
Rational Engineering of Non-Ubiquinone Containing Corynebacterium glutamicum for Enhanced Coenzyme Q10 Production


 and
and
Abstract
:1. Introduction
2. Results
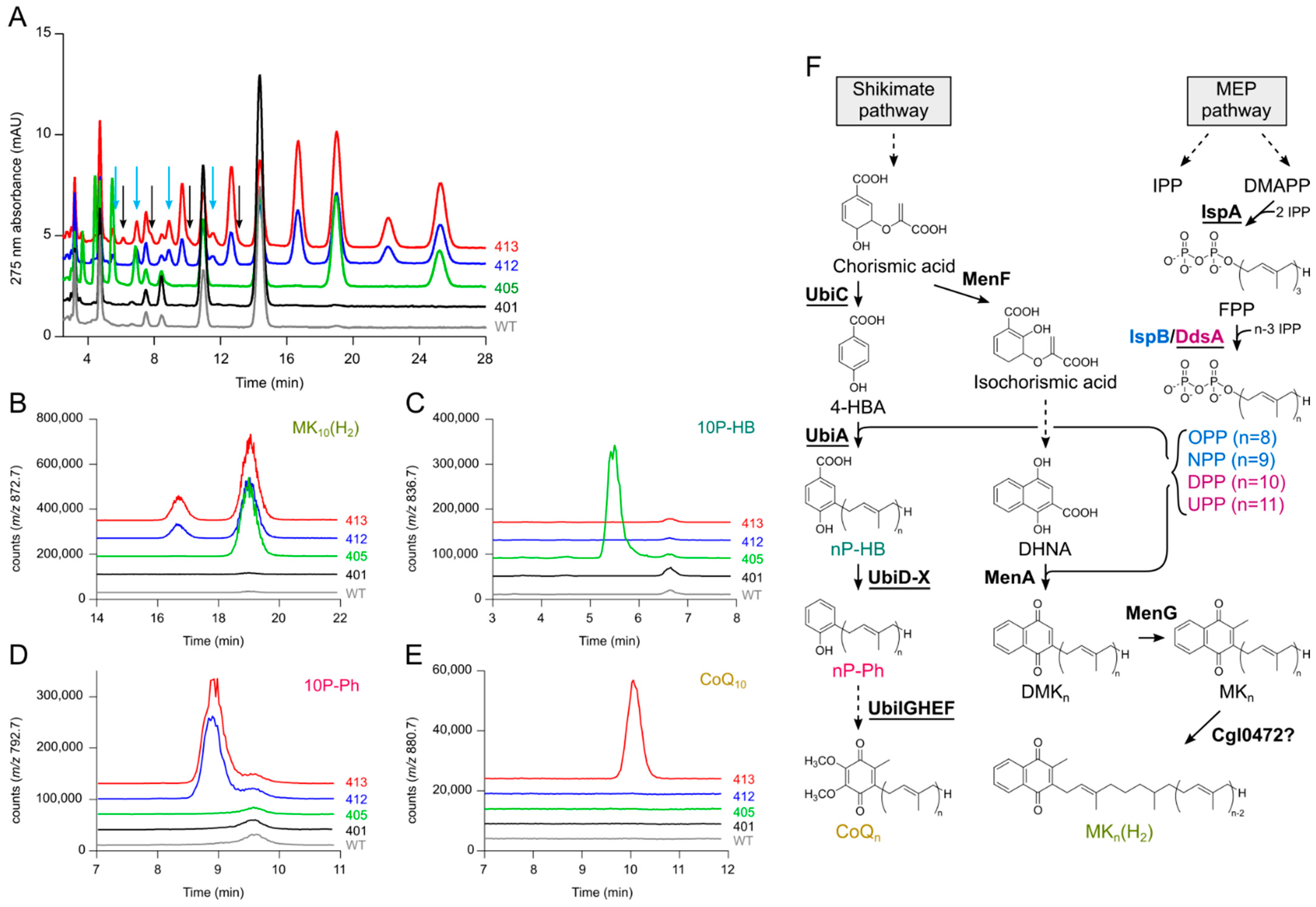
2.1. Identification of Accumulating Compounds in the Parent Strains
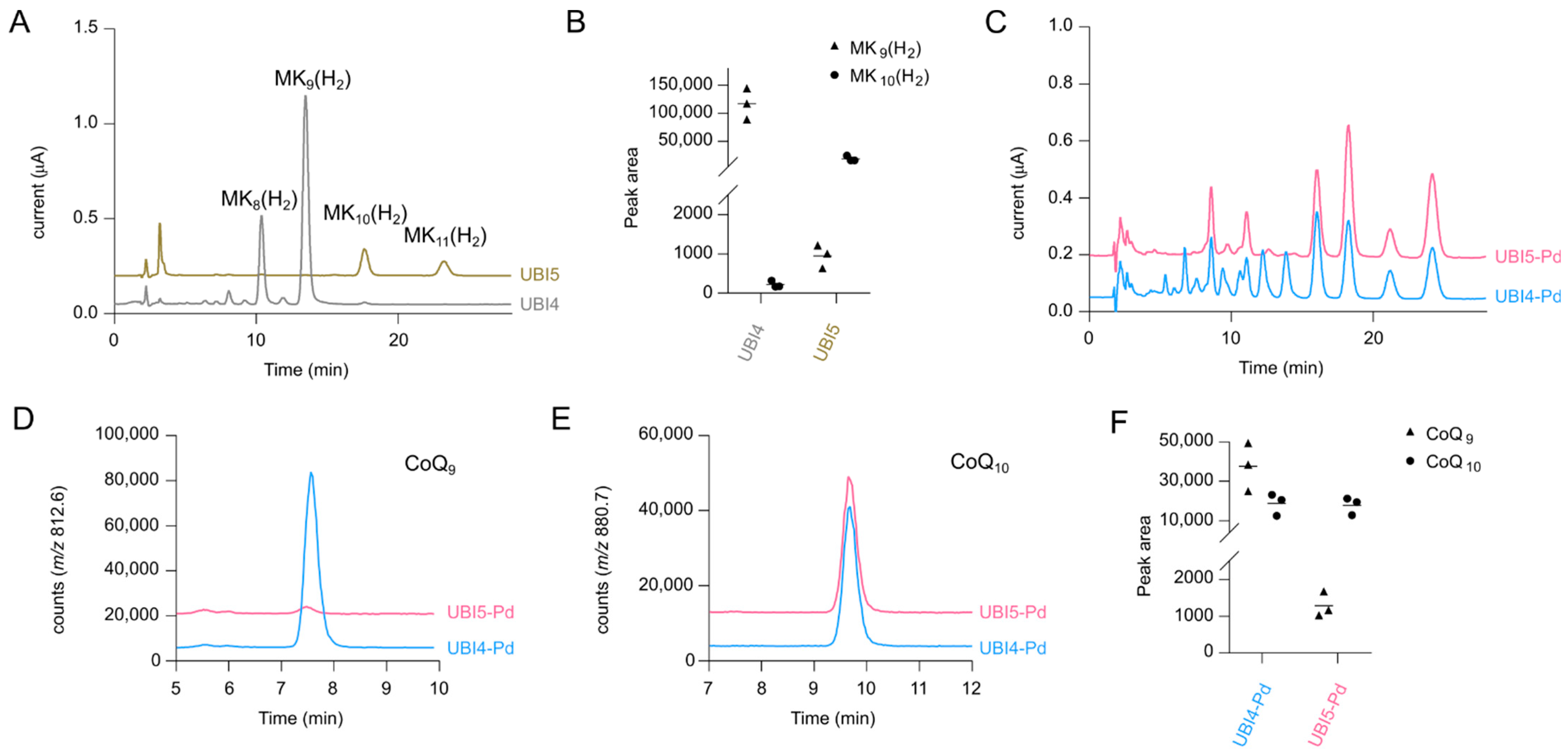
2.2. Deletion of ispB Diminishes Formation of 8- and 9-Isoprenologs
2.3. Screening of Different Decaprenyl Diphosphate Synthases
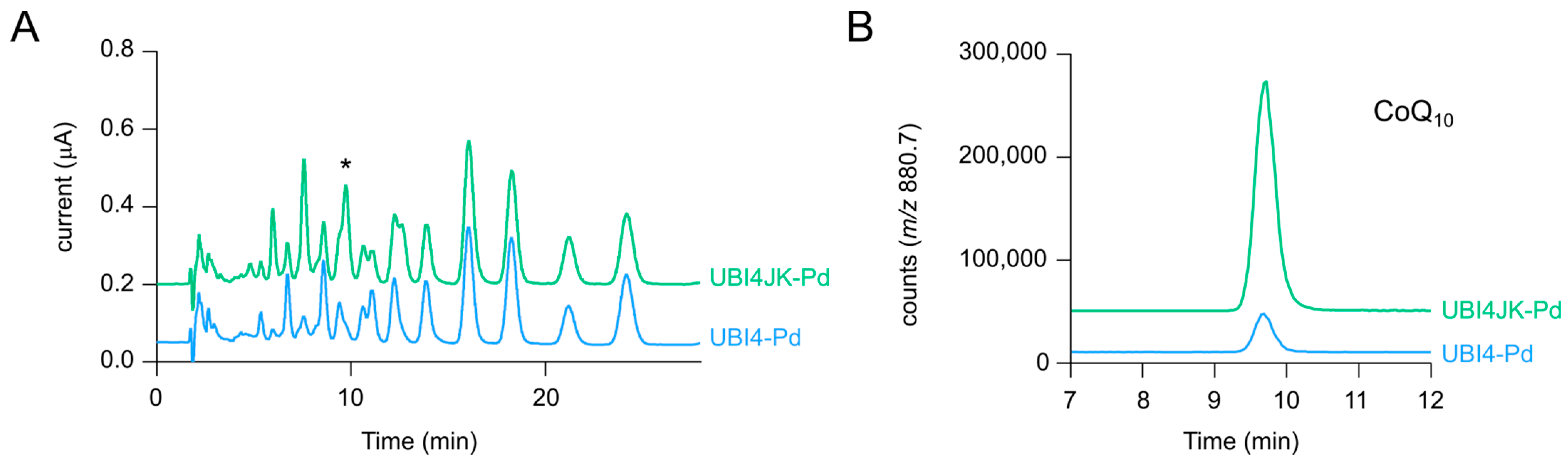
2.4. Expression of ubiJK Alleviates a Major Bottleneck
2.5. Combinatorial Approach for Maximized CoQ10 Production
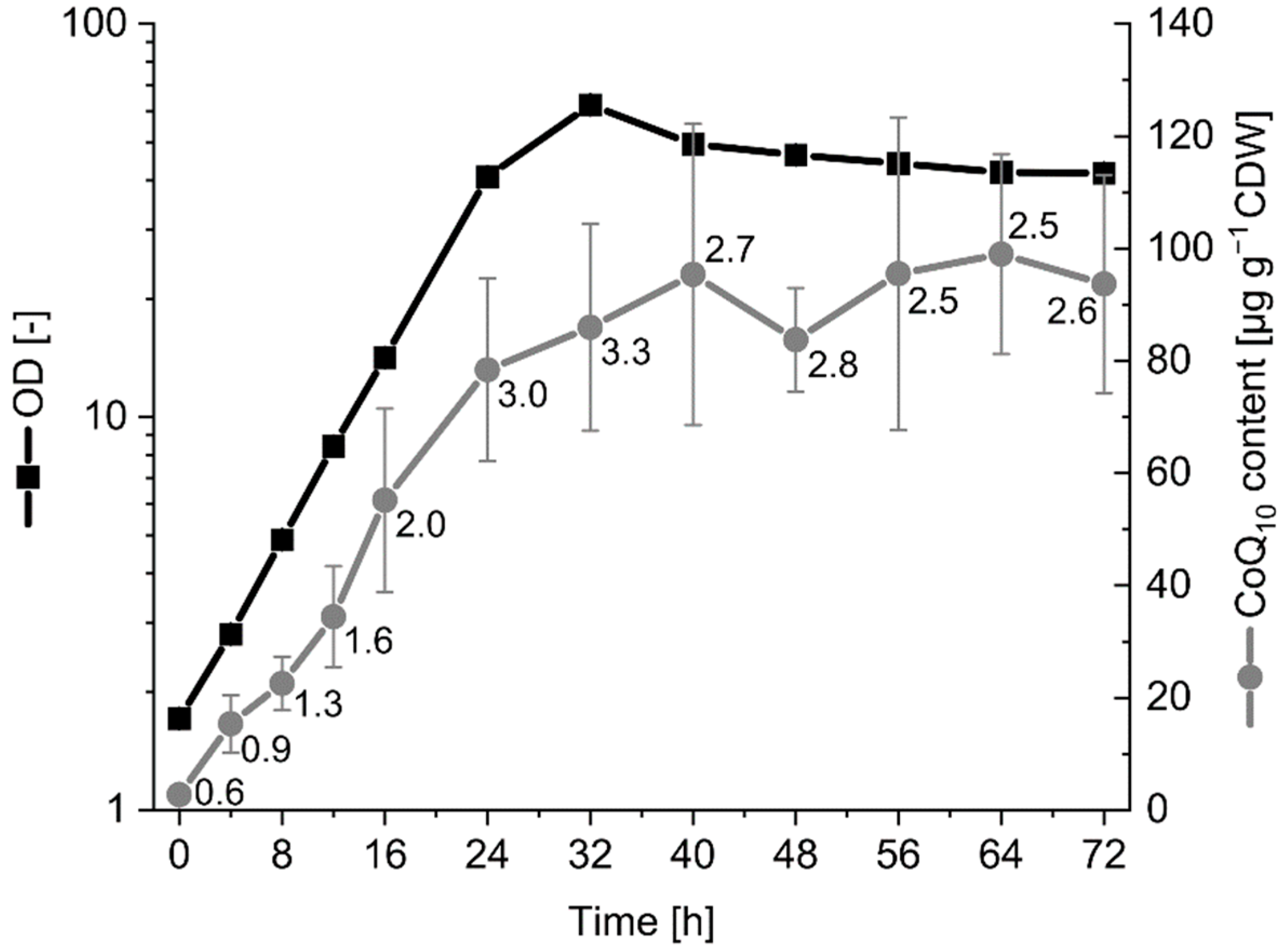
2.6. Influence of Growth Phase and Medium on CoQ10 Production
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Molecular Genetic Techniques and Strain Construction
4.3. Quinone Extraction and Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, A.M.; Smith, R.A.J.; Murphy, M.P. Antioxidant and Prooxidant Properties of Mitochondrial Coenzyme Q. Arch. Biochem. Biophys. 2004, 423, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Echtay, K.S.; Winkler, E.; Klingenberg, M. Coenzyme Q Is an Obligatory Cofactor for Uncoupling Protein Function. Nature 2000, 408, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Baschiera, E.; Sorrentino, U.; Calderan, C.; Desbats, M.A.; Salviati, L. The Multiple Roles of Coenzyme Q in Cellular Homeostasis and Their Relevance for the Pathogenesis of Coenzyme Q Deficiency. Free. Radic. Biol. Med. 2021, 166, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Camacho, J.D.; García-Corzo, L.; Fernández-Ayala, D.J.M.; Navas, P.; López-Lluch, G. Coenzyme Q at the Hinge of Health and Metabolic Diseases. Antioxidants 2021, 10, 1785. [Google Scholar] [CrossRef]
- Salviati, L.; Trevisson, E.; Doimo, M.; Navas, P. Primary Coenzyme Q10 Deficiency. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Mirzaa, G.M., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 2017. [Google Scholar]
- Hargreaves, I.; Heaton, R.A.; Mantle, D. Disorders of Human Coenzyme Q10 Metabolism: An Overview. Int. J. Mol. Sci. 2020, 21, 6695. [Google Scholar] [CrossRef] [PubMed]
- Yubero-Serrano, E.M.; Gonzalez-Guardia, L.; Rangel-Zuñiga, O.; Delgado-Lista, J.; Gutierrez-Mariscal, F.M.; Perez-Martinez, P.; Delgado-Casado, N.; Cruz-Teno, C.; Tinahones, F.J.; Villalba, J.M.; et al. Mediterranean Diet Supplemented With Coenzyme Q10 Modifies the Expression of Proinflammatory and Endoplasmic Reticulum Stress–Related Genes in Elderly Men and Women. J. Gerontol. Ser. A 2012, 67A, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, A.; Iannuzzo, G.; Parlato, A.; Cuomo, G.; Testa, C.; Coppola, M.; D’Ambrosio, G.; Oliviero, D.A.; Sarullo, S.; Vitale, G.; et al. Clinical Evidence for Q10 Coenzyme Supplementation in Heart Failure: From Energetics to Functional Improvement. J. Clin. Med. 2020, 9, 1266. [Google Scholar] [CrossRef]
- Arenas-Jal, M.; Suñé-Negre, J.M.; García-Montoya, E. Coenzyme Q10 Supplementation: Efficacy, Safety, and Formulation Challenges. Compr. Rev. Food Sci. Food Saf. 2020, 19, 574–594. [Google Scholar] [CrossRef] [Green Version]
- Žmitek, K.; Pogačnik, T.; Mervic, L.; Žmitek, J.; Pravst, I. The Effect of Dietary Intake of Coenzyme Q10 on Skin Parameters and Condition: Results of a Randomised, Placebo-Controlled, Double-Blind Study: The Effect of Dietary Intake of Coenzyme Q10 on Skin Parameters and Condition. BioFactors 2017, 43, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Yang, X.; Hu, J.; Ruan, X.; Mu, F.; Fu, Y. The Synthesis of Coenzyme Q10. Curr. Org. Chem. 2017, 21, 489–502. [Google Scholar] [CrossRef]
- Zhu, Y.; Ye, L.; Chen, Z.; Hu, W.; Shi, Y.; Chen, J.; Wang, C.; Li, Y.; Li, W.; Yu, H. Synergic Regulation of Redox Potential and Oxygen Uptake to Enhance Production of Coenzyme Q10 in Rhodobacter sphaeroides. Enzym. Microb. Technol. 2017, 101, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-S.; Yoo, J.-H.; Kim, S.-Y.; Pan, C.-H.; Kalia, V.C.; Kang, Y.C.; Lee, J.-K. Screening and Characterization of an Agrobacterium tumefaciens Mutant Strain Producing High Level of Coenzyme Q10. Process Biochem. 2015, 50, 33–39. [Google Scholar] [CrossRef]
- Zou, R.-S.; Li, S.; Zhang, L.-L.; Zhang, C.; Han, Y.-J.; Gao, G.; Sun, X.; Gong, X. Mutagenesis of Rhodobacter sphaeroides Using Atmospheric and Room Temperature Plasma Treatment for Efficient Production of Coenzyme Q10. J. Biosci. Bioeng. 2019, 127, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Abby, S.S.; Kazemzadeh, K.; Vragniau, C.; Pelosi, L.; Pierrel, F. Advances in Bacterial Pathways for the Biosynthesis of Ubiquinone. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148259. [Google Scholar] [CrossRef]
- Lu, W.; Ye, L.; Lv, X.; Xie, W.; Gu, J.; Chen, Z.; Zhu, Y.; Li, A.; Yu, H. Identification and Elimination of Metabolic Bottlenecks in the Quinone Modification Pathway for Enhanced Coenzyme Q10 Production in Rhodobacter sphaeroides. Metab. Eng. 2015, 29, 208–216. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, W.; Ye, L.; Chen, Z.; Hu, W.; Wang, C.; Chen, J.; Yu, H. Enhanced Synthesis of Coenzyme Q10 by Reducing the Competitive Production of Carotenoids in Rhodobacter sphaeroides. Biochem. Eng. J. 2017, 125, 50–55. [Google Scholar] [CrossRef]
- Martínez, I.; Méndez, C.; Berríos, J.; Altamirano, C.; Díaz-Barrera, A. Batch Production of Coenzyme Q10 by Recombinant Escherichia coli Containing the Decaprenyl Diphosphate Synthase Gene from Sphingomonas baekryungensis. J. Ind. Microbiol. Biotechnol. 2015, 42, 1283–1289. [Google Scholar] [CrossRef]
- Eggeling, L.; Bott, M. Handbook of Corynebacterium glutamicum; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-1-4200-3969-6. [Google Scholar]
- Burgardt, A.; Moustafa, A.; Persicke, M.; Sproß, J.; Patschkowski, T.; Risse, J.M.; Peters-Wendisch, P.; Lee, J.-H.; Wendisch, V.F. Coenzyme Q10 Biosynthesis Established in the Non-Ubiquinone Containing Corynebacterium glutamicum by Metabolic Engineering. Front. Bioeng. Biotechnol. 2021, 9, 650961. [Google Scholar] [CrossRef]
- Wendisch, V.F. Metabolic Engineering Advances and Prospects for Amino Acid Production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef]
- Prell, C.; Burgardt, A.; Meyer, F.; Wendisch, V.F. Fermentative Production of L-2-Hydroxyglutarate by Engineered Corynebacterium glutamicum via Pathway Extension of L-Lysine Biosynthesis. Front. Bioeng. Biotechnol. 2021, 8, 630476. [Google Scholar] [CrossRef]
- Kurpejović, E.; Wendisch, V.F.; Sariyar Akbulut, B. Tyrosinase-Based Production of L-DOPA by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2021, 105, 9103–9111. [Google Scholar] [CrossRef] [PubMed]
- Kerbs, A.; Mindt, M.; Schwardmann, L.; Wendisch, V.F. Sustainable Production of N-Methylphenylalanine by Reductive Methylamination of Phenylpyruvate Using Engineered Corynebacterium glutamicum. Microorganisms 2021, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Walter, T.; Al Medani, N.; Burgardt, A.; Cankar, K.; Ferrer, L.; Kerbs, A.; Lee, J.-H.; Mindt, M.; Risse, J.M.; Wendisch, V.F. Fermentative N-Methylanthranilate Production by Engineered Corynebacterium glutamicum. Microorganisms 2020, 8, 866. [Google Scholar] [CrossRef]
- Kogure, T.; Suda, M.; Hiraga, K.; Inui, M. Protocatechuate Overproduction by Corynebacterium glutamicum via Simultaneous Engineering of Native and Heterologous Biosynthetic Pathways. Metab. Eng. 2021, 65, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Kitade, Y.; Hashimoto, R.; Suda, M.; Hiraga, K.; Inui, M. Production of 4-Hydroxybenzoic Acid by an Aerobic Growth-Arrested Bioprocess Using Metabolically Engineered Corynebacterium glutamicum. Appl. Environ. Microbiol. 2018, 84, e02587-17. [Google Scholar] [CrossRef] [Green Version]
- Purwanto, H.S.; Kang, M.; Ferrer, L.; Han, S.; Lee, J.-Y.; Kim, H.; Lee, J. Rational Engineering of the Shikimate and Related Pathways in Corynebacterium glutamicum for 4-Hydroxybenzoate Production. J. Biotechnol. 2018, 282, 92–100. [Google Scholar] [CrossRef]
- Henke, N.A.; Wichmann, J.; Baier, T.; Frohwitter, J.; Lauersen, K.J.; Risse, J.M.; Peters-Wendisch, P.; Kruse, O.; Wendisch, V.F. Patchoulol Production with Metabolically Engineered Corynebacterium glutamicum. Genes 2018, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Henke, N.A.; Wendisch, V.F. Improved Astaxanthin Production with Corynebacterium glutamicum by Application of a Membrane Fusion Protein. Mar. Drugs 2019, 17, 621. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Swofford, C.A.; Rückert, C.; Chatzivasileiou, A.O.; Ou, R.W.; Opdensteinen, P.; Luttermann, T.; Zhou, K.; Stephanopoulos, G.; Jones Prather, K.L.; et al. Heterologous Production of α-Carotene in Corynebacterium glutamicum Using a Multi-Copy Chromosomal Integration Method. Bioresour. Technol. 2021, 341, 125782. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Peters-Wendisch, P.; Beekwilder, J.; Wendisch, V.F. IdsA Is the Major Geranylgeranyl Pyrophosphate Synthase Involved in Carotenogenesis in Corynebacterium glutamicum. FEBS J. 2014, 281, 4906–4920. [Google Scholar] [CrossRef]
- Loiseau, L.; Fyfe, C.; Aussel, L.; Hajj Chehade, M.; Hernández, S.B.; Faivre, B.; Hamdane, D.; Mellot-Draznieks, C.; Rascalou, B.; Pelosi, L.; et al. The UbiK Protein Is an Accessory Factor Necessary for Bacterial Ubiquinone (UQ) Biosynthesis and Forms a Complex with the UQ Biogenesis Factor UbiJ. J. Biol. Chem. 2017, 292, 11937–11950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, K.; Young, I.G. Alternative Hydroxylases for the Aerobic and Anaerobic Biosynthesis of Ubiquinone in Escherichia coli. Biochemistry 1978, 17, 4750–4755. [Google Scholar] [CrossRef] [PubMed]
- Jeya, M.; Moon, H.-J.; Lee, J.-L.; Kim, I.-W.; Lee, J.-K. Current State of Coenzyme Q10 Production and Its Applications. Appl. Microbiol. Biotechnol. 2010, 85, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Lu, H.; Zhang, G.; Ren, Z. Production of Coenzyme Q10 by Purple Non-Sulfur Bacteria: Current Development and Future Prospect. J. Clean. Prod. 2021, 307, 127326. [Google Scholar] [CrossRef]
- Hajj Chehade, M.; Pelosi, L.; Fyfe, C.D.; Loiseau, L.; Rascalou, B.; Brugière, S.; Kazemzadeh, K.; Vo, C.D.T.; Ciccone, L.; Aussel, L.; et al. A Soluble Metabolon Synthesizes the Isoprenoid Lipid Ubiquinone. Cell Chem. Biol. 2019, 26, 482–492.e7. [Google Scholar] [CrossRef]
- Veit, A.; Rittmann, D.; Georgi, T.; Youn, J.-W.; Eikmanns, B.J.; Wendisch, V.F. Pathway Identification Combining Metabolic Flux and Functional Genomics Analyses: Acetate and Propionate Activation by Corynebacterium glutamicum. J. Biotechnol. 2009, 140, 75–83. [Google Scholar] [CrossRef]
- Aussel, L.; Loiseau, L.; Chehade, M.H.; Pocachard, B.; Fontecave, M.; Pierrel, F.; Barras, F. UbiJ, a New Gene Required for Aerobic Growth and Proliferation in Macrophage, Is Involved in Coenzyme Q Biosynthesis in Escherichia coli and Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2014, 196, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Burgardt, A.; Prell, C.; Wendisch, V.F. Utilization of a Wheat Sidestream for 5-Aminovalerate Production in Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2021, 9, 732271. [Google Scholar] [CrossRef]
- Ioannidou, S.M.; Pateraki, C.; Ladakis, D.; Papapostolou, H.; Tsakona, M.; Vlysidis, A.; Kookos, I.K.; Koutinas, A. Sustainable Production of Bio-Based Chemicals and Polymers via Integrated Biomass Refining and Bioprocessing in a Circular Bioeconomy Context. Bioresour. Technol. 2020, 307, 123093. [Google Scholar] [CrossRef]
- Li, G.-W.; Burkhardt, D.; Gross, C.; Weissman, J.S. Quantifying Absolute Protein Synthesis Rates Reveals Principles Underlying Allocation of Cellular Resources. Cell 2014, 157, 624–635. [Google Scholar] [CrossRef] [Green Version]
- Tyedmers, J.; Mogk, A.; Bukau, B. Cellular Strategies for Controlling Protein Aggregation. Nat. Rev. Mol. Cell Biol. 2010, 11, 777–788. [Google Scholar] [CrossRef]
- Yim, S.S.; An, S.J.; Kang, M.; Lee, J.; Jeong, K.J. Isolation of Fully Synthetic Promoters for High-Level Gene Expression in Corynebacterium glutamicum. Biotechnol. Bioeng. 2013, 110, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Kugler, P.; Fröhlich, D.; Wendisch, V.F. Development of a Biosensor for Crotonobetaine-CoA Ligase Screening Based on the Elucidation of Escherichia coli Carnitine Metabolism. ACS Synth. Biol. 2020, 9, 2460–2471. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, F.; Jorge, J.M.P.; Dreyszas, A.; Risse, J.M.; Wendisch, V.F. Efficient Production of the Dicarboxylic Acid Glutarate by Corynebacterium glutamicum via a Novel Synthetic Pathway. Front. Microbiol. 2018, 9, 2589. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.N.; Lee, Y.; Hussein, R. Fundamental Relationship between Operon Organization and Gene Expression. Proc. Natl. Acad. Sci. USA 2011, 108, 10626–10631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, A.; Fontes, F.L.; Gonzalez-Juarrero, M.; McNeil, M.R.; Crans, D.C.; Jackson, M.; Crick, D.C. Partial Saturation of Menaquinone in Mycobacterium tuberculosis: Function and Essentiality of a Novel Reductase, MenJ. ACS Cent. Sci. 2015, 1, 292–302. [Google Scholar] [CrossRef]
- Xu, W.; Yang, S.; Zhao, J.; Su, T.; Zhao, L.; Liu, J. Improving Coenzyme Q8 Production in Escherichia coli Employing Multiple Strategies. J. Ind. Microbiol. Biotechnol. 2014, 41, 1297–1303. [Google Scholar] [CrossRef]
- Xu, W.; Yao, J.; Liu, L.; Ma, X.; Li, W.; Sun, X.; Wang, Y. Improving Squalene Production by Enhancing the NADPH/NADP+ Ratio, Modifying the Isoprenoid-Feeding Module and Blocking the Menaquinone Pathway in Escherichia coli. Biotechnol. Biofuels 2019, 12, 68. [Google Scholar] [CrossRef]
- Iqbal, I.K.; Bajeli, S.; Akela, A.K.; Kumar, A. Bioenergetics of Mycobacterium: An Emerging Landscape for Drug Discovery. Pathogens 2018, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Kurosu, M.; Narayanasamy, P.; Biswas, K.; Dhiman, R.; Crick, D.C. Discovery of 1,4-Didydroxy-2-Naphthoate Prenyltransferase Inhibitors. J. Med. Chem. 2007, 50, 3973–3975. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Shi, F.; Tan, S.; Yu, X.; Lai, W.; Li, Y. Engineering a Bifunctional ComQXPA-PsrfA Quorum-Sensing Circuit for Dynamic Control of Gene Expression in Corynebacterium glutamicum. ACS Synth. Biol. 2021, 10, 1761–1774. [Google Scholar] [CrossRef] [PubMed]
- Pierrel, F.; Burgardt, A.; Lee, J.-H.; Pelosi, L.; Wendisch, V.F. Recent Advances in the Metabolic Pathways and Microbial Production of Coenzyme Q. World J. Microbiol. Biotechnol. 2022, 38, 58. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sun, Y.; Ramos, K.R.M.; Nisola, G.M.; Valdehuesa, K.N.G.; Lee, W.; Park, S.J.; Chung, W.-J. Combination of Entner-Doudoroff Pathway with MEP Increases Isoprene Production in Engineered Escherichia coli. PLoS ONE 2013, 8, e83290. [Google Scholar] [CrossRef] [PubMed]
- Göttl, V.L.; Schmitt, I.; Braun, K.; Peters-Wendisch, P.; Wendisch, V.F.; Henke, N.A. CRISPRi-Library-Guided Target Identification for Engineering Carotenoid Production by Corynebacterium glutamicum. Microorganisms 2021, 9, 670. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, L.; Wang, C.; Choi, E.-S.; Kim, S.-W. Enhanced Performance of the Methylerythritol Phosphate Pathway by Manipulation of Redox Reactions Relevant to IspC, IspG, and IspH. J. Biotechnol. 2017, 248, 1–8. [Google Scholar] [CrossRef]
- Wu, T.; Ye, L.; Zhao, D.; Li, S.; Li, Q.; Zhang, B.; Bi, C.; Zhang, X. Membrane Engineering—A Novel Strategy to Enhance the Production and Accumulation of β-Carotene in Escherichia coli. Metab. Eng. 2017, 43, 85–91. [Google Scholar] [CrossRef]
- Ha, S.; Kim, S.; Seo, J.; Moon, H.; Lee, K.-M.; Lee, J. Controlling the Sucrose Concentration Increases Coenzyme Q10 Production in Fed-Batch Culture of Agrobacterium tumefaciens. Appl. Microbiol. Biotechnol. 2007, 76, 109–116. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, L.; Wang, K.-F.; Xu, L.; Zhou, L.; Wang, W.; Li, C.; Xu, Z.; Shi, T.; Chen, H.; et al. Phosphate Limitation Increases Coenzyme Q10 Production in Industrial Rhodobacter sphaeroides HY01. Synth. Syst. Biotechnol. 2019, 4, 212–219. [Google Scholar] [CrossRef]
- Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Almanaa, T.N.; Innasimuthu, G.M.; Rajoo, B.; Alanzi, K.F.; Rajaram, S.K. Optimization of Glutamic Acid Production by Corynebacterium glutamicum Using Response Surface Methodology. J. King Saud Univ.-Sci. 2020, 32, 1403–1408. [Google Scholar] [CrossRef]
- Kiefer, D.; Merkel, M.; Lilge, L.; Hausmann, R.; Henkel, M. High Cell Density Cultivation of Corynebacterium glutamicum on Bio-Based Lignocellulosic Acetate Using pH-Coupled Online Feeding Control. Bioresour. Technol. 2021, 340, 125666. [Google Scholar] [CrossRef]
- Wendisch, V.F.; Nampoothiri, K.M.; Lee, J.-H. Metabolic Engineering for Valorization of Agri- and Aqua-Culture Sidestreams for Production of Nitrogenous Compounds by Corynebacterium glutamicum. Front. Microbiol. 2022, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Baritugo, K.-A.G.; Kim, H.T.; David, Y.C.; Choi, J.H.; Choi, J.; Kim, T.W.; Park, C.; Hong, S.H.; Na, J.-G.; Jeong, K.J.; et al. Recent Advances in Metabolic Engineering of Corynebacterium glutamicum as a Potential Platform Microorganism for Biorefinery. Biofuels Bioprod. Biorefining 2018, 12, 899–925. [Google Scholar] [CrossRef]
- Zhang, B.; Jiang, Y.; Li, Z.; Wang, F.; Wu, X.-Y. Recent Progress on Chemical Production From Non-Food Renewable Feedstocks Using Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2020, 8, 606047. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, F.; Klein, V.J.; Brito, L.F.; Brautaset, T. From Brown Seaweed to a Sustainable Microbial Feedstock for the Production of Riboflavin. Front. Bioeng. Biotechnol. 2022, 10, 863690. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Studies on Transformation of Escherichia coli with Plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for In Vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Gauttam, R.; Desiderato, C.; Jung, L.; Shah, A.; Eikmanns, B.J. A Step Forward: Compatible and Dual-Inducible Expression Vectors for Gene Co-Expression in Corynebacterium glutamicum. Plasmid 2019, 101, 20–27. [Google Scholar] [CrossRef]
- Kirchner, O.; Tauch, A. Tools for Genetic Engineering in the Amino Acid-Producing Bacterium Corynebacterium glutamicum. J. Biotechnol. 2003, 104, 287–299. [Google Scholar] [CrossRef]
- Stansen, C.; Uy, D.; Delaunay, S.; Eggeling, L.; Goergen, J.-L.; Wendisch, V.F. Characterization of a Corynebacterium glutamicum Lactate Utilization Operon Induced during Temperature-Triggered Glutamate Production. Appl. Environ. Microbiol. 2005, 71, 5920–5928. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Ruan, Y.; Zhu, L.; Li, Q. Improving the Electro-Transformation Efficiency of Corynebacterium glutamicum by Weakening Its Cell Wall and Increasing the Cytoplasmic Membrane Fluidity. Biotechnol. Lett. 2015, 37, 2445–2452. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic Assembly of DNA Molecules up to Several Hundred Kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small Mobilizable Multi-Purpose Cloning Vectors Derived from the Escherichia coli Plasmids PK18 and PK19: Selection of Defined Deletions in the Chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Henke, N.A.; Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F. Production of the Marine Carotenoid Astaxanthin by Metabolically Engineered Corynebacterium glutamicum. Mar. Drugs 2016, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.E.; Peters-Wendisch, P.; Netzer, R.; Stafnes, M.; Brautaset, T.; Wendisch, V.F. Production and Glucosylation of C50 and C40 Carotenoids by Metabolically Engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 1223–1235. [Google Scholar] [CrossRef]
- Kabus, A.; Niebisch, A.; Bott, M. Role of Cytochrome bd Oxidase from Corynebacterium glutamicum in Growth and Lysine Production. Appl. Environ. Microbiol. 2007, 73, 861–868. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 10P-Ph/ 9P-Ph | CoQ10/ CoQ9 | CoQ10/ CoQ11 | Yx (µg g−1 CDW) | Titer (mg L−1) | Vol. Productivity (µg L−1 h−1) |
|---|---|---|---|---|---|---|
| UBI4-Pd | 1.1 ± 0.1 | 0.5 ± 0.0 | 1.2 ± 0.1 | 18.2 ± 5.4 | 0.15 ± 0.05 | 2.1 ± 0.6 |
| UBI4-At | 1.2 ± 0.3 | 0.6 ± 0.2 | 145.4 ± 12.4 *** | 21.3 ± 4.6 | 0.14 ± 0.04 | 2.0 ± 0.6 |
| UBI4-Rs | 1.6 ± 0.2 ** | 0.9 ± 0.1 ** | 7.6 ± 0.0 *** | 24.9 ± 5.9 | 0.18 ± 0.04 | 2.5 ± 0.6 |
| Strain | CoQ10/ 10P-Ph | CoQ10/ CoQ9 | Yx (µg g−1 CDW) | Titer (mg L−1) | Vol. Productivity (µg L−1 h−1) |
|---|---|---|---|---|---|
| UBI4-Pd | 0.3 ± 0.1 | 0.5 ± 0.0 | 18.2 ± 5.4 | 0.15 ± 0.05 | 2.1 ± 0.6 |
| UBI4JK-Pd | 1.5 ± 0.2 *** | 0.7 ± 0.1 * | 78.0 ± 12.0 ** | 0.64 ± 0.08 *** | 9.0 ± 1.1 *** |
| UBI5-Pd | 0.2 ± 0.1 | 14.4 ± 5.5 * | 17.3 ± 4.4 | 0.15 ± 0.04 | 2.1 ± 0.6 |
| UBI6-Pd | 1.2 ± 0.2 ** | 38.6 ± 1.9 *** | 69.6 ± 9.4 ** | 0.58 ± 0.06 *** | 8.0 ± 0.9 *** |
| Strain/ medium | CoQ10/ 10P-Ph | CoQ10/ CoQ9 | CoQ10/ CoQ11 | Yx (µg g−1 CDW) | Titer (mg L−1) | Vol. Productivity (µg L−1 h−1) |
|---|---|---|---|---|---|---|
| UBI6-Pd | 1.2 ± 0.2 | 38.6 ± 1.9 | 1.5 ± 0.0 | 69.6 ± 9.4 | 0.58 ± 0.06 | 8.0 ± 0.9 |
| UBI6-At | 1.2 ± 0.2 | 5.1 ± 0.4 ** | 3.5 ± 0.5 ** | 64.3 ± 4.6 | 0.61 ± 0.04 | 8.4 ± 0.6 |
| UBI6-Rs | 1.9 ± 0.5 | 41.6 ± 3.4 | 3.4 ± 0.2 *** | 126.9 ± 10.7 ** | 1.21 ± 0.12 ** | 16.8 ± 1.7 ** |
| Microcultivation of UBI6-Rs in CGXII medium and WSCH medium | ||||||
| CGXII | 1.0 ± 0.2 | 55.4 ± 7.2 | 4.2 ± 0.1 | 92.2 ± 17.2 | 0.89 ± 0.15 | 12.3 ± 2.1 |
| WSCH | 1.5 ± 0.2 | 31.4 ± 1.1 | 8.8 ± 0.6 | 37.7 ± 7.4 | 0.49 ± 0.08 | 6.8 ± 1.2 |
| Strains | Description | Source |
|---|---|---|
| Corynebacterium glutamicum | ||
| WT | C. glutamicum wild-type strain ATCC 13032 | ATCC |
| UBI4 | WT with following modifications: ΔcrtOP (cg0717-cg0723), ΔidsA (cg2384), ΔcrtB2I’I2 (cg2668-cg2672), LP4::Ptuf-ispA (ispA from E. coli), ΔpobA (cg1226), ΔpcaHG::Psod-ubiCFBR (cg2631-cg2630, ubiCL31A from E. coli), Δvdh::PilvC-aroGFBR (cg2953, aroGD146N from E. coli), ΔqsuABCD::Ptuf-qsuC (cg0501-cg0504); named UBI400 in [20] | [20] |
| UBI401 | UBI4 carrying pRG_Duet2, pEC-XT99A, and pEKEx3 | This work |
| UBI405 | UBI4 carrying pRG_Duet2-ddsAPd-ubiA, pEC-XT99A, and pEKEx3 | This work |
| UBI412 | UBI4 carrying pRG_Duet2-ddsAPd-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3 | This work |
| UBI4-Pd | UBI4 carrying pRG_Duet2-ddsAPd-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF; named UBI413 in [20] | [20] |
| UBI4-At | UBI4 carrying pRG_Duet2-ddsAAt-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| UBI4-Rs | UBI4 carrying pRG_Duet2-ddsARs-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| UBI5 | ΔispB::Ptuf-ddsAPd mutant of UBI4 | This work |
| UBI5-Pd | UBI5 carrying pRG_Duet2-ddsAPd-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| UBI4JK | ΔactA::ubiJK mutant of UBI4 | This work |
| UBI4JK-Pd | UBI4JK carrying pRG_Duet2-ddsAPd-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| UBI6 | ΔispB::Ptuf-ddsAPd mutant of UBI4JK | This work |
| UBI6-Pd | UBI6 carrying pRG_Duet2-ddsAPd-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| UBI6-At | UBI6 carrying pRG_Duet2-ddsAAt-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| UBI6-Rs | UBI6 carrying pRG_Duet2-ddsARs-ubiA, pEC-XT99A-ubiDIBX, and pEKEx3-ubiGHEF | This work |
| Escherichia coli | ||
| DH5α | F-thi-1 endA1 hsdr17(r-, m-) supE44 1lacU169 (Φ80lacZ1M15) recA1 gyrA96 | [67] |
| S17-1 | recA pro hsdR RP4-2-Tc::Mu-Km::Tn7 | [68] |
| Plasmids | Description | Source |
|---|---|---|
| pRG_Duet2 | KanR, Ptac, lacIq, PtetR/tetA, tetR, pBL1 oriVCg, dual-inducible C. glutamicum/E. coli expression shuttle vector | [69] |
| pRG_Duet2-ddsAPd-ubiA | KanR, pRG_Duet2 overexpressing ddsA from P. denitrificans (induced by IPTG) and ubiA from E. coli (induced by ATc) | [20] |
| pRG_Duet2-ddsAAt-ubiA | KanR, pRG_Duet2 overexpressing ddsA from A. tumefaciens (induced by IPTG) and ubiA from E. coli (induced by ATc) | This work |
| pRG_Duet2-ddsARs-ubiA | KanR, pRG_Duet2 overexpressing ddsA from R. sphaeroides (induced by IPTG) and ubiA from E. coli (induced by ATc) | This work |
| pEC-XT99A | TetR, Ptrc, lacIq, pGA1 oriVCg, C. glutamicum/E. coli expression shuttle vector | [70] |
| pEC-XT99A-ubiDIBX | TetR, pEC-XT99A overexpressing ubiD, ubiI, ubiB, and ubiX from E. coli | [20] |
| pEKEx3 | SpecR, Ptac, lacIq, pBL1 oriVCg, C. glutamicum/E. coli expression shuttle vector | [71] |
| pEKEx3-ubiGHEF | SpecR, pEKEx3 overexpressing ubiG, ubiH, ubiE, and ubiF from E. coli | [20] |
| pK19mobsacB | KanR, pK19 oriVEc, sacB, lacZα, E. coli/C. glutamicum shuttle vector for construction of insertion and deletion mutants in C. glutamicum | [75] |
| pK19mobsacB-ΔactA:ubiJK | pK19mobsacB with a construct for deletion of actA (cg2840) and insertion of ubiJ and ubiK from E. coli under control of the native actA promoter | This work |
| pK19mobsacB-ΔispB:Ptuf-ddsAPd | pK19mobsacB with a construct for deletion of ispB (cg0559) and insertion of ddsA from P. denitrificans under control of C. glutamicum promoter Ptuf | This work |
| Primers | Sequence (5′ to 3′) |
|---|---|
| ddsA_At-fw | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGGGCGTCGTCATACCGCTTG |
| ddsA_At-rv | GAGCTCGGTACCCGGGGATCTTAGTTGAGACGCTCGATGCAG |
| ddsA_Rs-fw | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGGGATTGGACGAGGTTTC |
| ddsA_Rs-rv | GAGCTCGGTACCCGGGGATCTTAGGCGATGCGTTCGAC |
| actA-US-fw | GCATGCCTGCAGGTCGACTCTAGAGTCCCGTGCGTTGCATTTCCTG |
| actA-US-rv | CGGTTTCTAAACCAAGAAAAAACGGATCCCAGGTAATCGGACTTTTTCAAATTTTTCCC |
| actA-DS-fw | ATTTGAAAAAGTCCGATTACCTGGGATCCGTTTTTTCTTGGTTTAGAAACCG |
| actA-DS-rv | AATTCGAGCTCGGTACCCGGGGATCAGCCAATCGTCGTAAAGCG |
| ubiJ-fw | AATTTGAAAAAGTCCGATTACCTGGCTCCCCCTTAGTAGAAAAGGAGGTTTTTCTATGCCTTTTAAACCTTTAGTGACG |
| ubiJ-rv | CTCAATTTTTTTCGGGTCAATCATCTGAAGGGCCTCCTTTCTCATTTAGCCTCCAGTTTTTCC |
| ubiK-fw | GGAAAAACTGGAGGCTAAATGAGAAAGGAGGCCCTTCAGATGATTGACCCGAAAAAAATTGAG |
| ubiK-rv | TTTCTAAACCAAGAAAAAACGGATCTTACAGCGTTGGGGGGAGAG |
| actA-conf-fw | TTTCATCCGGCGCGAAGGTG |
| actA-conf-rv | GCTTCTGCGCAAAGCAAGCC |
| pSH1-ddsA-fw | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGGGCATGAACGAAAACGT |
| pSH1-ddsA-rv | GAGCTCGGTACCCGGGGATCTTAGGACAGGCGCGAGACGA |
| ispB-US-fw | CCTGCAGGTCGACTCTAGAGTCATGAGATTTTGCCAAGCGG |
| ispB-US-rv | GGTTAAGTGGTGGATTACGGGGACTAGTTCATCGCTACCTTTGGTGATCG |
| ispB-DS-fw | CGATCACCAAAGGTAGCGATGAACTAGTCCCCGTAATCCACCACTTAACC |
| ispB-DS-rv | GAGCTCGGTACCCGGGGATCTATGAGAAGTCAGCACACGC |
| Ptuf-ddsA-fw | CTCGATCACCAAAGGTAGCGATGAATGGCCGTTACCCTGCGAATG |
| Ptuf-ddsA-rv | TTAAGTGGTGGATTACGGGGACTAGTTAGGACAGGCGCGAGACGAC |
| ispB-conf-fw | ATCACATGCTTCGCCTTGAC |
| ispB-conf-rv | TTTCTCGAAGGCAACACCTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgardt, A.; Pelosi, L.; Chehade, M.H.; Wendisch, V.F.; Pierrel, F. Rational Engineering of Non-Ubiquinone Containing Corynebacterium glutamicum for Enhanced Coenzyme Q10 Production. Metabolites 2022, 12, 428. https://doi.org/10.3390/metabo12050428
Burgardt A, Pelosi L, Chehade MH, Wendisch VF, Pierrel F. Rational Engineering of Non-Ubiquinone Containing Corynebacterium glutamicum for Enhanced Coenzyme Q10 Production. Metabolites. 2022; 12(5):428. https://doi.org/10.3390/metabo12050428
Chicago/Turabian StyleBurgardt, Arthur, Ludovic Pelosi, Mahmoud Hajj Chehade, Volker F. Wendisch, and Fabien Pierrel. 2022. "Rational Engineering of Non-Ubiquinone Containing Corynebacterium glutamicum for Enhanced Coenzyme Q10 Production" Metabolites 12, no. 5: 428. https://doi.org/10.3390/metabo12050428
APA StyleBurgardt, A., Pelosi, L., Chehade, M. H., Wendisch, V. F., & Pierrel, F. (2022). Rational Engineering of Non-Ubiquinone Containing Corynebacterium glutamicum for Enhanced Coenzyme Q10 Production. Metabolites, 12(5), 428. https://doi.org/10.3390/metabo12050428