 × 10−12 | Up | -- | -- | -- | -- | -- | -- |
| LOC101248087 | 5.05 | 3.14 × 10−8 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101245886 | −2.20 | 3.09 × 10−6 | Down | 2.30 | 3.42 × 10−10 | Up | 2.18 | 3.96 × 10−5 | Up | |
| LOC101247305 | 7.39 | 1.06 × 10−4 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101260610 | 5.16 | 1.78 × 10−4 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101256271 | 1.60 | 1.93 × 10−3 | Up | -- | -- | -- | -- | -- | -- | |
| LOC101252161 | −2.31 | 2.14 × 10−3 | Down | -- | -- | -- | 2.28 | 7.77 × 10−4 | Up | |
| LOC101244961 | -- | -- | -- | −1.58 | 1.04 × 10−3 | Down | -- | -- | -- | |
| LOC101246106 | -- | -- | -- | −1.24 | 3.17 × 10−3 | Down | -- | -- | -- | |
| LOC101266953 | -- | -- | -- | 1.62 | 5.32 × 10−3 | Up | -- | -- | -- | |
| Caffeoyl-CoA O-methyltransferase (CCoAOMT) | LOC101253032 | 2.12 | 8.52 × 10−10 | Up | -- | -- | -- | -- | -- | -- |
| LOC101252203 | -- | -- | -- | 2.70 | 1.47 × 10−6 | Up | -- | -- | -- | |
| Trans-cinnamate 4-monooxygenase (C4H) | LOC101262919 | 3.28 | 2.83 × 10−6 | Up | -- | -- | -- | −2.45 | 1.56 × 10−6 | Down |
| LOC101244496 | -- | -- | -- | -- | -- | -- | 1.15 | 1.52 × 10−3 | Up | |
| Flavonol synthase (FLS) | LOC101260801 | 3.70 | 7.56 × 10−4 | Up | −1.51 | 4.18 × 10−3 | Down | -- | -- | -- |
| LOC101260380 | -- | -- | -- | -- | -- | -- | 9.94 | 2.70 × 10−16 | Up | |
| LOC101249699 | -- | -- | -- | -- | -- | -- | 1.73 | 2.63 × 10−5 | Up | |
| Chalcone isomerase (CHI) | CHI1 | −1.14 | 1.42 × 10−3 | Down | -- | -- | -- | -- | -- | -- |
| LOC101266223 | -- | -- | -- | -- | -- | -- | 2.84 | 3.54 × 10−9 | Up | |
| Coumaroylquinate(coumaroylshikimate) 3′-monooxygenase (C3H) | LOC101246092 | 1.77 | 3.55 × 10−3 | Up | 1.17 | 3.53 × 10−3 | Up | -- | -- | -- |
| Leucoanthocyanidin dioxygenase (LDOX) | LOC101248628 | -- | -- | -- | −3.34 | 1.70 × 10−6 | Down | -- | -- | -- |
| Naringenin 3-dioxygenase (F3H) | F3H | -- | -- | -- | -- | -- | -- | 2.51 | 6.12 × 10−10 | Up |
| Flavonoid 3′-monooxygenase (F3′H) | LOC101266618 | -- | -- | -- | -- | -- | -- | 2.31 | 2.27 × 10−9 | Up |
| Chalcone synthase (CHS) | CHS1 | -- | -- | -- | -- | -- | -- | 3.13 | 5.24 × 10−5 | Up |
| CHS2 | -- | -- | -- | -- | -- | -- | 1.19 | 8.59 × 10−4 | Up | |
| 2-hydroxyisoflavanone dehydratase (HIDH) | ASH1 | -- | -- | -- | 2.65 | 9.81 × 10−8 | Up | -- | -- | -- |
| Vestitone reductase (VR) | LOC101264524 | -- | -- | -- | 4.15 | 2.20 × 10−4 | Up | -- | -- | -- |
| Isoflavone/4′-methoxyisoflavone 2’-hydroxylase(I2′H) | LOC101250559 | -- | -- | -- | 1.68 | 9.19 × 10−3 | Up | 1.81 | 2.91 × 10−3 | Up |
| Flavonol-3-O-glucoside/galactoside glucosyltransferase (FG3) | LOC104649610 | -- | -- | -- | 1.73 | 1.05 × 10−5 | Up | -- | -- | -- |
| LOC101244316 | -- | -- | -- | 1.25 | 1.07 × 10−3 | Up | 4.20 | 5.14 × 10−6 | Up | |
| Gene Name | Gene ID | Compounds | PCC |
|---|---|---|---|
| HCT | LOC101252161 | Sinapinaldehyde | −0.85 |
| 5-O-p-Coumaroylquinic acid | −0.81 | ||
| LOC101244961 | p-Coumaraldehyde | 0.945 | |
| BGL | LOC101246223 | Sinapinaldehyde | −0.82 |
| 5-O-p-Coumaroylquinic acid | −0.83 | ||
| LOC101251735 | Sinapyl alcohol | 0.837 | |
| p-Coumaraldehyde | 0.879 | ||
| LOC101265077 | p-Coumaraldehyde | −0.89 | |
| Coniferin | −0.84 | ||
| LOC101256717 | p-Coumaraldehyde | −0.92 | |
| Coniferin | −0.82 | ||
| C4H | LOC101262919 | Sinapinaldehyde | 0.881 |
| TOGT1 | LOC101259704 | Sinapinaldehyde | −0.86 |
| 5-O-p-Coumaroylquinic acid | −0.82 | ||
| GAME1 | Coniferin | −0.89 | |
| p-Coumaraldehyde | −0.87 | ||
| LOC101258702 | Coniferin | 0.916 | |
| p-Coumaraldehyde | 0.897 | ||
| LOC101260915 | Ferulic acid * | 0.803 | |
| CAD | LOC101253340 | 5-O-p-Coumaroylquinic acid | −0.85 |
| LOC101250635 | Coniferin | −0.81 | |
| p-Coumaraldehyde | −0.88 | ||
| POD | LOC101244376 | Sinapinaldehyde | −0.97 |
| TMP1 | p-Coumaraldehyde | −0.94 | |
| Coniferin | −0.93 | ||
| TAP2 | Coniferin | −0.86 | |
| p-Coumaraldehyde | −0.9 | ||
| LOC101253648 | 7-Hydroxycoumarin | −0.86 | |
| p-Coumaraldehyde | −0.84 | ||
| CCR | LOC101250958 | Coniferin | −0.9 |
| p-Coumaraldehyde | −0.9 | ||
| LOC101265652 | p-Coumaraldehyde | −0.85 | |
| PAR2 | Coniferin | −0.81 | |
| p-Coumaraldehyde | −0.84 | ||
| 4CL | LOC101251363 | Coniferin | −0.82 |
| 5-O-p-Coumaroylquinic acid | −0.84 |
| Gene Name | Gene ID | Compounds | PCC |
|---|---|---|---|
| HCT | LOC101252161 | Naringenin chalcone | −0.816 |
| Pinobanksin | −0.803 | ||
| Hesperetin | −0.862 | ||
| 5-O-p-Coumaroylquinic acid | −0.813 | ||
| Phloretin | −0.815 | ||
| Butin | −0.821 | ||
| Aromadendrin (dihydrokaempferol) | −0.808 | ||
| Homoeriodictyol | −0.859 | ||
| LOC101253556 | Phloretin-2′-O-glucoside (phlorizin) | −0.868 | |
| LOC101247305 | Phloretin-2′-O-glucoside (phlorizin) | −0.902 | |
| LOC101244961 | Homoeriodictyol | 0.832 | |
| Hesperetin | 0.818 | ||
| Isosakuranetin (5,7-dihydroxy-4′-methoxyflavanone) | 0.925 | ||
| CHI | CHI1 | Eriodictyol (5,7,3′,4′-tetrahydroxyflavanone) | −0.846 |
| Homoeriodictyol | −0.834 | ||
| Pinobanksin | −0.823 | ||
| Naringenin (5,7,4′-trihydroxyflavanone) | −0.832 | ||
| Hesperetin | −0.825 | ||
| Aromadendrin (dihydrokaempferol) | −0.82 | ||
| C4H | LOC101262919 | Luteolin (5,7,3′,4′-tetrahydroxyflavone) | 0.832 |
| Naringenin (5,7,4′-trihydroxyflavanone) | 0.807 | ||
| Naringenin chalcone | 0.849 | ||
| Phloretin | 0.854 | ||
| Butin | 0.85 | ||
| Hesperetin | 0.839 | ||
| Aromadendrin (dihydrokaempferol) | 0.841 | ||
| Homoeriodictyol | 0.84 | ||
| Pinobanksin | 0.84 | ||
| CCOMT | LOC101253032 | Phloretin-2′-O-glucoside (Pplorizin) | −0.815 |
| CHS | CHS1 | Eriodictyol (5,7,3′,4′-tetrahydroxyflavanone) | 0.865 |
| Isoliquiritigenin | 0.832 | ||
| Luteolin (5,7,3′,4′-tetrahydroxyflavone) | 0.848 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Zhao, X.; Fu, D.; Zhao, Y. Integrated Analysis of Widely Targeted Metabolomics and Transcriptomics Reveals the Effects of Transcription Factor NOR-like1 on Alkaloids, Phenolic Acids, and Flavonoids in Tomato at Different Ripening Stages. Metabolites 2022, 12, 1296. https://doi.org/10.3390/metabo12121296
Yang X, Zhao X, Fu D, Zhao Y. Integrated Analysis of Widely Targeted Metabolomics and Transcriptomics Reveals the Effects of Transcription Factor NOR-like1 on Alkaloids, Phenolic Acids, and Flavonoids in Tomato at Different Ripening Stages. Metabolites. 2022; 12(12):1296. https://doi.org/10.3390/metabo12121296
Chicago/Turabian StyleYang, Xinyu, Xiaodan Zhao, Daqi Fu, and Ying Zhao. 2022. "Integrated Analysis of Widely Targeted Metabolomics and Transcriptomics Reveals the Effects of Transcription Factor NOR-like1 on Alkaloids, Phenolic Acids, and Flavonoids in Tomato at Different Ripening Stages" Metabolites 12, no. 12: 1296. https://doi.org/10.3390/metabo12121296