
Impacts of Formula Supplemented with Milk Fat Globule Membrane on the Neurolipidome of Brain Regions of Piglets

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
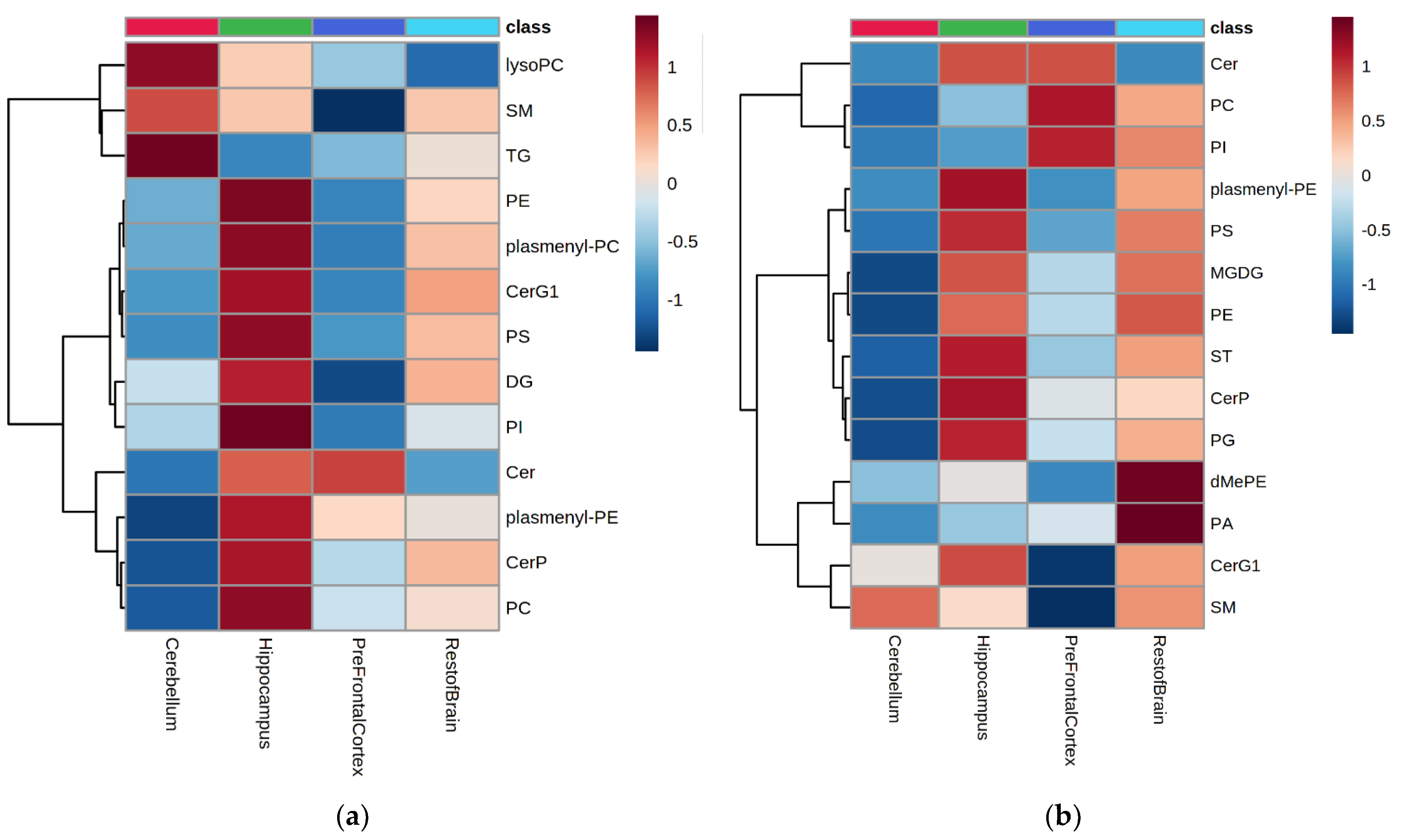
2.1. Brain Region Lipidome
2.1.1. Lipid Annotation and Identification
2.1.2. Lipid Class Composition
2.1.3. Lipid Species Composition
2.2. Dietary Intervention Impact on Brain Lipidome
2.2.1. Lipid Class-Level Analysis
2.2.2. Lipid Species-Level Analysis
2.2.3. Plasma Metabolomic and Lipidomic Analyses
3. Discussion
4. Materials and Methods
4.1. Animal Trial and Sampling
4.2. Brain and Plasma Extractions
4.3. LC-MS Lipidomics
4.4. LC-MS Metabolomics
4.5. Data Extraction and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, H.; Villanueva, N.; Bittar, T.; Arsenault, E.; Labonte, B.; Huan, T. Parallel metabolomics and lipidomics enables the comprehensive study of mouse brain regional metabolite and lipid patterns. Anal. Chim. Acta 2020, 1136, 168–177. [Google Scholar] [CrossRef]
- Dawson, G. Measuring brain lipids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 1026–1039. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, S.; Liang, X.; Zhang, H. Development of a mass-spectrometry-based lipidomics platform for the profiling of phospholipids and sphingolipids in brain tissues. Anal. Bioanal. Chem. 2015, 407, 6543–6555. [Google Scholar] [CrossRef]
- Lee, J.C.; Park, S.M.; Kim, I.Y.; Sung, H.; Seong, J.K.; Moon, M.H. High-fat diet-induced lipidome perturbations in the cortex, hippocampus, hypothalamus, and olfactory bulb of mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 980–990. [Google Scholar] [CrossRef]
- Fitzner, D.; Bader, J.M.; Penkert, H.; Bergner, C.G.; Su, M.; Weil, M.-T.; Surma, M.A.; Mann, M.; Klose, C.; Simons, M. Cell-Type- and Brain-Region-Resolved Mouse Brain Lipidome. Cell Rep. 2020, 32, 108132. [Google Scholar] [CrossRef]
- Giles, C.; Takechi, R.; Mellett, N.A.; Meikle, P.J.; Dhaliwal, S.; Mamo, J.C. The Effects of Long-Term Saturated Fat Enriched Diets on the Brain Lipidome. PLoS ONE 2016, 11, e0166964. [Google Scholar] [CrossRef]
- Chan, R.B.; Oliveira, T.G.; Cortes, E.P.; Honig, L.S.; Duff, K.E.; Small, S.A.; Wenk, M.R.; Shui, G.; Di Paolo, G. Comparative Lipidomic Analysis of Mouse and Human Brain with Alzheimer Disease. J. Biol. Chem. 2012, 287, 2678–2688. [Google Scholar] [CrossRef]
- Brink, L.R.; Lönnerdal, B. The role of milk fat globule membranes in behavior and cognitive function using a suckling rat pup supplementation model. J. Nutr. Biochem. 2018, 58, 131–137. [Google Scholar] [CrossRef]
- Hahn, K.; Hardimon, J.R.; Caskey, D.; Jost, D.A.; Roady, P.J.; Brenna, J.T.; Dilger, R.N. Safety and Efficacy of Sodium and Potassium Arachidonic Acid Salts in the Young Pig. Nutrients 2021, 13, 1482. [Google Scholar] [CrossRef]
- Hahn, K.E.; Dahms, I.; Butt, C.M.; Salem, N.; Grimshaw, V.; Bailey, E.; Fleming, S.A.; Smith, B.N.; Dilger, R.N. Impact of Arachidonic and Docosahexaenoic Acid Supplementation on Neural and Immune Development in the Young Pig. Front. Nutr. 2020, 7, 592364. [Google Scholar] [CrossRef]
- Lee, H.; Padhi, E.; Hasegawa, Y.; Larke, J.; Parenti, M.; Wang, A.; Hernell, O.; Lönnerdal, B.; Slupsky, C. Compositional Dynamics of the Milk Fat Globule and Its Role in Infant Development. Front. Pediatr. 2018, 6, 313. [Google Scholar] [CrossRef]
- Cilla, A.; Diego Quintaes, K.; Barberá, R.; Alegría, A. Phospholipids in Human Milk and Infant Formulas: Benefits and Needs for Correct Infant Nutrition. Crit. Rev. Food Sci. Nutr. 2016, 56, 1880–1892. [Google Scholar] [CrossRef]
- Gurnida, D.A.; Rowan, A.M.; Idjradinata, P.; Muchtadi, D.; Sekarwana, N. Association of complex lipids containing gangliosides with cognitive development of 6-month-old infants. Early Hum. Dev. 2012, 88, 595–601. [Google Scholar] [CrossRef]
- Ryan, J.M.; Rice, G.E.; Mitchell, M.D. The role of gangliosides in brain development and the potential benefits of perinatal supplementation. Nutr. Res. 2013, 33, 877–887. [Google Scholar] [CrossRef]
- Zou, L.; Pande, G.; Akoh, C.C. Infant Formula Fat Analogs and Human Milk Fat: New Focus on Infant Developmental Needs. Annu. Rev. Food Sci. Technol. 2016, 7, 139–165. [Google Scholar] [CrossRef]
- He, X.; Parenti, M.; Grip, T.; Lonnerdal, B.; Timby, N.; Domellof, M.; Hernell, O.; Slupsky, C.M. Fecal microbiome and metabolome of infants fed bovine MFGM supplemented formula or standard formula with breast-fed infants as reference: A randomized controlled trial. Sci. Rep. 2019, 9, 11589. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, L.; Tian, S.; Li, X.; Hussain, M.; Li, C.; Zhang, L.; Zhang, Q.; Leng, Y.; Jiang, S.; et al. Comparative analysis of interfacial composition and structure of fat globules in human milk and infant formulas. Food Hydrocoll. 2022, 124, 107290. [Google Scholar] [CrossRef]
- Veereman-Wauters, G.; Staelens, S.; Rombaut, R.; Dewettinck, K.; Deboutte, D.; Brummer, R.-J.; Boone, M.; Le Ruyet, P. Milk fat globule membrane (INPULSE) enriched formula milk decreases febrile episodes and may improve behavioral regulation in young children. Nutrition 2012, 28, 749–752. [Google Scholar] [CrossRef]
- Timby, N.; Domellöf, E.; Hernell, O.; Lönnerdal, B.; Domellöf, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef]
- Mudd, A.T.; Alexander, L.S.; Berding, K.; Waworuntu, R.V.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Prebiotics, Milk Fat Globule Membrane, and Lactoferrin Affects Structural Neurodevelopment in the Young Piglet. Front. Pediatr. 2016, 4, 4. [Google Scholar] [CrossRef]
- Fil, J.E.; Fleming, S.A.; Chichlowski, M.; Gross, G.; Berg, B.M.; Dilger, R.N. Evaluation of Dietary Bovine Milk Fat Globule Membrane Supplementation on Growth, Serum Cholesterol and Lipoproteins, and Neurodevelopment in the Young Pig. Front. Pediatr. 2019, 7, 417. [Google Scholar] [CrossRef]
- Thum, C.; Wall, C.; Day, L.; Szeto, I.M.Y.; Li, F.; Yan, Y.; Barnett, M.P.G. Changes in Human Milk Fat Globule Composition Throughout Lactation: A Review. Front. Nutr. 2022, 9, 835856. [Google Scholar] [CrossRef] [PubMed]
- Tracey, T.J.; Steyn, F.J.; Wolvetang, E.J.; Ngo, S.T. Neuronal Lipid Metabolism: Multiple Pathways Driving Functional Outcomes in Health and Disease. Front. Mol. Neurosci. 2018, 11, 10. [Google Scholar] [CrossRef]
- Cifkova, E.; Holcapek, M.; Lisa, M. Nontargeted lipidomic characterization of porcine organs using hydrophilic interaction liquid chromatography and off-line two-dimensional liquid chromatography-electrospray ionization mass spectrometry. Lipids 2013, 48, 915–928. [Google Scholar] [CrossRef]
- Müller, C.P.; Reichel, M.; Mühle, C.; Rhein, C.; Gulbins, E.; Kornhuber, J. Brain membrane lipids in major depression and anxiety disorders. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 1052–1065. [Google Scholar] [CrossRef] [PubMed]
- Aidoud, N.; Delplanque, B.; Baudry, C.; Garcia, C.; Moyon, A.; Balasse, L.; Guillet, B.; Antona, C.; Darmaun, D.; Fraser, K.; et al. A combination of lipidomics, MS imaging, and PET scan imaging reveals differences in cerebral activity in rat pups according to the lipid quality of infant formulas. FASEB J. 2018, 32, 4776–4790. [Google Scholar] [CrossRef]
- Bascoul-Colombo, C.; Guschina, I.A.; Maskrey, B.H.; Good, M.; O’Donnell, V.B.; Harwood, J.L. Dietary DHA supplementation causes selective changes in phospholipids from different brain regions in both wild type mice and the Tg2576 mouse model of Alzheimer’s disease. Biochim. Biophys. Acta 2016, 1861, 524–537. [Google Scholar] [CrossRef]
- Brink, L.R.; Herren, A.W.; McMillen, S.; Fraser, K.; Agnew, M.; Roy, N.; Lonnerdal, B. Omics analysis reveals variations among commercial sources of bovine milk fat globule membrane. J. Dairy Sci. 2020, 103, 3002–3016. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M. Phospholipids in Milk Fat: Composition, Biological and Technological Significance, and Analytical Strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Char, D.; Sheard, N.F. Choline, Phosphatidylcholine and Sphingomyelin in Human and Bovine Milk and Infant Formulas. J. Nutr. 1986, 116, 50–58. [Google Scholar] [CrossRef]
- Borg, M.L.; Omran, S.F.; Weir, J.; Meikle, P.J.; Watt, M.J. Consumption of a high-fat diet, but not regular endurance exercise training, regulates hypothalamic lipid accumulation in mice. J. Physiol. 2012, 590, 4377–4389. [Google Scholar] [CrossRef] [PubMed]
- Fil, J.E.; Joung, S.; Hayes, C.A.; Dilger, R.N. Influence of Rearing Environment on Longitudinal Brain Development, Object Recognition Memory, and Exploratory Behaviors in the Domestic Pig (Sus scrofa). Front. Neurosci. 2021, 15, 649536. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.E.; Fraser, K.; Kruger, M.C.; Sequeira, I.R.; Yip, W.; Lu, L.W.; Plank, L.D.; Murphy, R.; Cooper, G.J.S.; Martin, J.-C.; et al. Metabolomic signatures for visceral adiposity and dysglycaemia in Asian Chinese and Caucasian European adults: The cross-sectional TOFI_Asia study. Nutr. Metab. 2020, 17, 95. [Google Scholar] [CrossRef]
- Su, M.; Subbaraj, A.K.; Fraser, K.; Qi, X.; Jia, H.; Chen, W.; Gomes Reis, M.; Agnew, M.; Day, L.; Roy, N.C.; et al. Lipidomics of Brain Tissues in Rats Fed Human Milk from Chinese Mothers or Commercial Infant Formula. Metabolites 2019, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Holman, J.D.; Tabb, D.L.; Mallick, P. Employing ProteoWizard to convert raw mass spectrometry data. Curr. Protoc. Bioinform. 2014, 46, 13.24.1–13.24.9. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Giacomoni, F.; Le Corguillé, G.; Monsoor, M.; Landi, M.; Pericard, P.; Pétéra, M.; Duperier, C.; Tremblay-Franco, M.; Martin, J.-F.; Jacob, D.; et al. Workflow4Metabolomics: A collaborative research infrastructure for computational metabolomics. Bioinformatics 2014, 31, 1493–1495. [Google Scholar] [CrossRef]
- Kind, T.; Liu, K.-H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef]
- Liebisch, G.; Vizcaíno, J.A.; Köfeler, H.; Trötzmüller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Species | T Stat | Log2FC (Control/High) | p-Value |
|---|---|---|---|
| TG 53:7; [M + NH4]+ | 4.03 | 0.80 | 0.001 |
| TG 54:4; [M + NH4]+ | 3.95 | 0.65 | 0.002 |
| TG 58:3; [M + NH4]+ | 3.66 | 0.75 | 0.003 |
| TG(18:0/18:1/18:1) + NH4_MS2 | 3.55 | 0.65 | 0.004 |
| TG(16:0/18:1/18:2) + NH4_MS2 | 3.46 | 0.63 | 0.004 |
| TG 50:3; [M + NH4]+ | 3.41 | 0.70 | 0.005 |
| TG 53:6; [M + NH4]+ | 3.34 | 0.67 | 0.005 |
| TG(18:1/18:1/18:1) + NH4_MS2 | 3.32 | 0.51 | 0.005 |
| TG(16:0/18:1/18:1) + _MS2 | 3.23 | 0.57 | 0.007 |
| TG 48:2; [M + NH4]+ | 3.23 | 0.60 | 0.007 |
| TG(16:0/18:1/18:1) + NH4_MS2 | 3.15 | 0.66 | 0.008 |
| TG 58:2; [M + NH4]+ | 2.91 | 0.62 | 0.012 |
| TG(18:1/14:0/18:1) + NH4_MS2 | 2.87 | 0.48 | 0.013 |
| TG 57:2; [M + NH4]+ | 2.81 | 0.61 | 0.015 |
| TG(18:0/18:1/18:1) + NH4_MS2 | 2.24 | 0.65 | 0.043 |
| TG 58:1; [M + NH4]+ | 2.16 | 0.33 | 0.049 |
| PC(40:5) + H_MS2 | −2.62 | −0.24 | 0.021 |
| lysoPC 18:0; [M + H]+ | −2.57 | −0.22 | 0.023 |
| plasmenyl-PE 34:0; [M + H]+ | −2.56 | −0.21 | 0.024 |
| PC(36:1) + H_MS2 | −2.52 | −0.18 | 0.026 |
| PE(18:0/18:1) + H_MS2 | −2.51 | −0.20 | 0.026 |
| PC(36:1) + H_MS2a | −2.49 | −1.23 | 0.027 |
| CerG1(d38:1 + O) + _MS2 | −2.36 | −0.19 | 0.035 |
| PC(36:2) + H_MS2 | −2.35 | −0.14 | 0.035 |
| PC(34:0) + H_MS2 | −2.34 | −0.20 | 0.036 |
| PC(40:4) + H_MS2 | −2.27 | −0.28 | 0.041 |
| PE(18:0p/22:4) + H_MS2 | −2.26 | −0.19 | 0.042 |
| Cer(d20:1/18:0) + H_MS2 | −2.25 | −0.35 | 0.042 |
| PE(16:0p/22:6) + H_MS2 | −2.18 | −0.18 | 0.048 |
| plasmenyl-PC 40:5; [M + H]+ | −2.17 | −0.23 | 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraser, K.; Ryan, L.; Dilger, R.N.; Dunstan, K.; Armstrong, K.; Peters, J.; Stirrat, H.; Haggerty, N.; MacGibbon, A.K.H.; Dekker, J.; et al. Impacts of Formula Supplemented with Milk Fat Globule Membrane on the Neurolipidome of Brain Regions of Piglets. Metabolites 2022, 12, 689. https://doi.org/10.3390/metabo12080689
Fraser K, Ryan L, Dilger RN, Dunstan K, Armstrong K, Peters J, Stirrat H, Haggerty N, MacGibbon AKH, Dekker J, et al. Impacts of Formula Supplemented with Milk Fat Globule Membrane on the Neurolipidome of Brain Regions of Piglets. Metabolites. 2022; 12(8):689. https://doi.org/10.3390/metabo12080689
Chicago/Turabian StyleFraser, Karl, Leigh Ryan, Ryan N. Dilger, Kelly Dunstan, Kelly Armstrong, Jason Peters, Hedley Stirrat, Neill Haggerty, Alastair K. H. MacGibbon, James Dekker, and et al. 2022. "Impacts of Formula Supplemented with Milk Fat Globule Membrane on the Neurolipidome of Brain Regions of Piglets" Metabolites 12, no. 8: 689. https://doi.org/10.3390/metabo12080689
APA StyleFraser, K., Ryan, L., Dilger, R. N., Dunstan, K., Armstrong, K., Peters, J., Stirrat, H., Haggerty, N., MacGibbon, A. K. H., Dekker, J., Young, W., & Roy, N. C. (2022). Impacts of Formula Supplemented with Milk Fat Globule Membrane on the Neurolipidome of Brain Regions of Piglets. Metabolites, 12(8), 689. https://doi.org/10.3390/metabo12080689