
Sea Bass Essence from Lates calcarifer Improves Exercise Performance and Anti-Fatigue in Mice


 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Effect of SBE Supplementation on Exercise Performance
2.2. Effect of SBE Supplementation on Serum Lactate Levels after the 10-min Swim Test
2.3. Effect of SBE Supplementation on Fatigue-Related Biochemical Indicators after the 10-min Swim Test or a 90-min Swim Test and a 60-min Rest
2.4. Effect of SBE Supplementation on Liver and Muscle Glycogen Contents
2.5. Effect of SBE Supplementation on Biochemical Variables at the End of the Experiment
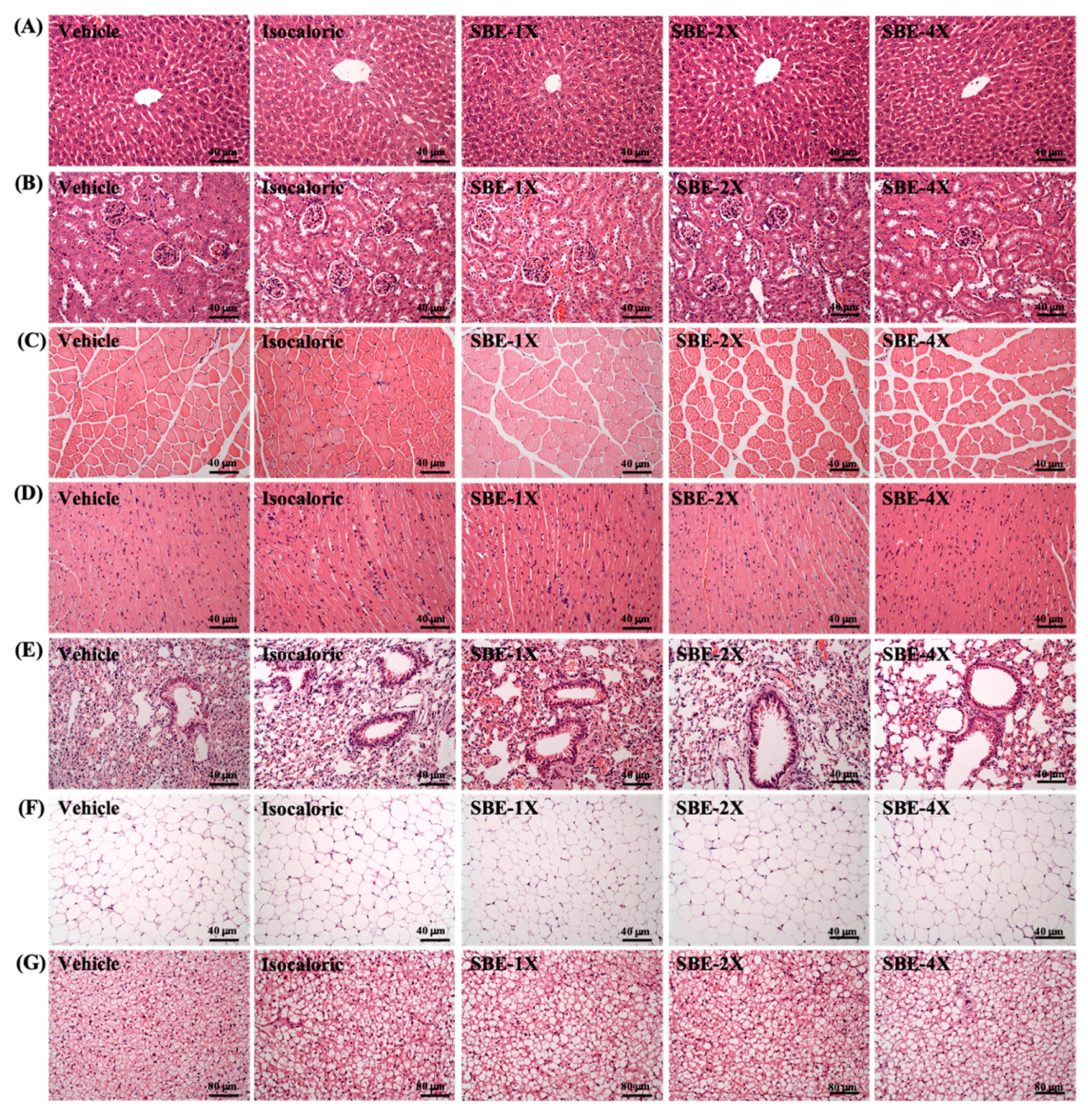
2.6. Subchronic Toxicity Evaluation of SBE Supplementation
3. Discussion
4. Materials and Methods
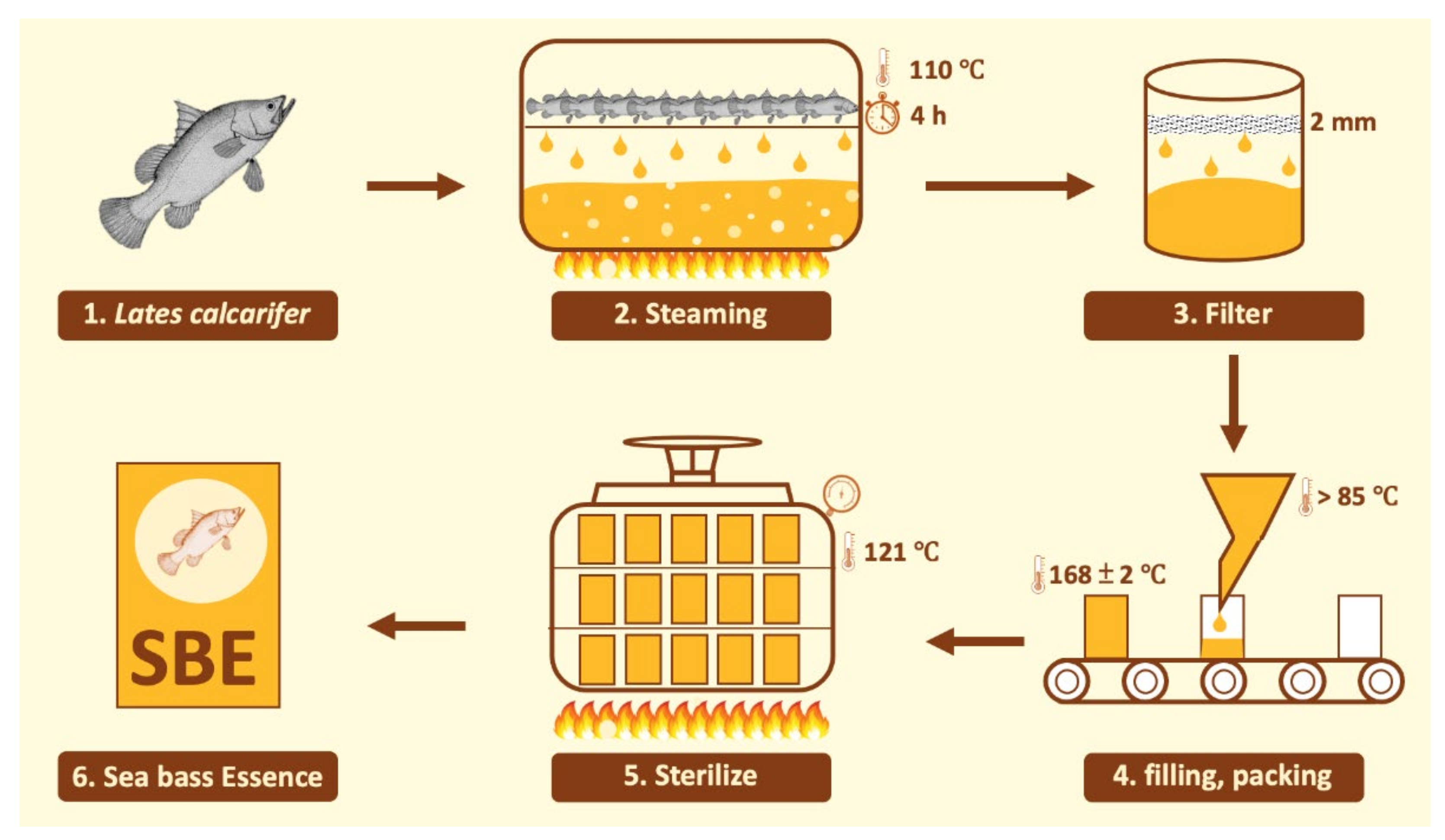
4.1. Hi-Q Sea Bass Essence (SBE) Preparation
4.2. Experimental Design
4.3. Swimming Exercise Performance Test
4.4. Determination of Fatigue-Associated Biochemical Variables
4.5. Clinical Biochemical Profiles
4.6. Visceral Tissue Weight and Histology Staining and Glycogen Determination
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wan, J.J.; Qin, Z.; Wang, P.Y.; Sun, Y.; Liu, X. Muscle fatigue: General understanding and treatment. Exp. Mol. Med. 2017, 49, e384. [Google Scholar] [CrossRef] [PubMed]
- Vashistha, V.; Singh, B.; Kaur, S.; Prokop, L.J.; Kaushik, D. The Effects of Exercise on Fatigue, Quality of Life, and Psychological Function for Men with Prostate Cancer: Systematic Review and Meta-analyses. Eur. Urol. Focus. 2016, 2, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.T.; Fuchs, C.J.; Betts, J.A.; van Loon, L.J. Liver glycogen metabolism during and after prolonged endurance-type exercise. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E543–E553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, S.K.; Nelson, W.B.; Hudson, M.B. Exercise-induced oxidative stress in humans: Cause and consequences. Free Radic. Biol. Med. 2011, 51, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Baskaran, R.; Shibu, M.A.; Lin, W.T. Anti-Fatigue and Exercise Performance Improvement Effect of Glossogyne tenuifolia Extract in Mice. Nutrients 2022, 14, 1011. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.L. Fatigue mediates the effects of exercise on quality of life. Qual. Life Res. 1999, 8, 529–538. [Google Scholar] [CrossRef]
- Aoi, W.; Naito, Y.; Yoshikawa, T. Exercise and functional foods. Nutr. J. 2006, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Michalak, M.; Agellon, L.B. Importance of Nutrients and Nutrient Metabolism on Human Health. Yale J. Biol. Med. 2018, 91, 95–103. [Google Scholar]
- Guo, Z.; Lin, D.; Guo, J.; Zhang, Y.; Zheng, B. In Vitro Antioxidant Activity and In Vivo Anti-Fatigue Effect of Sea Horse (Hippocampus) Peptides. Molecules 2017, 22, 482. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Lin, H.; Deng, S.G. Study of Anti-Fatigue Effect in Rats of Ferrous Chelates Including Hairtail Protein Hydrolysates. Nutrients 2015, 7, 9860–9871. [Google Scholar] [CrossRef] [Green Version]
- Borow, K.M.; Nelson, J.R.; Mason, R.P. Biologic plausibility, cellular effects, and molecular mechanisms of eicosapentaenoic acid (EPA) in atherosclerosis. Atherosclerosis 2015, 242, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacon, A.G.; Metian, M. Food matters: Fish, income, and food supply—A comparative analysis. Rev. Fish. Sci. Aquac. 2018, 26, 15–28. [Google Scholar] [CrossRef]
- Gajanan, P.G.; Elavarasan, K.; Shamasundar, B.A. Bioactive and functional properties of protein hydrolysates from fish frame processing waste using plant proteases. Environ. Sci. Pollut. Res. Int. 2016, 23, 24901–24911. [Google Scholar] [CrossRef] [PubMed]
- Phadke, G.G.; Rathod, N.B.; Ozogul, F.; Elavarasan, K.; Karthikeyan, M.; Shin, K.H.; Kim, S.K. Exploiting of Secondary Raw Materials from Fish Processing Industry as a Source of Bioactive Peptide-Rich Protein Hydrolysates. Mar. Drugs 2021, 19, 480. [Google Scholar] [CrossRef]
- Manninen, A.H. Protein hydrolysates in sports and exercise: A brief review. J. Sports Sci. Med. 2004, 3, 60–63. [Google Scholar]
- Majidiyan, N.; Hadidi, M.; Azadikhah, D.; Moreno, A. Protein complex nanoparticles reinforced with industrial hemp essential oil: Characterization and application for shelf-life extension of Rainbow trout fillets. Food Chem. X 2022, 13, 100202. [Google Scholar] [CrossRef]
- Bertrais, S.; Galan, P.; Renault, N.; Zarebska, M.; Preziosi, P.; Hercberg, S. Consumption of soup and nutritional intake in French adults: Consequences for nutritional status. J. Hum. Nutr. Diet. 2001, 14, 121–128. [Google Scholar] [CrossRef]
- Leeb, E.; Kulozik, U.; Cheison, S. Thermal pre-treatment of β-Lactoglobulin as a tool to steer enzymatic hydrolysis and control the release of peptides. Procedia Food Sci. 2011, 1, 1540–1546. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Tao, N.; Su, H.; Zhang, J.; Zhong, J. Migration of nutrients and formation of micro/nano-sized particles in Atlantic salmon (Salmo salar) and bighead carp (Aristichthys nobilis) head soups. Food Biosci. 2020, 36, 100646. [Google Scholar] [CrossRef]
- Rengpipat, S.; Rueangruklikhit, T.; Piyatiratitivorakul, S. Evaluations of lactic acid bacteria as probiotics for juvenile seabass Lates calcarifer. Aquac. Res. 2008, 39, 134–143. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Pateiro, M.; Domínguez, R.; Zhou, J.; Barba, F.J.; Lorenzo, J.M. Nutritional Characterization of Sea Bass Processing By-Products. Biomolecules 2020, 10, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekjing, S.; Venkatachalam, K.; Wangbenmad, C. Biochemical evaluation of novel seabass (Lates calcarifer) fish essence soup prepared by prolonged boiling process. Arabian J. Chem. 2021, 14, 103365. [Google Scholar] [CrossRef]
- Hsu, T.H.; Chiu, C.C.; Wang, Y.C.; Chen, T.H.; Chen, Y.H.; Lee, Y.P.; Hung, S.W.; Wu, C.P.; Chuang, H.L. Supplementation with Beef Extract Improves Exercise Performance and Reduces Post-Exercise Fatigue Independent of Gut Microbiota. Nutrients 2018, 10, 1740. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.W.; Hsu, Y.J.; Lee, M.C.; Li, H.S.; Yeo, P.C.W.; Lim, A.L.; Huang, C.C. In Vitro and In Vivo Functional Characterization of Essence of Chicken as an Ergogenic Aid. Nutrients 2018, 10, 1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [Green Version]
- Samples, S. Towards a more sustainable production of fish as an important source for human nutrition. J. Fish. Livest. Prod. 2014, 2, 119. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Manoli, I.; Venditti, C.P. Disorders of branched chain amino acid metabolism. Transl. Sci. Rare Dis. 2016, 1, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Adeva-Andany, M.M.; López-Maside, L.; Donapetry-García, C.; Fernández-Fernández, C.; Sixto-Leal, C. Enzymes involved in branched-chain amino acid metabolism in humans. Amino Acids 2017, 49, 1005–1028. [Google Scholar] [CrossRef]
- Felig, P.; Wahren, J. Amino acid metabolism in exercising man. J. Clin. Investig. 1971, 50, 2703–2714. [Google Scholar] [CrossRef] [Green Version]
- Ahlborg, G.; Felig, P.; Hagenfeldt, L.; Hendler, R.; Wahren, J. Substrate turnover during prolonged exercise in man. Splanchnic and leg metabolism of glucose, free fatty acids, and amino acids. J. Clin. Investig. 1974, 53, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- De Campos-Ferraz, P.L.; Ribeiro, S.M.; Luz Sdos, S.; Lancha, A.H., Jr.; Tirapegui, J. Exercise x BCAA Supplementation in Young Trained Rats: What are their Effects on Body Growth? J. Sports Sci. Med. 2011, 10, 483–490. [Google Scholar] [PubMed]
- De Araujo, J.A., Jr.; Falavigna, G.; Rogero, M.M.; Pires, I.S.; Pedrosa, R.G.; Castro, I.A.; Donato, J., Jr.; Tirapegui, J. Effect of chronic supplementation with branched-chain amino acids on the performance and hepatic and muscle glycogen content in trained rats. Life Sci. 2006, 79, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcinko, K.; Steinberg, G.R. The role of AMPK in controlling metabolism and mitochondrial biogenesis during exercise. Exp. Physiol. 2014, 99, 1581–1585. [Google Scholar] [CrossRef] [Green Version]
- Gibala, M.J.; Little, J.P.; Macdonald, M.J.; Hawley, J.A. Physiological adaptations to low-volume, high-intensity interval training in health and disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef]
- Ørtenblad, N.; Westerblad, H.; Nielsen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef]
- Shimomura, Y.; Murakami, T.; Nakai, N.; Nagasaki, M.; Obayashi, M.; Li, Z.; Xu, M.; Sato, Y.; Kato, T.; Shimomura, N.; et al. Supression of glycogen consumption during acute exercise by dietary branched-chain amino acids in rats. J. Nutr. Sci. Vitamol. 2000, 46, 71–77. [Google Scholar] [CrossRef]
- Matsumoto, K.; Koba, T.; Hamada, K.; Tsujimoto, H.; Mitsuzono, R. Branched-chain amino acid supplementation increases the lactate threshold during an incremental exercise test in trained individuals. J. Nutr. Sci. Vitaminol. 2009, 55, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Lin, C.I.; Chiu, C.C.; Lin, Y.T.; Huang, W.K.; Huang, H.Y.; Huang, C.C. Chicken essence improves exercise performance and ameliorates physical fatigue. Nutrients 2014, 6, 2681–2696. [Google Scholar] [CrossRef] [Green Version]
- Falavigna, G.; Alves de Araújo, J., Jr.; Rogero, M.M.; Pires, I.S.; Pedrosa, R.G.; Martins, E., Jr.; Alves de Castro, I.; Tirapegui, J. Effects of diets supplemented with branched-chain amino acids on the performance and fatigue mechanisms of rats submitted to prolonged physical exercise. Nutrients 2012, 4, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M. Dietary supplements and sports performance: Amino acids. J. Int. Soc. Sports Nutr. 2005, 2, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.L.; Lee, M.C.; Hsu, Y.J.; Huang, W.C.; Huang, C.C.; Huang, S.W. Isolated Soy Protein Supplementation and Exercise Improve Fatigue-Related Biomarker Levels and Bone Strength in Ovariectomized Mice. Nutrients 2018, 10, 1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Kim, S.H.; Jeong, W.S.; Lee, H.Y. Effect of BCAA intake during endurance exercises on fatigue substances, muscle damage substances, and energy metabolism substances. J. Exerc. Nutrition Biochem. 2013, 17, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef] [PubMed]
- De Palo, E.F.; Gatti, R.; Cappellin, E.; Schiraldi, C.; De Palo, C.B.; Spinella, P. Plasma lactate, GH and GH-binding protein levels in exercise following BCAA supplementation in athletes. Amino Acids. 2001, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- MacLean, D.A.; Graham, T.E.; Saltin, B. Stimulation of muscle ammonia production during exercise following branched-chain amino acid supplementation in humans. J Physiol. 1996, 493, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Holecek, M.; Kandar, R.; Sispera, L.; Koverik, M. Acute hyperammonemia activates branched-chain amino acid catabolism and decreases their extracellular concentrations: Different sensitivity of red and white muscle. Amino Acids 2011, 40, 575–584. [Google Scholar] [CrossRef]
- Kitaoka, Y. McArdle Disease and Exercise Physiology. Biology 2014, 3, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Korzeniewski, B. AMP deamination delays muscle acidification during heavy exercise and hypoxia. J. Biol. Chem. 2006, 281, 3057–3066. [Google Scholar] [CrossRef] [Green Version]
- Mikulski, T.; Dabrowski, J.; Hilgier, W.; Ziemba, A.; Krzeminski, K. Effects of supplementation with branched chain amino acids and ornithine aspartate on plasma ammonia and central fatigue during exercise in healthy men. Folia Neuropathol. 2015, 53, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Hayakawa, F.; Xichang, W.; Shunsheng, C.; Yokoyama, M.; Kasai, M.; Takeutchi, F.; Hatae, K. Comparative study on flavor preference between Japanese and Chinese for dried bonito stock and chicken bouillon. J. Food Sci. 2005, 70, S193–S198. [Google Scholar] [CrossRef]
- Hsu, Y.J.; Lee, M.C.; Huang, C.C.; Ho, C.S. The effects of different types of aquatic exercise training interventions on a high-fructose diet-fed mice. Int. J. Med. Sci. 2021, 18, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.J.; Jhang, W.L.; Lee, M.C.; Bat-Otgon, B.; Narantungalag, E.; Huang, C.C. Lactose-riched Mongolian mare’s milk improves physical fatigue and exercise performance in mice. Int. J. Med. Sci. 2021, 18, 564–574. [Google Scholar] [CrossRef]
- Lee, M.C.; Hsu, Y.J.; Ho, H.H.; Kuo, Y.W.; Lin, W.Y.; Tsai, S.Y.; Chen, W.L.; Lin, C.L.; Huang, C.C. Effectiveness of human-origin Lactobacillus plantarum PL-02 in improving muscle mass, exercise performance and anti-fatigue. Sci. Rep. 2021, 11, 19469. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Vehicle | Isocaloric | SBE-1X | SBE-2X | SBE-5X | |
|---|---|---|---|---|---|---|
| Time Point | Lactate (mmol/L) | |||||
| Before swimming (A) | 3.34 ± 0.38 a | 3.32 ± 0.32 a | 3.25 ± 0.37 a | 3.45 ± 0.29 a | 3.33 ± 0.36 a | |
| After swimming (B) | 7.65 ± 0.57 c | 7.26 ± 0.50 c | 6.18 ± 0.71 b | 6.00 ± 0.72 ab | 5.54 ± 0.74 a | |
| After a 20 min resting (C) | 6.39 ± 0.47 c | 6.09 ± 0.54 c | 4.92 ± 0.52 b | 4.76 ± 0.42 b | 4.27 ± 0.52 a | |
| Rates of lactate production and clearance | ||||||
| Production rate = B/A | 2.30 ± 0.10 e | 2.19 ± 0.09 d | 1.90 ± 0.06 c | 1.74 ± 0.08 b | 1.66 ± 0.06 a | |
| Clearance rate = (B − C)/B | 0.16 ± 0.01 a | 0.16 ± 0.03 a | 0.20 ± 0.04 b | 0.21 ± 0.03 b | 0.23 ± 0.04 b | |
| Groups | Vehicle | Isocaloric | SBE-1X | SBE-2X | SBE-5X | |
|---|---|---|---|---|---|---|
| Parameters | ||||||
| AST (U/L) | 73 ± 11 | 71 ± 7 | 75 ± 8 | 72 ± 6 | 74 ± 5 | |
| ALT (U/L) | 47 ± 5 | 47 ± 5 | 49 ± 5 | 47 ± 5 | 47 ± 5 | |
| ALB (mg/dL) | 3.44 ± 0.11 | 3.28 ± 0.23 | 3.37 ± 0.24 | 3.37 ± 0.32 | 3.35 ± 0.21 | |
| BUN (mg/dL) | 27.1 ± 3.7 | 26.1 ± 1.9 | 26.3 ± 2.3 | 26.2 ± 2.6 | 26.3 ± 2.7 | |
| CREA (mg/dL) | 0.43 ± 0.02 | 0.43 ± 0.02 | 0.44 ± 0.03 | 0.44 ± 0.03 | 0.43 ± 0.03 | |
| UA (mg/dL) | 2.1 ± 0.8 | 2.1 ± 0.5 | 2.0 ± 0.5 | 2.2 ± 0.4 | 2.1 ± 0.8 | |
| TP (mg/dL) | 5.7 ± 0.4 | 5.7 ± 0.4 | 5.7 ± 0.3 | 5.8 ± 0.3 | 5.8 ± 0.3 | |
| TG (mg/dL) | 130 ± 12 | 131 ± 16 | 131 ± 13 | 129 ± 12 | 129 ± 10 | |
| CK (U/L) | 252 ± 48 | 269 ± 46 | 259 ± 47 | 269 ± 49 | 269 ± 46 | |
| Characteristics | Vehicle | Isocaloric | SBE-1X | SBE-2X | SBE-5X |
|---|---|---|---|---|---|
| Initial BW (g) | 29.9 ± 0.7 | 29.7 ± 0.6 | 29.7 ± 0.9 | 29.7 ± 0.7 | 29.7 ± 0.4 |
| 1st wk BW | 33.8 ± 1.1 | 33.5 ± 1.4 | 33.3 ± 0.7 | 33.4 ± 1.3 | 33.4 ± 1.2 |
| 2nd wk BW | 35.5 ± 1.9 | 35.5 ± 1.4 | 35.2 ± 1.4 | 34.8 ± 1.6 | 34.4 ± 1.3 |
| 3rd wk BW | 36.6 ± 2.0 | 36.8 ± 2.0 | 36.7 ± 2.0 | 36.2 ± 2.2 | 35.7 ± 1.5 |
| 4th wk BW | 37.4 ± 2.4 | 37.6 ± 2.1 | 37.5 ± 2.2 | 36.9 ± 2.3 | 36.5 ± 1.7 |
| 5th wk BW | 37.9 ± 2.5 | 38.4 ± 2.1 | 37.9 ± 2.3 | 37.4 ± 2.2 | 36.9 ± 1.7 |
| Final BW (g) | 38.8 ± 2.7 | 39.0 ± 2.2 | 39.0 ± 1.6 | 38.5 ± 2.2 | 38.0 ± 1.2 |
| Water intake (mL/mouse/day) | 7.1 ± 0.4 | 7.2 ± 0.4 | 7.2 ± 0.5 | 7.1 ± 0.6 | 7.2 ± 0.5 |
| Diet (g/mouse/day) | 6.1 ± 0.9 | 6.2 ± 0.9 | 6.3 ± 0.8 | 6.1 ± 0.9 | 6.3 ± 0.7 |
| Calorie intake from diet (Chow 5001) (Kcal/mouse/day) (A) | 20.5 ± 3.1 | 20.8 ± 2.9 | 21.2 ± 2.8 | 20.4 ± 3.0 | 21.1 ± 2.4 |
| Calorie intake from supplements (Kcal/mouse/day) (B) | 0.0 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 b | 0.3 ± 0.0 c | 0.5 ± 0.1 c |
| Total daily calorie intake (Kcal/mouse/day) (A) + (B) | 20.5 ± 3.1 | 20.9 ± 2.9 | 21.3 ± 2.8 | 20.7 ± 3.0 | 21.7 ± 2.4 |
| Liver (g) | 2.34 ± 0.30 | 2.31 ± 0.30 | 2.25 ± 0.21 | 2.29 ± 0.31 | 2.35 ± 0.16 |
| Kidney (g) | 0.64 ± 0.06 | 0.64 ± 0.08 | 0.64 ± 0.05 | 0.63 ± 0.05 | 0.63 ± 0.04 |
| Muscle (g) | 0.37 ± 0.03 | 0.38 ± 0.02 | 0.39 ± 0.04 | 0.36 ± 0.05 | 0.36 ± 0.03 |
| Heart (g) | 0.21 ± 0.03 | 0.21 ± 0.02 | 0.23 ± 0.02 | 0.21 ± 0.02 | 0.21 ± 0.02 |
| Lung (g) | 0.26 ± 0.03 | 0.26 ± 0.03 | 0.26 ± 0.03 | 0.26 ± 0.03 | 0.26 ± 0.04 |
| EFP (g) | 0.44 ± 0.08 | 0.43 ± 0.07 | 0.44 ± 0.05 | 0.43 ± 0.07 | 0.43 ± 0.05 |
| BAT (g) | 0.11 ± 0.03 | 0.10 ± 0.02 | 0.11 ± 0.02 | 0.11 ± 0.02 | 0.09 ± 0.02 |
| Relative liver weight (%) | 5.98 ± 0.38 | 5.86 ± 0.54 | 5.73 ± 0.57 | 5.88 ± 0.59 | 6.14 ± 0.27 |
| Relative kidney weight (%) | 1.63 ± 0.20 | 1.63 ± 0.17 | 1.62 ± 0.07 | 1.64 ± 0.14 | 1.64 ± 0.10 |
| Relative muscle weight (%) | 0.96 ± 0.10 | 0.98 ± 0.05 | 0.98 ± 0.12 | 0.94 ± 0.12 | 0.95 ± 0.08 |
| Relative heart weight (%) | 0.55 ± 0.07 | 0.52 ± 0.06 | 0.58 ± 0.07 | 0.54 ± 0.05 | 0.54 ± 0.05 |
| Relative lung weight (%) | 0.67 ± 0.08 | 0.67 ± 0.09 | 0.66 ± 0.07 | 0.66 ± 0.05 | 0.68 ± 0.11 |
| Relative EFP weight (%) | 1.12 ± 0.18 | 1.10 ± 0.17 | 1.11 ± 0.12 | 0.95 ± 0.03 | 0.93 ± 0.08 |
| Relative BAT weight (%) | 0.28 ± 0.07 | 0.26 ± 0.05 | 0.27 ± 0.04 | 0.28 ± 0.06 | 0.24 ± 0.05 |
| Nutrition Facts | 100 mL SBE |
|---|---|
| Total calories (kcal) | 30.4 |
| Protein (g/100 mL) | 7.6 |
| Fat (g/100 mL) | - |
| Saturated fat (g/100 mL) | - |
| Trans fat (g/100 mL) | - |
| Moisture (g/100 mL) | 94.0 |
| Sodium (mg/100 mL) | 39.6 |
| Carbohydrate (g/100 mL) | - |
| Total BCAA (leucine, isoleucine, and valine) | 6.86% in protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tai, H.-J.; Lee, M.-C.; Hsu, Y.-J.; Kuo, C.-Y.; Huang, C.-C.; Wang, M.-F. Sea Bass Essence from Lates calcarifer Improves Exercise Performance and Anti-Fatigue in Mice. Metabolites 2022, 12, 531. https://doi.org/10.3390/metabo12060531
Tai H-J, Lee M-C, Hsu Y-J, Kuo C-Y, Huang C-C, Wang M-F. Sea Bass Essence from Lates calcarifer Improves Exercise Performance and Anti-Fatigue in Mice. Metabolites. 2022; 12(6):531. https://doi.org/10.3390/metabo12060531
Chicago/Turabian StyleTai, Hong-Jun, Mon-Chien Lee, Yi-Ju Hsu, Chun-Yen Kuo, Chi-Chang Huang, and Ming-Fu Wang. 2022. "Sea Bass Essence from Lates calcarifer Improves Exercise Performance and Anti-Fatigue in Mice" Metabolites 12, no. 6: 531. https://doi.org/10.3390/metabo12060531