
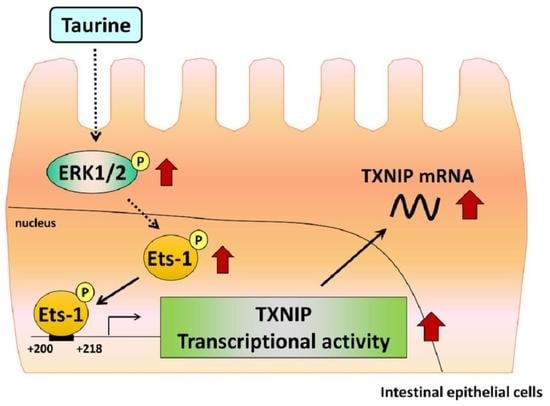
Signaling Pathway of Taurine-Induced Upregulation of TXNIP

 ,
,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Results
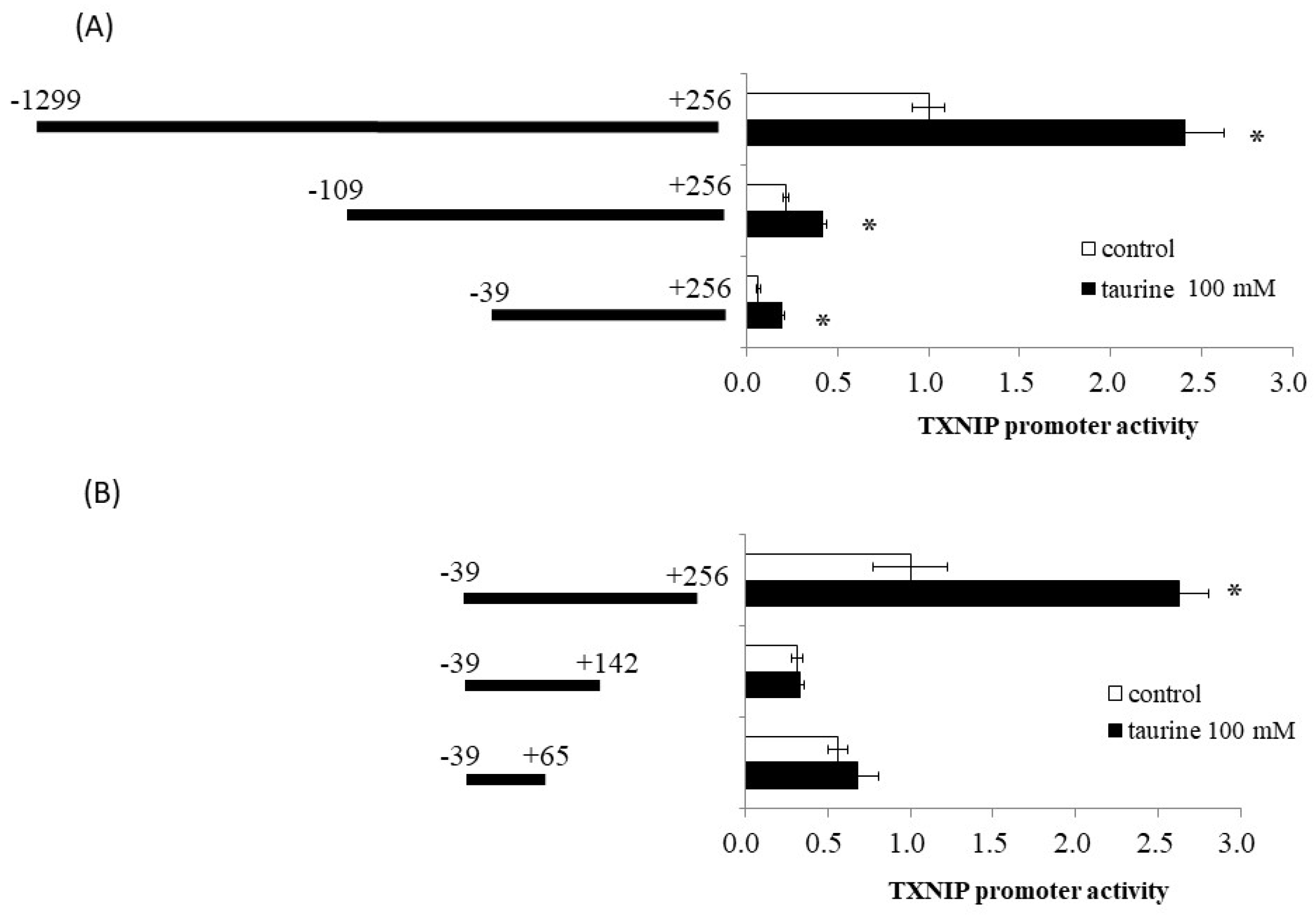
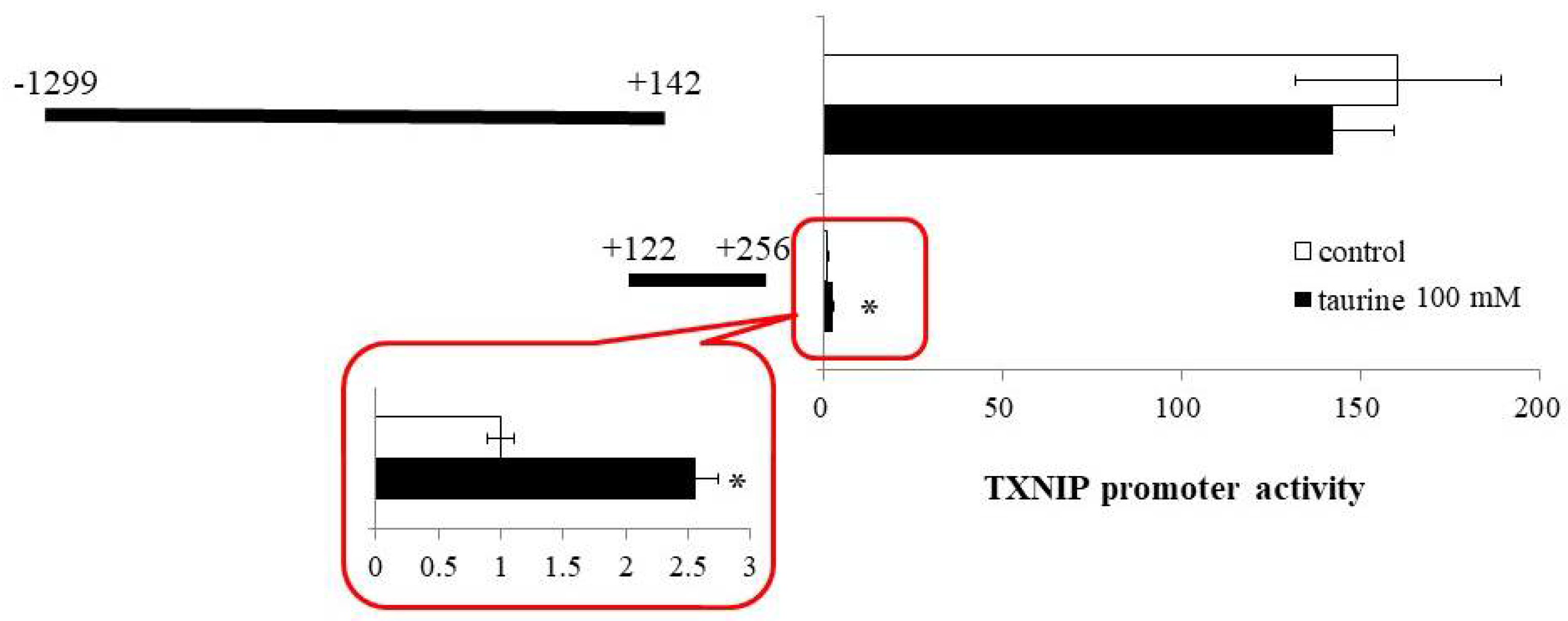
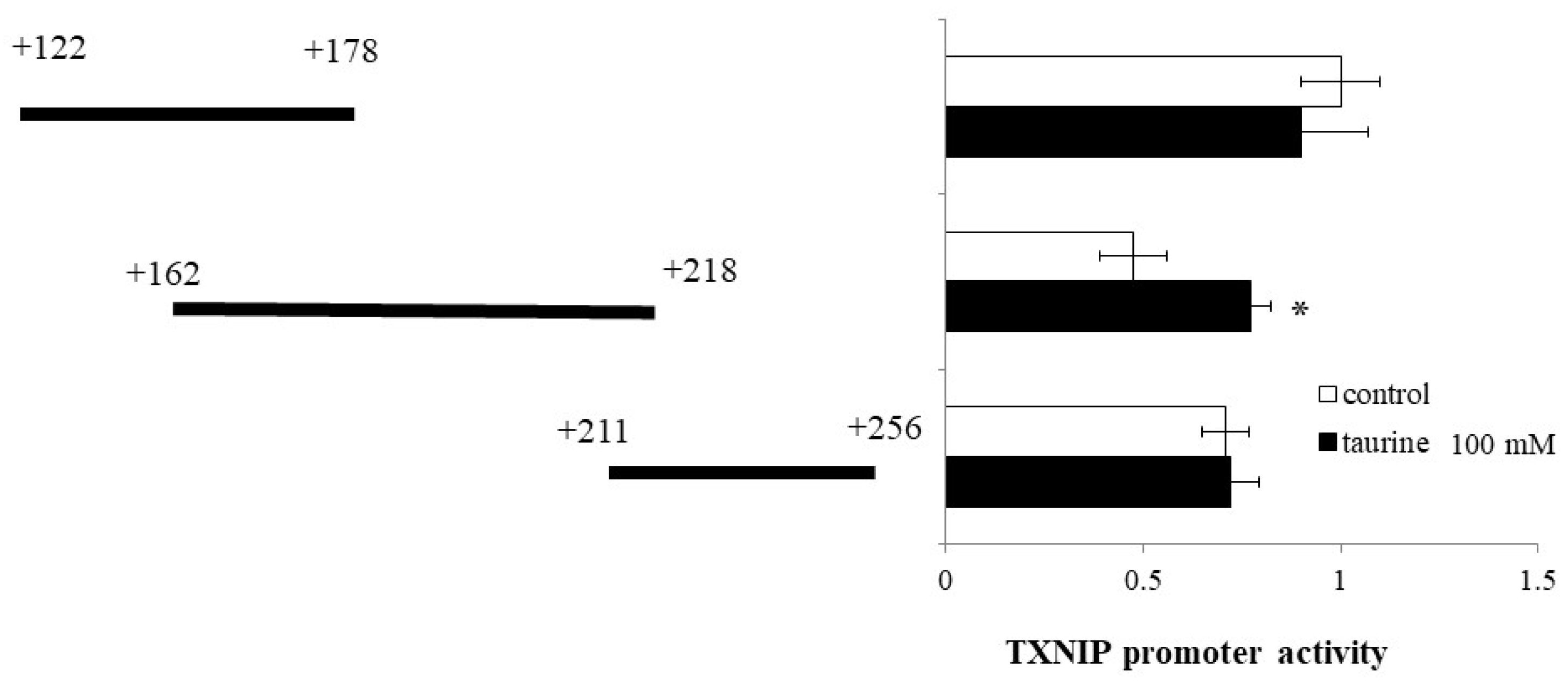
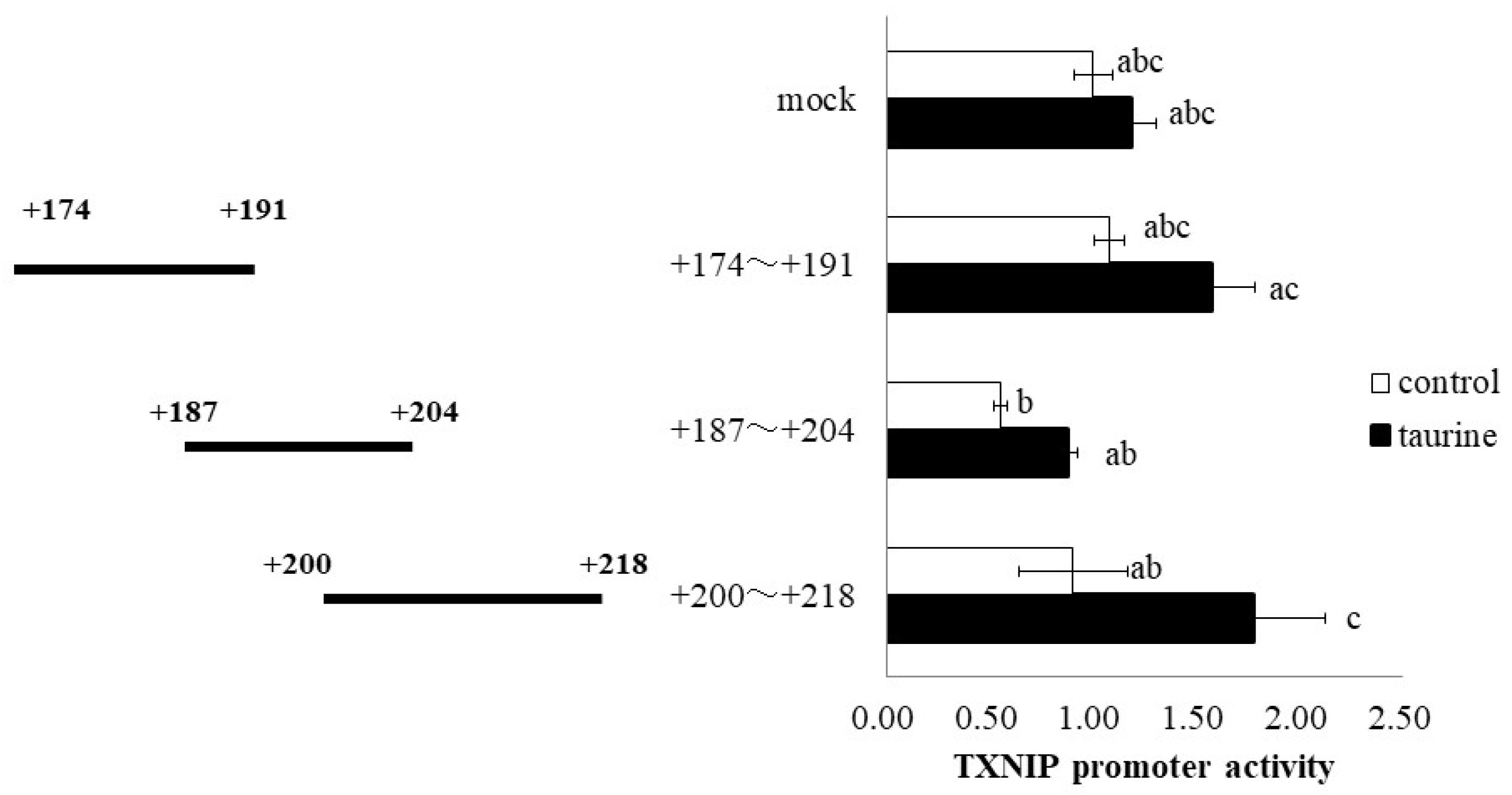
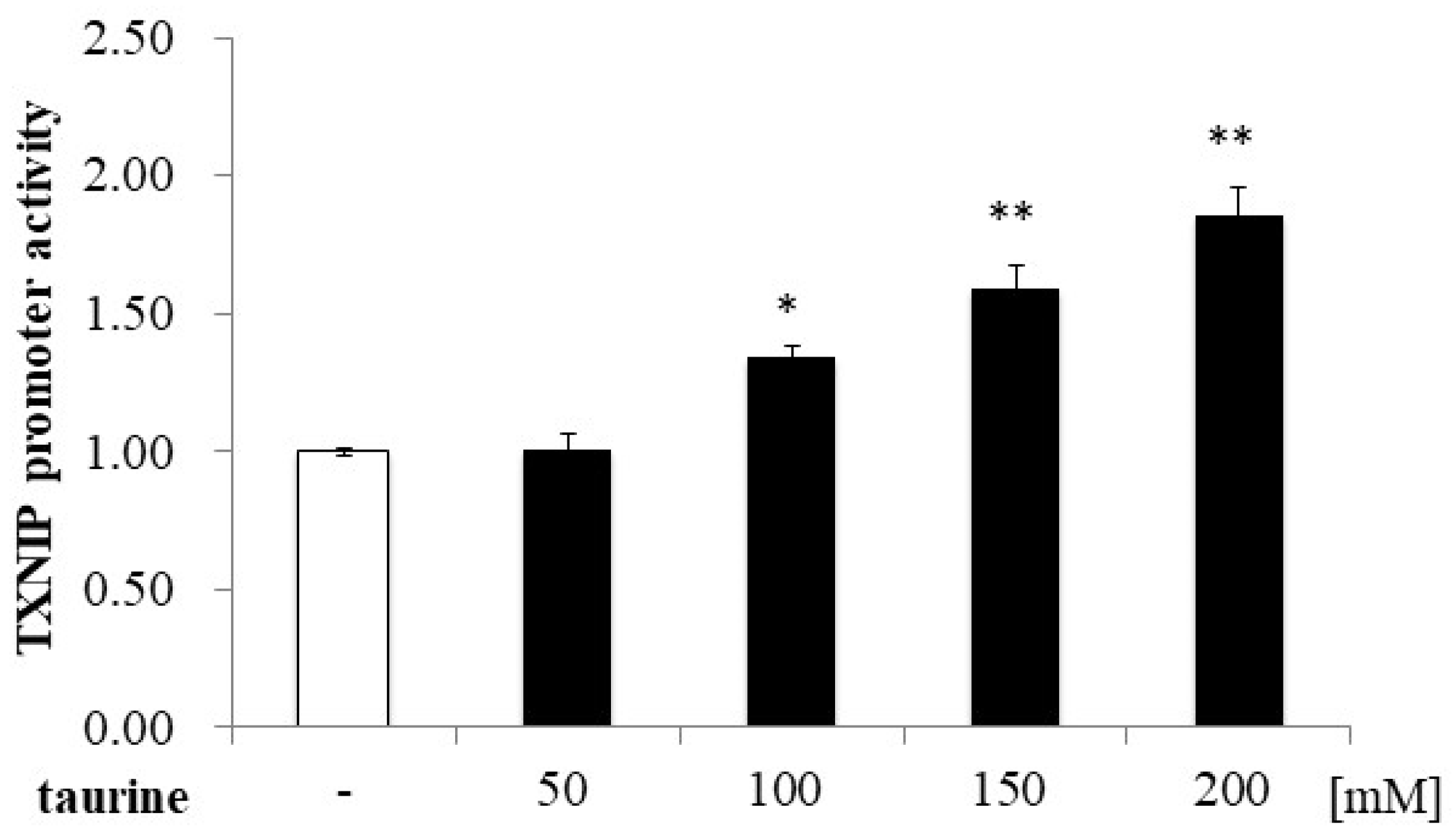
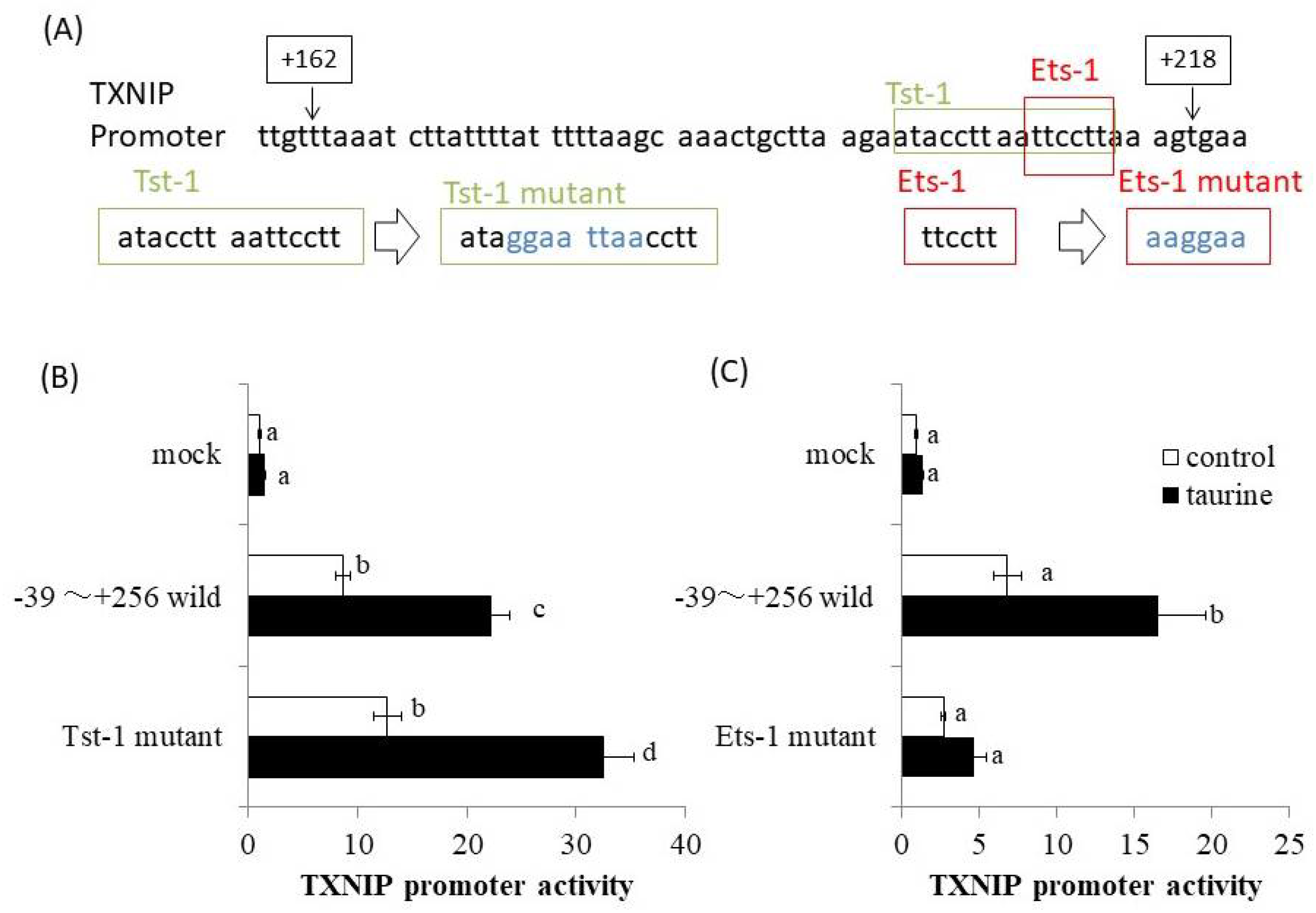
2.1. Effect of Taurine on Luciferase Activity Involving the TXNIP Promoter Region
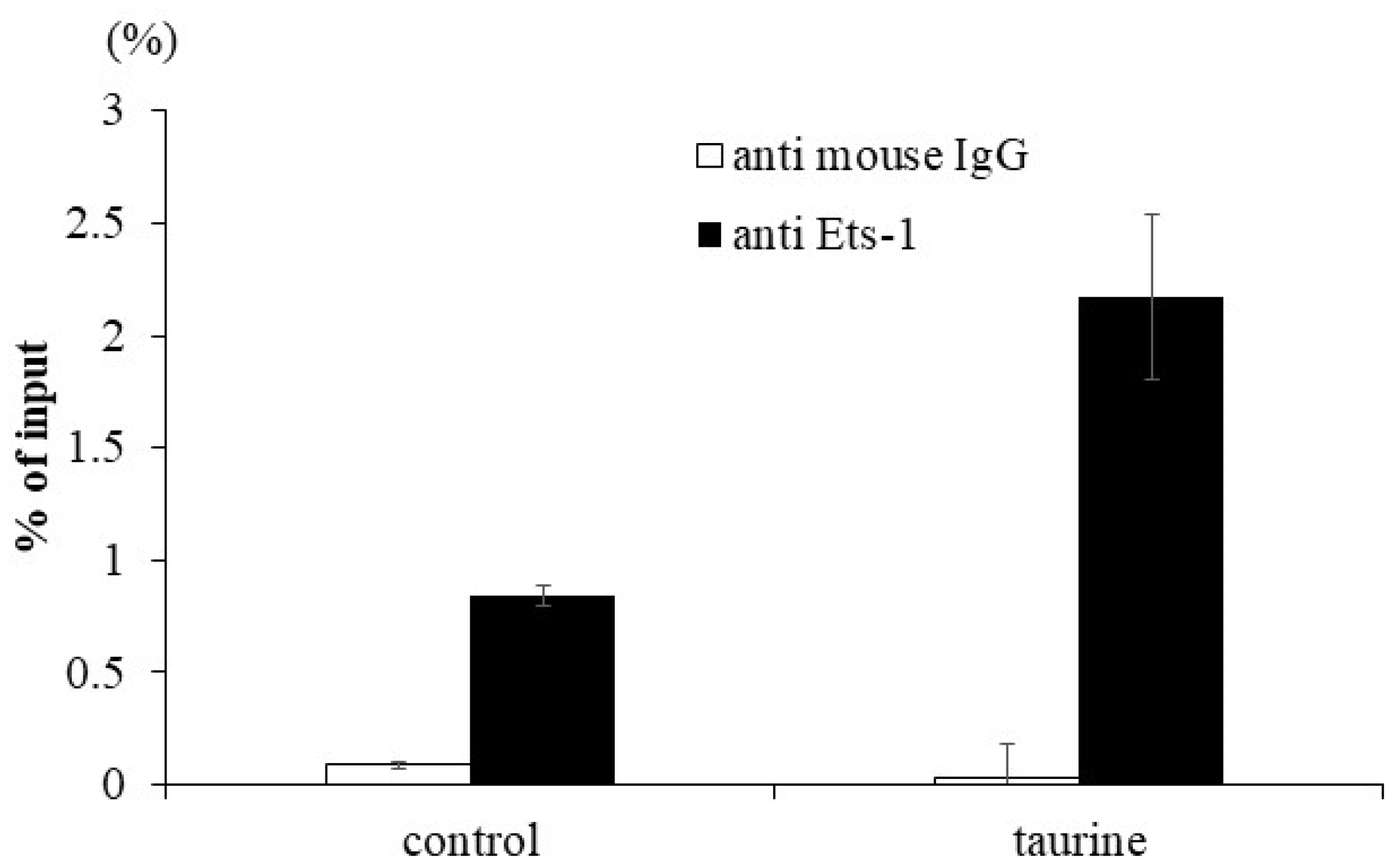
2.2. Effect of Taurine on Ets-1 Binding to TXNIP Promoter (ChIP Assay)
2.3. Effect of Taurine on Ets-1 Activation (Phosphorylation)
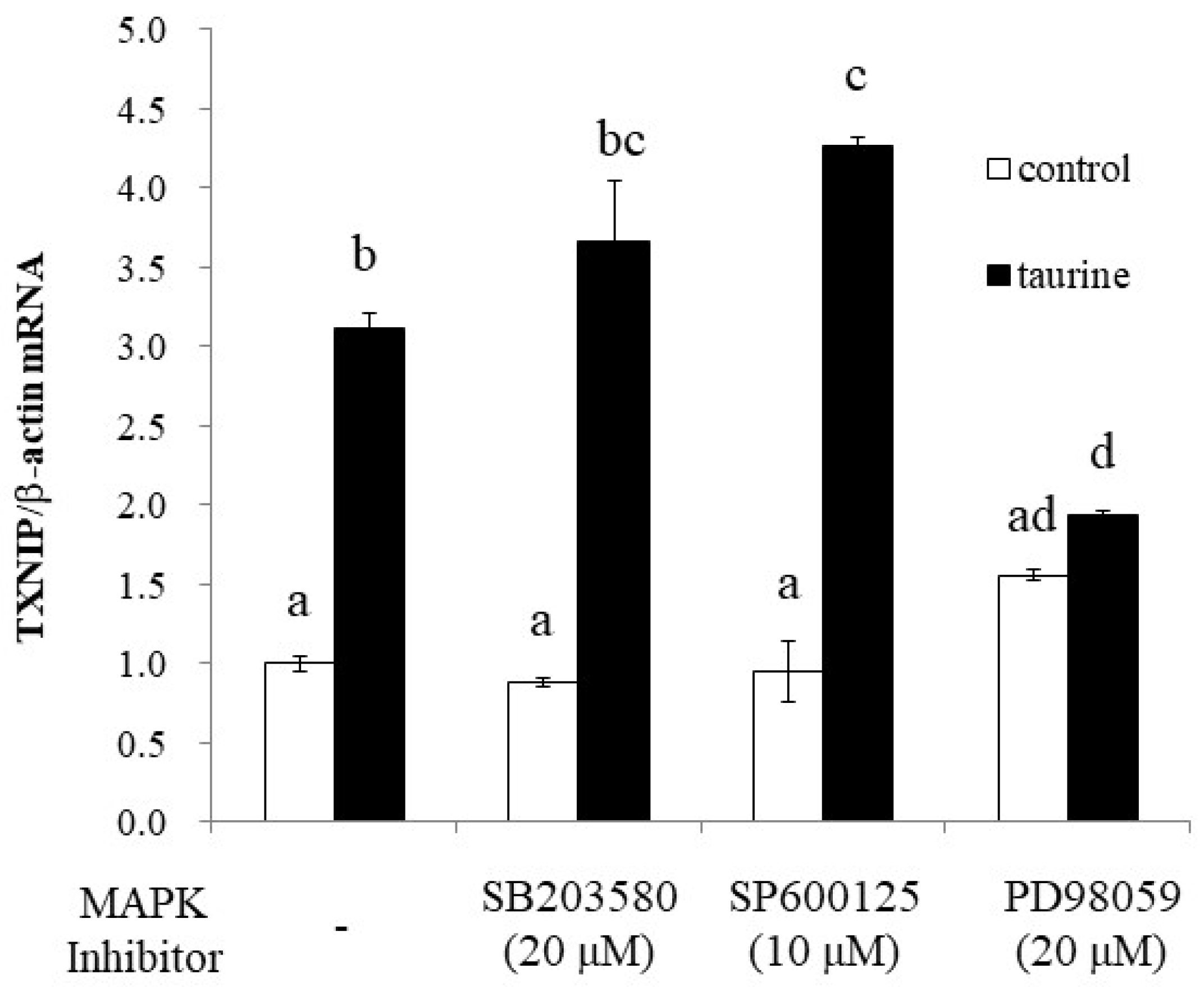
2.4. The Involvement of MAP Kinase Family on Taurine-Induced Induction of TXNIP mRNA
2.5. Effect of Taurine on ERK1/2 Activation in Caco-2 Cells
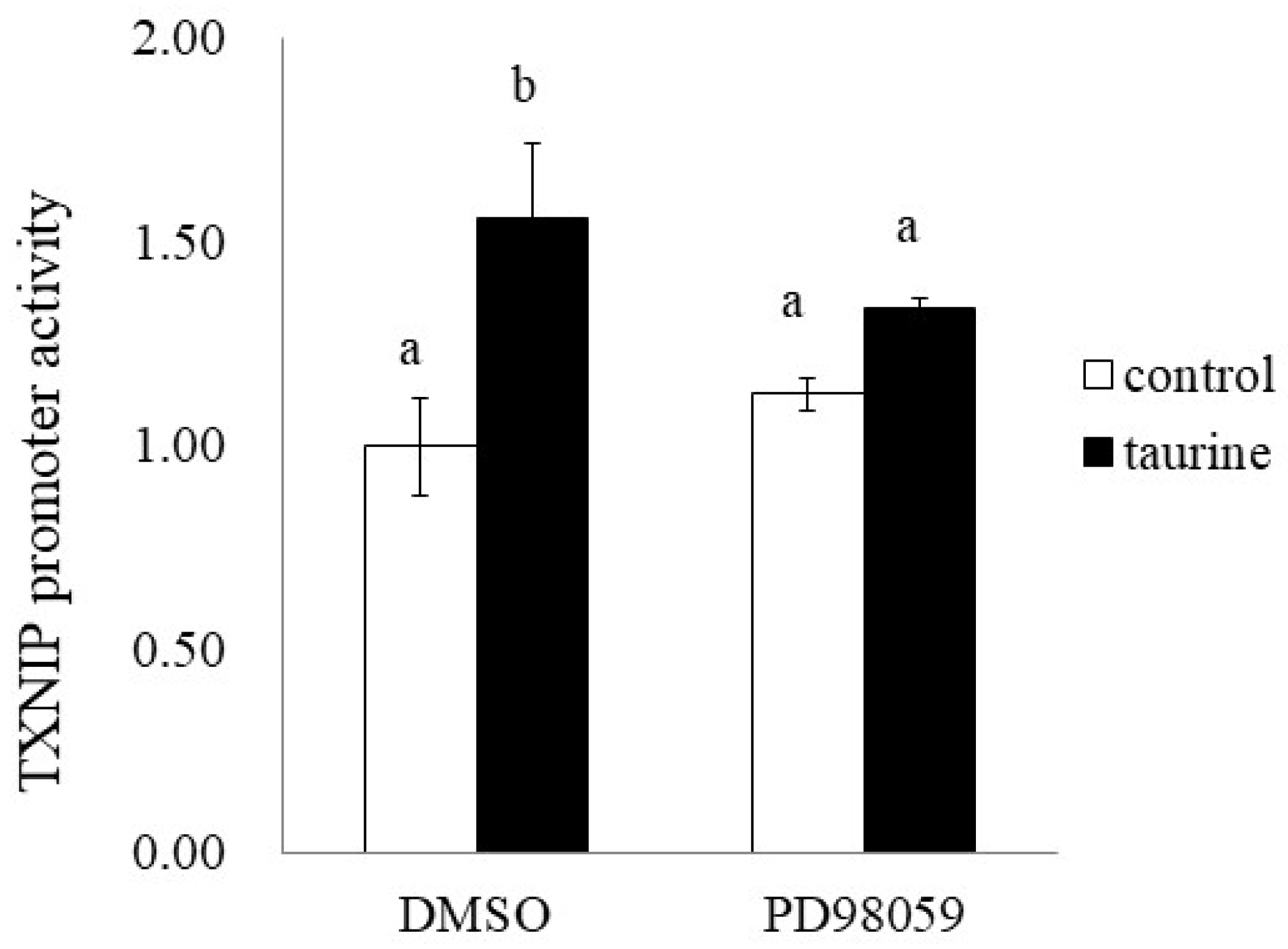
2.6. Involvement of ERK Signaling Pathway in Taurine-Induced Enhancement of Transcriptional Activity of TXNIP
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Plasmid Construct
4.4. Transfection and Reporter Assay
4.5. Real-Time PCR Analysis
4.6. Chip Assay
4.7. Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Primer Sequences (5′→3′) | |
|---|---|---|
| −1299/+256 | forward | CCGGTACCCCAACAAAGAATGAAGAGAGAG |
| reverse | GCAAGCTTCTCCAAATCGAGGAAACCC | |
| −109/+256 | forward | CCGGTACCAGCCAATGGGAGGGATG |
| reverse | GCAAGCTTCTCCAAATCGAGGAAACCC | |
| −39/+256 | forward | CCGGTACCCGGGCTACTATATAGAGACG |
| reverse | GCAAGCTTCTCCAAATCGAGGAAACCC | |
| −39/+142 | forward | CCGGTACCCGGGCTACTATATAGAGACG |
| reverse | GCAAGCTTCTAGGTTTTCGAAAAGGCGCC | |
| −39/+65 | forward | CCGGTACCCGGGCTACTATATAGAGACG |
| reverse | GCAAGCTTCCCCAATTGCTGGAGAAAAG | |
| −1299/+142 | forward | CCGGTACCCCAACAAAGAATGAAGAGAGAG |
| reverse | GCAAGCTTCTAGGTTTTCGAAAAGGCGCC | |
| +122/+256 | forward | CCGGTACCGGCGCCTTTTCGAAAACCTAG |
| reverse | GCAAGCTTCTCCAAATCGAGGAAACCC |
| Region | Primer Sequences (5′→3′) | |
|---|---|---|
| +122/+178 | forward | cggcgccttttcgaaaacctagtagttaatattcatttgtttaaatcttattttata |
| reverse | agcttataaaataagatttaaacaaatgaatattaactactaggttttcgaaaaggcgccggtac | |
| +162/+218 | forward | cttaaatcttattttatttttaagctcaaactgcttaagaataccttaattccttaaaga |
| reverse | agcttctttaaggaattaaggtattcttaagcagtttgagcttaaaaataaaataagatttaaggtac | |
| +211/+256 | forward | cccttaaagtgaaataattttttgcaaaggggtttcctcgatttggaga |
| reverse | agcttctccaaatcgaggaaacccctttgcaaaaaattatttcactttaaggggtac |
| Region | Primer Sequences (5′→3′) | |
|---|---|---|
| +174/+191 | forward | cttatttttaagctcaaaca |
| reverse | agcttgtttgagcttaaaaataaggtac | |
| +187/+204 | forward | ccaaactgcttaagaataca |
| reverse | agcttgtattcttaagcagtttgggtac | |
| +200/+218 | forward | caataccttaattccttaaaa |
| reverse | agctttttaaggaattaaggtattggtac |

References
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffer, S.W.; Azuma, J.; Takahashi, K.; Mozaffari, M. Why is taurine cytoprotective? Adv. Exp. Med. Biol. 2003, 526, 307–321. [Google Scholar] [PubMed]
- Sturman, J.A. Taurine in development. J. Nutr. 1988, 118, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Tochitani, S. Functions of Maternally-Derived Taurine in Fetal and Neonatal Brain Development. Adv. Exp. Med. Biol. 2019, 975, 17–25. [Google Scholar]
- Satsu, H.; Watanabe, H.; Arai, S.; Shimizu, M. Characterization and regulation of taurine transport in Caco-2, human intestinal cells. J. Biochem. 1997, 121, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Satsu, H.; Miyamoto, Y.; Shimizu, M. Hypertonicity stimulates taurine uptake and transporter gene expression in Caco-2 cells. Biochim. Biophys. Acta 1999, 1419, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, T.; Satsu, H.; Shimizu, M. Tumor necrosis factor alpha stimulates taurine uptake and transporter gene expression in human intestinal Caco-2 cells. FEBS Lett. 2002, 517, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Satsu, H.; Fujisawa, M.; Hori, M.; Ishimoto, Y.; Totsuka, M.; Nambu, A.; Kakuta, S.; Ozaki, H.; Shimizu, M. Attenuation by dietary taurine of dextran sulfate sodium-induced colitis in mice and of THP-1-induced damage to intestinal Caco-2 cell monolayers. Amino Acids 2008, 35, 217–224. [Google Scholar] [CrossRef]
- Gondo, Y.; Satsu, H.; Ishimoto, Y.; Iwamoto, T.; Shimizu, M. Effect of taurine on mRNA expression of thioredoxin interacting protein in Caco-2 cells. Biochem. Biophys. Res. Commun. 2012, 426, 433–437. [Google Scholar] [CrossRef]
- Kaimul, A.M.; Nakamura, H.; Masutani, H.; Yodoi, J. Thioredoxin and thioredoxin-binding protein-2 in cancer and metabolic syndrome. Free Radic. Biol. Med. 2007, 43, 861–868. [Google Scholar] [CrossRef]
- Kim, S.Y.; Suh, H.W.; Chung, J.W.; Yoon, S.R.; Choi, I. Diverse functions of VDUP1 in cell proliferation, differentiation, and diseases. Cell Mol. Immunol. 2007, 4, 345–351. [Google Scholar] [PubMed]
- Andres, A.M.; Ratliff, E.P.; Sachithanantham, S.; Hui, S.T. Diminished AMPK signaling response to fasting in thioredoxin-interacting protein knockout mice. FEBS Lett. 2011, 585, 1223–1230. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, K.L.; Margosian, M.R.; Sheth, S.S.; Lusis, A.J.; Parks, E.J. Increased lipogenesis and fatty acid reesterification contribute to hepatic triacylglycerol stores in hyperlipidemic Txnip-/- mice. J. Nutr. 2004, 134, 1475–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, S.T.; Andres, A.M.; Miller, A.K.; Spann, N.J.; Potter, D.W.; Post, N.M.; Chen, A.Z.; Sachithanantham, S.; Jung, D.Y.; Kim, J.K.; et al. Txnip balances metabolic and growth signaling via PTEN disulfide reduction. Proc. Natl. Acad. Sci. USA 2008, 105, 3921–3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, S.; Liu, W.; Masutani, H.; Hirata, H.; Shinkai, Y.; Yamada, S.; Yoshida, T.; Nakamura, H.; Yodoi, J. Impaired fatty acid utilization in thioredoxin binding protein-2 (TBP-2)-deficient mice: A unique animal model of Reye syndrome. FASEB J. 2006, 20, 121–123. [Google Scholar] [CrossRef]
- Sheth, S.S.; Castellani, L.W.; Chari, S.; Wagg, C.; Thipphavong, C.K.; Bodnar, J.S.; Tontonoz, P.; Attie, A.D.; Lopaschuk, G.D.; Lusis, A.J. Thioredoxin-interacting protein deficiency disrupts the fasting-feeding metabolic transition. J. Lipid Res. 2005, 46, 123–134. [Google Scholar] [CrossRef] [Green Version]
- van Greevenbroek, M.M.; Vermeulen, V.M.; Feskens, E.J.; Evelo, C.T.; Kruijshoop, M.; Hoebee, B.; van der Kallen, C.J.; de Bruin, T.W. Genetic variation in thioredoxin interacting protein (TXNIP) is associated with hypertriglyceridaemia and blood pressure in diabetes mellitus. Diabet. Med. 2007, 24, 498–504. [Google Scholar] [CrossRef]
- Chutkow, W.A.; Patwari, P.; Yoshioka, J.; Lee, R.T. Thioredoxin-interacting protein (Txnip) is a critical regulator of hepatic glucose production. J. Biol. Chem. 2008, 283, 2397–2406. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, N.E.; Omae, S.; Pereira, A.; Rodrigues, M.V.; Miyakawa, A.A.; Campos, L.C.; Santos, P.C.; Dallan, L.A.; Martinez, T.L.; Santos, R.D.; et al. Thioredoxin interacting protein genetic variation is associated with diabetes and hypertension in the Brazilian general population. Atherosclerosis 2012, 221, 131–136. [Google Scholar] [CrossRef]
- Yoshioka, J.; Schulze, P.C.; Cupesi, M.; Sylvan, J.D.; MacGillivray, C.; Gannon, J.; Huang, H.; Lee, R.T. Thioredoxin-interacting protein controls cardiac hypertrophy through regulation of thioredoxin activity. Circulation 2004, 109, 2581–2586. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Masuda, H.; Ishii, Y.; Nishida, Y.; Kobayashi, M.; Asai, S. Decreased expression of thioredoxin interacting protein mRNA in inflamed colonic mucosa in patients with ulcerative colitis. Oncol. Rep. 2007, 18, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Satsu, H.; Gondo, Y.; Shimanaka, H.; Watari, K.; Fukumura, M.; Shimizu, M. Effect of taurine on cell function via TXNIP induction in Caco-2 cells. Adv. Exp. Med. Biol. 2019, 1155, 163–172. [Google Scholar] [PubMed]
- Garrett-Sinha, L.A. Review of Ets1 structure, function, and roles in immunity. Cell. Mol. Life Sci. 2013, 70, 3375–3390. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Cook, T.J.; Sinko, P.J. Evidence for diminished functional expression of intestinal transporters in Caco-2 cell monolayers at high passages. Pharm. Res. 1997, 14, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Steffansen, B.; Pedersen, M.D.L.; Laghmoch, A.M.; Nielsen, C.U. SGLT1-Mediated Transport in Caco-2 Cells Is Highly Dependent on Cell Bank Origin. J. Pharm. Sci. 2017, 106, 2664–2670. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Gerrero, R.; Simmons, D.M.; Park, R.E.; Lin, C.J.; Swanson, L.W.; Rosenfeld, M.G. Tst-1, a member of the POU domain gene family, binds the promoter of the gene encoding the cell surface adhesion molecule P0. Mol. Cell. Biol. 1991, 11, 1739–1744. [Google Scholar]
- Yang, X.; McDonough, J.; Fyodorov, D.; Morris, M.; Wang, F.; Deneris, E.S. Characterization of an acetylcholine receptor alpha 3 gene promoter and its activation by the POU domain factor SCIP/Tst-1. J. Biol. Chem. 1994, 269, 10252–10264. [Google Scholar] [CrossRef]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Sementchenko, V.I.; Watson, D.K. Ets target genes: Past, present and future. Oncogene 2000, 19, 6533–6548. [Google Scholar] [CrossRef] [Green Version]
- Dittmer, J. The biology of the Ets1 proto-oncogene. Mol. Cancer 2003, 2, 29. [Google Scholar] [CrossRef] [Green Version]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; He, F.; Hu, L.; Hai, L.; Huang, M.; Xu, Z.; Zhang, J.; Zhou, Z.; Liu, F.; Dai, Y.S. Transcription factor Ets1 regulates expression of thioredoxin-interacting protein and inhibits insulin secretion in pancreatic beta-cells. PLoS ONE 2014, 9, e99049. [Google Scholar]
- Fiorito, V.; Neri, F.; Pala, V.; Silengo, L.; Oliviero, S.; Altruda, F.; Tolosano, E. Hypoxia controls Flvcr1 gene expression in Caco2 cells through HIF2alpha and ETS1. Biochim. Biophys. Acta 2014, 1839, 259–264. [Google Scholar] [CrossRef]
- Hashiguchi, K.; Tsuchiya, H.; Tomita, A.; Ueda, C.; Akechi, Y.; Sakabe, T.; Kurimasa, A.; Nozaki, M.; Yamada, T.; Tsuchida, S.; et al. Involvement of ETS1 in thioredoxin-binding protein 2 transcription induced by a synthetic retinoid CD437 in human osteosarcoma cells. Biochem. Biophys. Res. Commun. 2010, 391, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Beek, J.P.; Kennedy, L.; Rockel, J.S.; Bernier, S.M.; Leask, A. The induction of CCN2 by TGFbeta1 involves Ets-1. Arthritis Res. Ther. 2006, 8, R36. [Google Scholar] [CrossRef] [Green Version]
- Geisinger, M.T.; Astaiza, R.; Butler, T.; Popoff, S.N.; Planey, S.L.; Arnott, J.A. Ets-1 is essential for connective tissue growth factor (CTGF/CCN2) induction by TGF-beta1 in osteoblasts. PLoS ONE 2012, 7, e35258. [Google Scholar] [CrossRef] [Green Version]
- Yoshimatsu, Y.; Yamazaki, T.; Mihira, H.; Itoh, T.; Suehiro, J.; Yuki, K.; Harada, K.; Morikawa, M.; Iwata, C.; Minami, T.; et al. Ets family members induce lymphangiogenesis through physical and functional interaction with Prox1. J. Cell Sci. 2011, 124, 2753–2762. [Google Scholar] [CrossRef] [Green Version]
- Itoh, T.; Ando, M.; Tsukamasa, Y.; Akao, Y. Expression of BMP-2 and Ets1 in BMP-2-stimulated mouse pre-osteoblast differentiation is regulated by microRNA-370. FEBS Lett. 2012, 586, 1693–1701. [Google Scholar] [CrossRef]
- Ohtani, N.; Zebedee, Z.; Huot, T.J.; Stinson, J.A.; Sugimoto, M.; Ohashi, Y.; Sharrocks, A.D.; Peters, G.; Hara, E. Opposing effects of Ets and Id proteins on p16INK4a expression during cellular senescence. Nature 2001, 409, 1067–1070. [Google Scholar] [CrossRef]
- Kola, I.; Brookes, S.; Green, A.R.; Garber, R.; Tymms, M.; Papas, T.S.; Seth, A. The Ets1 transcription factor is widely expressed during murine embryo development and is associated with mesodermal cells involved in morphogenetic processes such as organ formation. Proc. Natl. Acad. Sci. USA 1993, 90, 7588–7592. [Google Scholar] [CrossRef] [Green Version]
- Wei, G.; Srinivasan, R.; Cantemir-Stone, C.Z.; Sharma, S.M.; Santhanam, R.; Weinstein, M.; Muthusamy, N.; Man, A.K.; Oshima, R.G.; Leone, G.; et al. Ets1 and Ets2 are required for endothelial cell survival during embryonic angiogenesis. Blood 2009, 114, 1123–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, K.; Muthusamy, N.; Fischer, C.; Ting, C.N.; Walunas, T.L.; Lanier, L.L.; Leiden, J.M. The Ets-1 transcription factor is required for the development of natural killer cells in mice. Immunity 1998, 9, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Harrisingh, M.C.; Lloyd, A.C. Ras/Raf/ERK signalling and NF1. Cell Cycle 2004, 3, 1255–1258. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, J.; Li, L.; Liu, L.; Wu, L. Role of Ras/PKCzeta/MEK/ERK1/2 signaling pathway in angiotensin II-induced vascular smooth muscle cell proliferation. Regul. Pept. 2005, 128, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Crawley, S.; Hokari, R.; Kwon, S.; Kim, Y.S. Bile acid regulates MUC2 transcription in colon cancer cells via positive EGFR/PKC/Ras/ERK/CREB, PI3K/Akt/IkappaB/NF-kappaB and p38/MSK1/CREB pathways and negative JNK/c-Jun/AP-1 pathway. Int. J. Oncol. 2010, 36, 941–953. [Google Scholar]
- Wen-Sheng, W. Protein kinase C alpha trigger Ras and Raf-independent MEK/ERK activation for TPA-induced growth inhibition of human hepatoma cell HepG2. Cancer Lett. 2006, 239, 27–35. [Google Scholar] [CrossRef]
- Loo, D.D.; Hirsch, J.R.; Sarkar, H.K.; Wright, E.M. Regulation of the mouse retinal taurine transporter (TAUT) by protein kinases in Xenopus oocytes. FEBS Lett. 1996, 392, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.S.; Hauser, C.A.; Henkel, G.; Colman, M.S.; Van Beveren, C.; Stacey, K.J.; Hume, D.A.; Maki, R.A.; Ostrowski, M.C. Ras-mediated phosphorylation of a conserved threonine residue enhances the transactivation activities of c-Ets1 and c-Ets2. Mol. Cell. Biol. 1996, 16, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Huang, L.; Gest, C.; Xi, X.; Janin, A.; Soria, C.; Li, H.; Lu, H. Opposite regulation by PI3K/Akt and MAPK/ERK pathways of tissue factor expression, cell-associated procoagulant activity and invasiveness in MDA-MB-231 cells. J. Hematol. Oncol. 2012, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Taber, K.H.; Lin, C.T.; Liu, J.W.; Thalmann, R.H.; Wu, J.Y. Taurine in hippocampus: Localization and postsynaptic action. Brain Res. 1986, 386, 113–121. [Google Scholar] [CrossRef]
- del Olmo, N.; Bustamante, J.; del Río, R.M.; Solís, J.M. Taurine activates GABA(A) but not GABA(B) receptors in rat hippocampal CA1 area. Brain Res. 2000, 864, 298–307. [Google Scholar] [CrossRef]
- Hussy, N.; Deleuze, C.; Pantaloni, A.; Desarménien, M.G.; Moos, F. Agonist action of taurine on glycine receptors in rat supraoptic magnocellular neurones: Possible role in osmoregulation. J. Physiol. 1997, 502, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.A.; Ahmed, Z.; Faber, D.S. A characterization of glycinergic receptors present in cultured rat medullary neurons. J. Neurophysiol. 1991, 66, 1291–1303. [Google Scholar] [CrossRef] [PubMed]
- Whatley, V.J.; Harris, R.A. The cytoskeleton and neurotransmitter receptors. Int. Rev. Neurobiol. 1996, 39, 113–143. [Google Scholar] [PubMed]
- Horikoshi, T.; Asanuma, A.; Yanagisawa, K.; Anzai, K.; Goto, S. Taurine and beta-alanine act on both GABA and glycine receptors in Xenopus oocyte injected with mouse brain messenger RNA. Brain Res. 1988, 464, 97–105. [Google Scholar] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satsu, H.; Gondo, Y.; Shimanaka, H.; Imae, M.; Murakami, S.; Watari, K.; Wakabayashi, S.; Park, S.-J.; Nakai, K.; Shimizu, M. Signaling Pathway of Taurine-Induced Upregulation of TXNIP. Metabolites 2022, 12, 636. https://doi.org/10.3390/metabo12070636
Satsu H, Gondo Y, Shimanaka H, Imae M, Murakami S, Watari K, Wakabayashi S, Park S-J, Nakai K, Shimizu M. Signaling Pathway of Taurine-Induced Upregulation of TXNIP. Metabolites. 2022; 12(7):636. https://doi.org/10.3390/metabo12070636
Chicago/Turabian StyleSatsu, Hideo, Yusuke Gondo, Hana Shimanaka, Masato Imae, Shigeru Murakami, Kenji Watari, Shunichi Wakabayashi, Sung-Joon Park, Kenta Nakai, and Makoto Shimizu. 2022. "Signaling Pathway of Taurine-Induced Upregulation of TXNIP" Metabolites 12, no. 7: 636. https://doi.org/10.3390/metabo12070636
APA StyleSatsu, H., Gondo, Y., Shimanaka, H., Imae, M., Murakami, S., Watari, K., Wakabayashi, S., Park, S.-J., Nakai, K., & Shimizu, M. (2022). Signaling Pathway of Taurine-Induced Upregulation of TXNIP. Metabolites, 12(7), 636. https://doi.org/10.3390/metabo12070636