

Anthraquinone Production from Cell and Organ Cultures of Rubia Species: An Overview

 , ,
, ,
Abstract
:
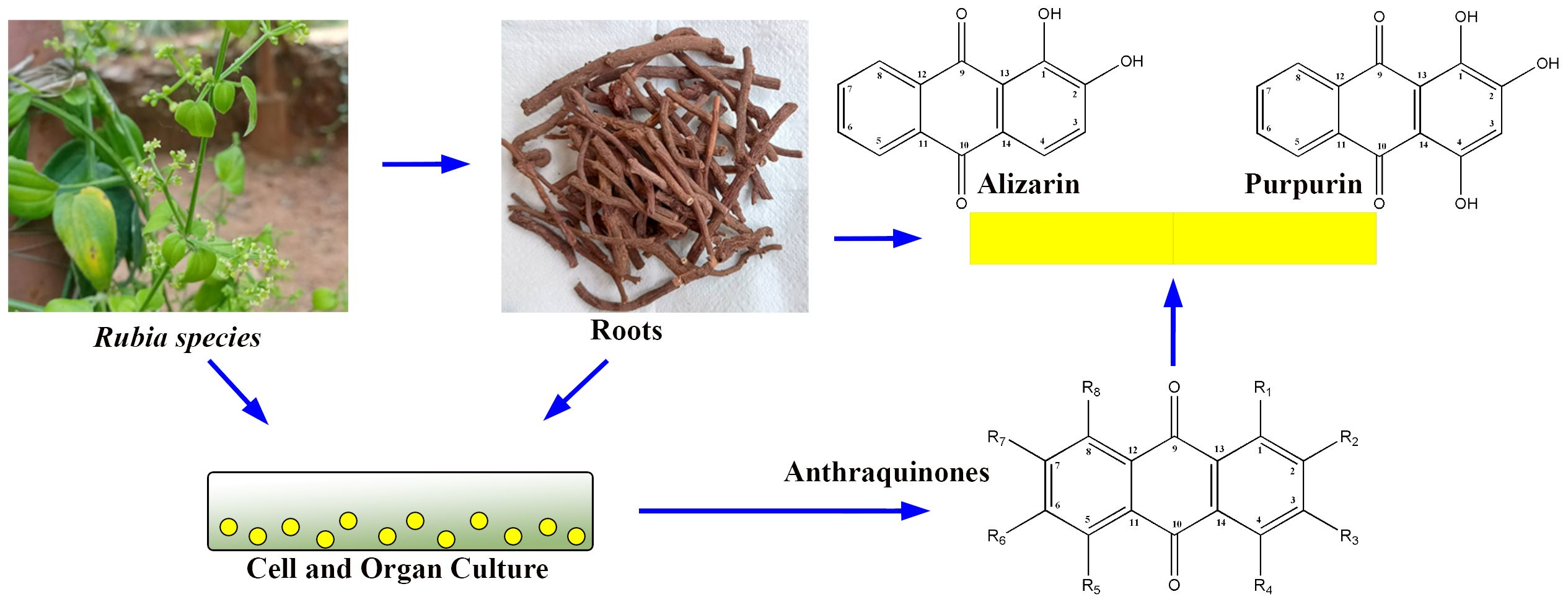
1. Introduction
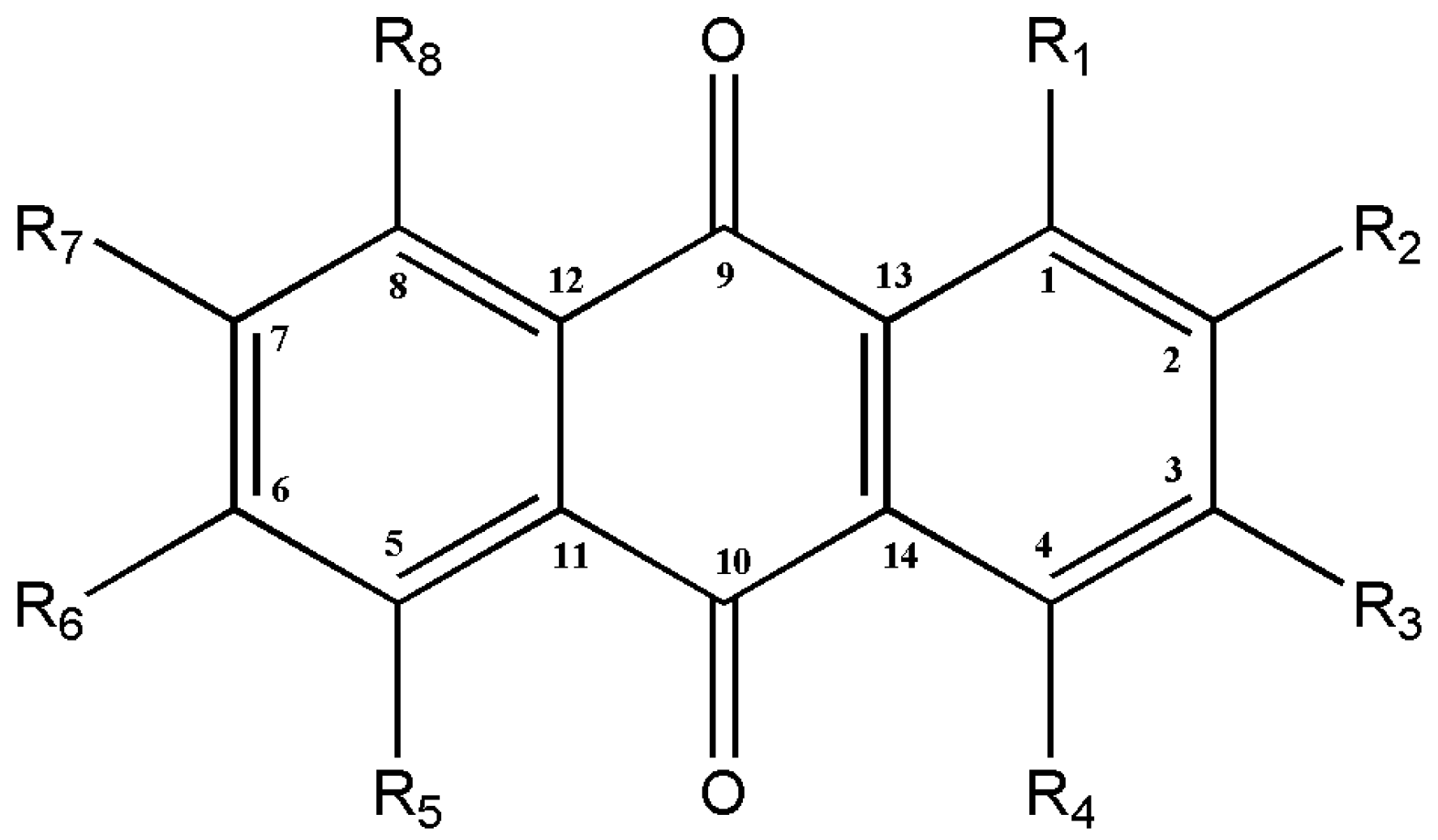
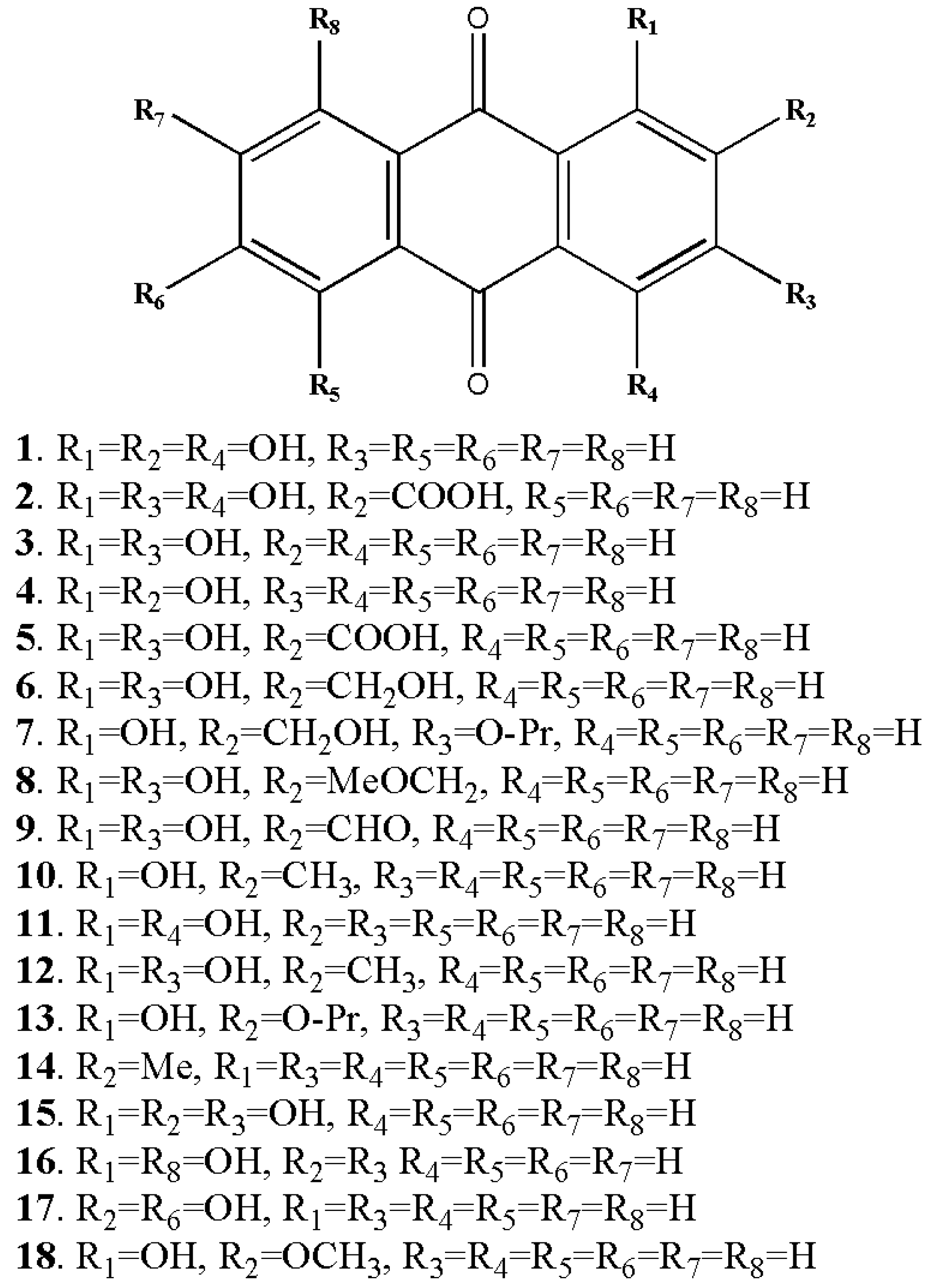
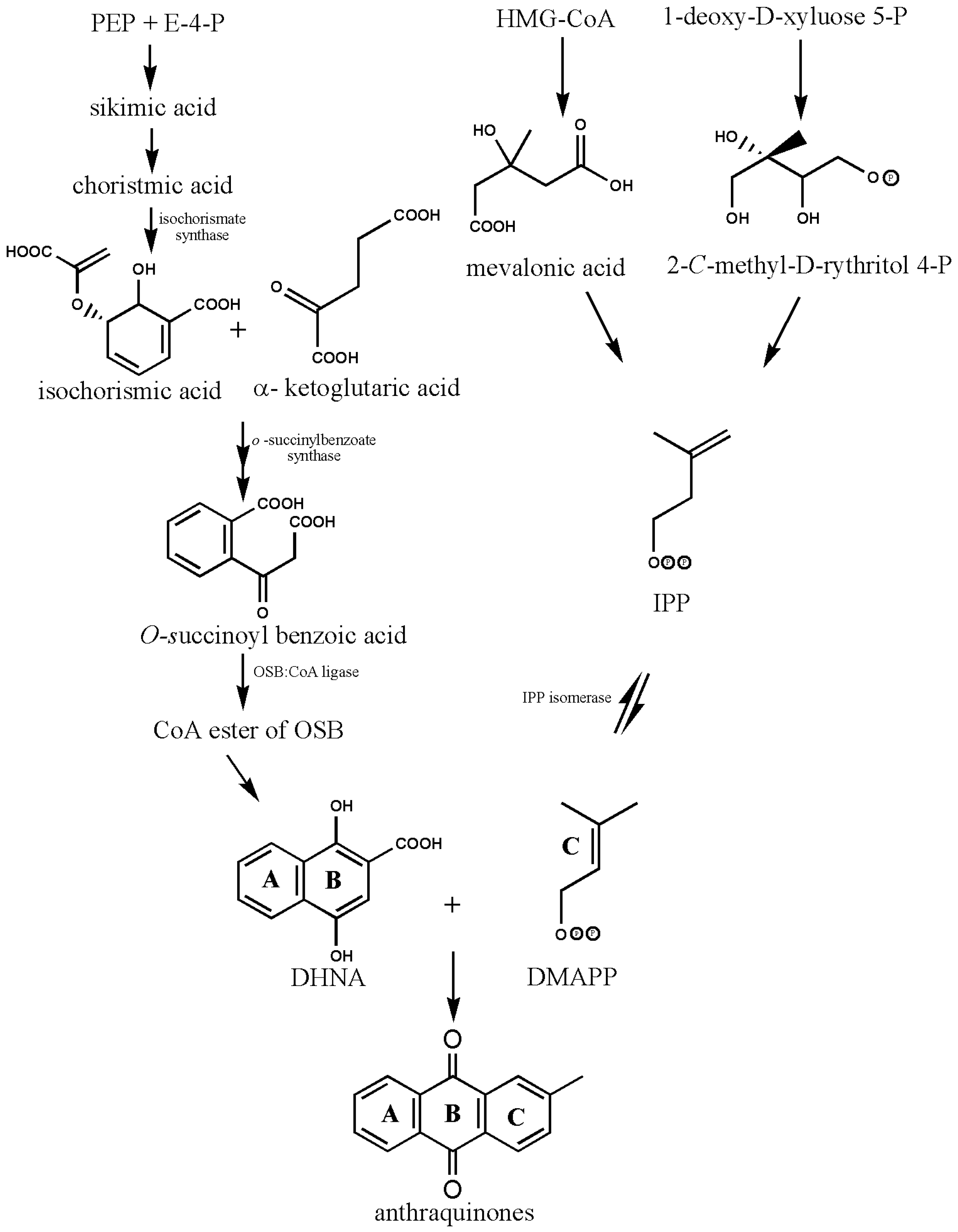
2. Biosynthesis of Anthraquinones
3. Callus Cultures for the Production of Anthraquinones
3.1. Optimization of Chemical and Physical Parameters
3.2. Transformation of Root Loci (rolA, rolB, rolC) and Other Genes and Elicitation
3.3. Immobilization and Other Strategies
4. Cell Suspension Cultures for the Production of Anthraquinones
5. Hairy and Adventitious Root Cultures for the Production of Anthraquinones
6. Bioreactor Cultures for the Production of Anthraquinones
7. Conclusions and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, R.; Geetanjali; Chauhan, S.M.S. 9,10-anthraquinones and other biologically active compounds from the genus Rubia. Chem. Biodivers. 2004, 1, 1241–1264. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.X.; Wang, P.; Wang, L.; Liu, C.; Xu, S.; Cheng, Y.; Wang, Y.; Li, Q.; Lei, H. Quinone derivatives from the genus Rubia and their bioactivities. Chem. Biodivers. 2014, 11, 341–363. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Chen, Q.; Chen, W.; Yang, J.; Zhou, X.; Zhang, C.; Wu, A.; Lai, J.; Chen, J.; Mei, Q.; et al. A comprehensive review of Rubia cordifolia L.: Traditional uses, phytochemistry, pharmacological activities, and clinical applications. Front. Pharmacol. 2022, 13, 965390. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, R.S. Natural dyes in madder (Rubia spp.) and their extraction and analysis in historical textiles. Color. Technol. 2017, 133, 449–462. [Google Scholar] [CrossRef]
- Caro, Y.; Anamale, L.; Fouillaud, M.; Laurent, P.; Petit, T.; Dufosse, L. Natural hydroxyanthraquinoid pigments as potent food grade colorants: An overview. Nat. Prod. Bioprospect. 2012, 2, 174–193. [Google Scholar] [CrossRef]
- Bechtold, T.; Mussak, R. Handbook of Natural Colorants; Wiley: Chichester, UK, 2009. [Google Scholar]
- Angelini, L.G.; Pistelli, L.; Belloni, P.; Bertoli, A.; Panconesi, S. Rubia tinctorum a source of natural dyes: Agronomic evaluation, quantitative analysis of alizarin and industrial assays. Ind. Crops Prod. 1997, 6, 303–311. [Google Scholar] [CrossRef]
- Banyai, P.; Kuzovkina, N.; Kursinszki, L.; Szoke, E. HPLC analysis of alizarin and purpurin produced by Rubia tinctorum L. hairy root cultures. Chromatographia 2006, 63, S111–S114. [Google Scholar] [CrossRef]
- Drivas, I.; Blackburn, R.S.; Rayner, C.M. Natural anthraquinoid colorants as platform chemicals in the synthesis of sustainable disperse for polyesters. Dyes Pigm. 2011, 88, 7–17. [Google Scholar] [CrossRef]
- Kaur, P.; Chandel, M.; Kumar, S.; Kumar, N.; Singh, B.; Kaur, S. Modulatory role of alizarin from Rubia cordifolia L. against genotoxicity of mutagens. Food Chem. Toxicol. 2010, 48, 320–325. [Google Scholar] [CrossRef]
- Rafaelly, L.; Heron, S.; Nowik, W.; Tchapla, A. Optimization of ESI-MS detection for HPLC of anthraquinone dyes. Dyes Pigm. 2008, 77, 19–203. [Google Scholar] [CrossRef]
- Singh, J.; Husain, Y.; Luqman, S.; Meena, A. Purpurin: A natural anthraquinone with multifaceted pharmacological activities. Phytother. Res. 2020, 35, 2418–2428. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, J.; Qi, W.; Su, R.; He, Z.; Peng, X. Alizarin and purpurin from Rubia tinctorum L. suppress insulin fibrillation and reduce the amyloid-induced cytotoxicity. ACS Chem. Neurosci. 2021, 12, 2182–2193. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, E.S.B.; Hulme, A.N.; McNab, H.; Quye, A. The natural constituents of historical textile dyes. Chem. Soc. Rev. 2004, 33, 329–336. [Google Scholar] [CrossRef]
- Baydar, H.; Karadogan, T. Agronomic potential and industrial value of madder (Rubia tinctorum L.) as a dye crop. Turk. J. Agric. For. 2006, 30, 287–293. [Google Scholar]
- Murthy, H.N.; Lee, E.J.; Paek, K.Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tiss. Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Verpoorte, R.; Memelink, J. Engineering secondary metabolite production in plants. Curr. Opin. Biotechnol. 2002, 13, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Kim, Y.S.; Jeong, C.S.; Kim, S.J.; Zhong, J.J.; Paek, K.Y. Production of Ginsenosides from Adventitious Root Cultures of Panax ginseng. In Production of Biomass and Bioactive Compounds Using Bioreactor Technology; Paek, K.Y., Murthy, H.N., Zhong, J.J., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 625–651. [Google Scholar]
- Murthy, H.N.; Dalawai, D. Biotechnological production of diterpenoid lactones from cell and organ cultures of Andrographis paniculata. Appl. Microbiol. Biotechnol. 2021, 105, 7683–7694. [Google Scholar] [CrossRef]
- Murthy, H.N. Biotechnological production of bacosides from cell and organ cultures of Bacopa monnieri. Appl. Microbiol. Biotechnol. 2022, 106, 1799–1811. [Google Scholar] [CrossRef]
- Han, Y.S.; Heijden, R.V.D.; Lefeber, A.W.M.; Erkelens, C.; Verpoorte, R. Biosynthesis of anthraquinones in cell cultures of Cinchona ‘Robusta’ proceeds via the methylerythritol-4-phosphate pathway. Phytochemistry 2002, 59, 45–55. [Google Scholar] [CrossRef]
- Han, Y.S.; Heijden, R.V.D.; Verpoorte, R. Biosynthesis of anthraquinones in cell cultures of the Rubiaceae. Plant Cell Tiss. Organ Cult. 2001, 67, 201–220. [Google Scholar] [CrossRef]
- Leistner, E. Biosynthesis of Chorismate-Derived Quinones in Plant Cell Cultures. In Primary and Secondary Metabolism of Plant Cell Cultures. Proceedings in Life Sciences; Neumann, K.H., Barz, W., Reinhard, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 215–224. [Google Scholar]
- Shin, S.H. Studies on the production of anthraquinone derivatives by tissue culture of Rubia species. Arch. Pharm. Res. 1989, 12, 99–102. [Google Scholar] [CrossRef]
- Mischenko, N.P.; Fedoreyev, S.A.; Glazunov, V.P.; Chernoded, G.K.; Bulgakov, V.P.; Zhuravlev, Y.N. Anthraquinone production by callus cultures of Rubia cordifolia. Fitoterapia 1999, 70, 552–557. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Khodakovskaya, M.V.; Glazunov, V.P.; Radchenko, S.V.; Zvereva, E.V.; Fedoreyev, S.A.; Zhuravlev, Y.N. Effect of salicylic acid, methyl jasmonate, ethephon, and cantharidin on anthraquinone production by Rubia cordifolia callus cultures transformed with the rolB and rolC genes. J. Biotechnol. 2002, 97, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Bulgakov, V.P.; Tehernoded, G.K.; Mischenko, N.P.; Shkryl, Y.N.; Glazunov, V.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. Effects of Ca2+ channel blockers and protein kinase/phosphatase inhibitors on growth and anthraquinone production in Rubia cordifolia callus cultures transformed by rolB and rolC genes. Planta 2003, 217, 349–355. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Shkryl, Y.N.; Glazunov, V.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. Increase in anthraquinone content in Rubia cordifolia cells transformed by rol genes does not involve activation of the NADPH oxidase signaling pathway. Biochemistry 2003, 68, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Shkryl, Y.N.; Fedoreyev, S.A.; Zhuravlev, Y.N. The rolB and rolC genes activate synthesis of anthraquinones in Rubia cordifolia cells by mechanism independent of octadecanoid signaling pathway. Plant Sci. 2004, 166, 1069–1075. [Google Scholar] [CrossRef]
- Mishchenko, N.P.; Fedoreev, S.A.; Bryukhanov, V.M.; Zverev, Y.F.; Lampatov, V.V.; Azarova, O.V.; Shkryl, Y.N.; Chernoded, G.K. Chemical composition and pharmacological activity of anthraquinones from Rubia cordifolia cell cultures. Pharm. Chem. J. 2007, 41, 605–609. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Aminin, D.I.; Shkryl, Y.N.; Gorpenchenko, T.Y.; Vermeichik, G.N.; Dmitrenok, P.S.; Zhuravlev, Y.N. Suppression of reactive oxygen species and enhanced stress tolerance in Rubia cordifolia cells expressing the rolC gene. Mol. Plant Microb. Interact. 2008, 21, 1561–1570. [Google Scholar] [CrossRef] [Green Version]
- Shkryl, Y.N.; Veremeichik, G.N.; Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. Individual and combined effects of the rolA, B, and C genes on anthraquinone production in Rubia cordifolia transformed calli. Biotechnol. Bioeng. 2008, 100, 118–125. [Google Scholar] [CrossRef]
- Shkryl, Y.N.; Veremeichik, G.N.; Bulgakov, V.P.; Gopenchenko, T.Y.; Aminin, D.L.; Zhuravlev, Y.N. Decreased ROS level and activation of antioxidant gene expression in Agrobacterium rhizhogenes pRiA4-transformed calli of Rubia cordifolia. Planta 2010, 232, 1023–1032. [Google Scholar] [CrossRef]
- Shkryl, Y.N.; Veremeichik, G.N.; Bulgakov, V.P.; Zhuravlev, Y.N. Induction of anthraquinone biosynthesis in Rubia cordifolia cells by heterologous expression of a calcium-dependent protein kinase gene. Biotechnol. Bioeng. 2011, 108, 1734–1738. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Gorpenchenko, T.Y.; Shkryl, Y.N.; Veremeichik, G.N.; Mischenko, N.P.; Aavarmenko, T.V.; Fedoreyev, S.A.; Zhuravlev, Y.N. CDPK-driven changes in the intracellular ROS level and plant secondary metabolism. Bioeng. Bugs. 2011, 2, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Veremeichik, G.N.; Shkryl, Y.N.; Bulgakov, V.P.; Avramenko, T.V.; Zhuravlev, Y.N. Molecular cloning and characterization of seven class III peroxidases induced by overexpression of agrobacterial rolB gene in Rubia cordifolia transgenic callus cultures. Plant Cell Rep. 2012, 31, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Bulgakov, V.P.; Gorpenchenko, T.Y.; Veremeichik, G.N.; Shkryl, Y.N.; Tcherenoded, G.K.; Bulgakov, D.; Aminin, D.L.; Zhuravlev, Y.N. The rolB gene suppresses reactive oxygen species in transformed plant cells through the sustained activation of antioxidant defense. Plant Physiol. 2012, 158, 1371–1381. [Google Scholar] [CrossRef] [Green Version]
- Nartop, P.; Akay, S.; Gurel, A. Immobilization of Rubia tinctorum L. suspension cultures and its effects on alizarin and purpurin accumulation and biomass production. Plant Cell Tiss. Organ Cult. 2013, 112, 123–128. [Google Scholar] [CrossRef]
- Shkryl, Y.N.; Veremeichik, G.N.; Makhazen, D.S.; Silantieva, S.A.; Mishchenko, N.P.; Vasileva, E.A.; Fedoreyev, S.A.; Bulgakov, V.P. Increase of anthraquinone content in Rubia cordifolia cells transformed by native and constitutively active forms of the AtCPK1 gene. Plant Cell Rep. 2016, 35, 1907–1916. [Google Scholar] [CrossRef]
- Veremeichik, G.N.; Bulgakov, V.P.; Shkryl, Y.N.; Silantieva, S.A.; Makhazen, D.S.; Tchnernoded, G.K.; Mischenko, N.P.; Fedoreyev, S.A.; Vasileva, E.A. Activation of anthraquinone biosynthesis in long-cultured callus culture of Rubia cordifolia transformed with the rolA plant oncogene. J. Biotechnol. 2019, 306, 38–46. [Google Scholar] [CrossRef]
- Mariadoss, A.; Satdive, R.; Fulzele, D.P.; Ramamoorthy, S.; Doss, G.P.C.; Zayed, H.; Younes, S.; Rajasekaran, C. Enhanced production of anthraquinones by gamma-irradiated cell cultures of Rubia cordifolia in a bioreactor. Ind. Crops Prod. 2020, 145, 111987. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Suzuki, H.; Matsumoto, T.; Mikami, Y. Effects of nutritional factors on the formation of anthraquinones by Rubia cordifolia plant cells in suspension culture. Agric. Biol. Chem. 1984, 48, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Matsumoto, T.; Mikami, Y. Effects of physical factors and surface-active agents on the formation of anthraquinones by Rubia cordifolia cells in suspension culture. Agric. Biol. Chem. 1985, 49, 519–520. [Google Scholar] [CrossRef]
- Sato, K.; Goda, Y.; Kawasaki, Y.; Okuyama, E.; Yoshihara, K.; Ozeki, Y.; Nakamura, M. Characteristic of anthraquinone production in plant roots and cell suspension cultures of Rubia tinctorum and R. akane. Plant Tiss. Cult. Lett. 1992, 9, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.H.; Shin, J.H.; Kim, J.H.; Chung, I.S.; Lee, H.J. Effect of chitosan elicitation and media components on the production of anthraquinone colorants in madder (Rubia akane Nakai) cell culture. Biotechnol. Bioprocess. Eng. 1999, 4, 300–304. [Google Scholar] [CrossRef]
- van Tegelen, L.J.P.; Bongaerts, R.J.M.; Croes, A.F.; Verpoorte, R.; Wullems, G.J. Isochorismate synthase isoforms from elicited cell cultures of Rubia tinctorum. Phytochemistry 1999, 51, 263–269. [Google Scholar] [CrossRef]
- Eichinger, D.; Bacher, A.; Zenk, M.H.; Eisenreich, W. Quantitative assessment of metabolic flux by 13C NMR analysis. Biosynthesis of anthraquinones in Rubia tinctorum. J. Am. Chem. Soc. 1999, 121, 7469–7475. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Giuletti, A.M.; Picotto, G.; Rodriguez-Talou, J.; Boland, R. Involvement of the PLC/PKC pathway in chitosan-induced anthraquinone production by Rubia tinctorum L. cell cultures. Plant Sci. 2003, 165, 429–436. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Giulietti, A.M.; Boland, R. Signal transduction events mediating chitosan stimulation of anthraquinone synthesis in Rubia tinctorum. Plant Sci. 2004, 166, 405–413. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Morelli, S.; Picotto, G.; Giulietti, A.M.; Boland, R. Intracellular calcium mobilization: A key step for chitosan-induced anthraquinone production in Rubia tinctorum. Plant Sci. 2005, 169, 712–720. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Picotto, G.; Giuletti, A.M.; Boland, R. Involvement of G-proteins in chitosan-induced anthraquinone synthesis in Rubia tinctorum. Physiol. Plant. 2006, 128, 29–37. [Google Scholar] [CrossRef]
- Orban, N.; Boldizsar, I.; Szucs, Z.; Danos, B. Influence of different elicitors on the synthesis of anthraquinone derivatives in Rubia tinctorum L. cell suspension cultures. Dyes Pigm. 2008, 77, 249–257. [Google Scholar] [CrossRef]
- Perassolo, M.; Quevedo, C.; Busto, V.; Ianone, F.; Giulietti, A.M.; Talou, J.R. Enhance of anthraquinone production by effect of proline and aminoindan-2-phosphonic acid in Rubia tinctorum suspension cultures. Enzyme Mirob. Technol. 2007, 41, 181–185. [Google Scholar] [CrossRef]
- Perassolo, M.; Quevedo, C.V.; Giulietti, A.M.; Talou, J.R. Stimulation of the proline cycle and anthraquinone accumulation in Rubia tinctorum cell suspension cultures in the presence of glutamate and two proline analogs. Plant Cell Tiss. Organ Cult. 2011, 106, 153–159. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Linsmaier, E.M.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K. Hairy root culture for mass-production of high-value secondary metabolites. Crit. Rev. Biotechnol. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Adventitious roots and secondary metabolism. Chin. J. Biotechnol. 2008, 24, 711–716. [Google Scholar] [CrossRef]
- Murthy, H.N.; Dandin, V.S.; Paek, K.Y. Tools for biotechnological production of useful phytochemicals from adventitious root cultures. Phytochem. Rev. 2016, 15, 129–145. [Google Scholar] [CrossRef]
- Shin, S.W.; Kim, Y.S. Production of anthraquinone derivatives by hairy roots of Rubia cordifolia var. pratensis. Kor. J. Pharmacogn. 1996, 27, 301–308. [Google Scholar]
- Lodhi, A.H.; Charlwood, B.V. Agrobacterium rhizogenes-mediated transformation of Rubia peregrina L.: In vitro accumulation of anthraquinones. Plant Cell Tiss Organ Cult. 1996, 46, 103–108. [Google Scholar] [CrossRef]
- Lodhi, A.H.; Bongaerts, R.J.M.; Verpoorte, R.; Coomber, S.A.; Charlwood, B.V. Expression of bacterial isochorismate synthase (EC 5.4.99.6) in transgenic root cultures of Rubia peregrina. Plant Cell Rep. 1996, 16, 54–57. [Google Scholar] [CrossRef]
- Park, S.U.; Kim, Y.K.; Lee, S.Y. Establishment of hairy root culture of Rubia akane Nakai for alizarin and purpurin production. Sci. Res. Essays 2009, 4, 94–97. [Google Scholar] [CrossRef]
- Park, S.U.; Lee, S.Y. Anthraquinone production by hairy root culture of Rubia akane Nakai: Influence of media and auxin treatment. Sci. Res. Essays 2009, 4, 690–693. [Google Scholar] [CrossRef]
- Sato, K.; Yamazaki, T.; Okuyama, E.; Yoshihira, K.; Shimomura, K. Anthraquinone production by transformed root cultures of Rubia tinctorum: Influence of phytohormones and sucrose concentration. Phytochemistry 1991, 30, 1507–1509. [Google Scholar] [CrossRef]
- Kino-Oka, M.; Mine, K.; Taya, M.; Tone, S.; Ichi, T. Production and release of anthraquinone pigments by hairy roots of madder (Rubia tinctorum L.) under improved culture conditions. J. Ferment. Bioeng. 1994, 77, 103–106. [Google Scholar] [CrossRef]
- Perassolo, M.; Cardillo, A.B.; Mugas, M.L.; Montoya, S.C.N.; Giulietti, A.M.; Talou, J.R. Enhancement of anthraquinone production and release by combination of culture medium selection and methyl jasmonate elicitation in hairy root cultures of Rubia tinctorum. Ind. Crops Prod. 2017, 105, 124–132. [Google Scholar] [CrossRef]
- Bicer, P.O.; Demirci, T.; Asci, O.A.; Baydar, N.G. Effects of methyl jasmonate and caffeic acid applications on secondary metabolites production in madder (Rubia tinctorum) root cultures. Indian J. Pham. Educ. Res. 2017, 51, S508–S512. [Google Scholar] [CrossRef] [Green Version]
- Demicri, T.; Asci, O.A.; Baydar, N.G. Influence of salicylic acid and L-phenylalanine on the accumulation of anthraquinone and phenolic compounds in adventitious root cultures of madder (Rubia tinctorum L.). Plant Cell Tiss. Organ Cult. 2021, 144, 313–324. [Google Scholar] [CrossRef]
- Nitisch, J.P.; Nitsch, C. Haploid plants form pollen grains. Science 1969, 163, 65–87. [Google Scholar] [CrossRef]
- Murthy, H.N.; Dalawai, D.; Bhat, M.A.; Dandin, V.S.; Paek, K.Y.; Park, S.Y. Biotechnological Production of Useful Phytochemicals from Adventitious Root Cultures. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawath, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer Nature: Geneva, Switzerland, 2021; pp. 469–486. [Google Scholar] [CrossRef]
- Murthy, H.N.; Georgiev, M.I.; Kim, Y.S.; Jeong, C.S.; Kim, S.J.; Park, S.Y.; Paek, K.Y. Ginsenosides: Prospective for sustainable biotechnological production. Appl. Microbiol. Biotechnol. 2014, 98, 6243–6254. [Google Scholar] [CrossRef]
- Murthy, H.N.; Kim, Y.S.; Georgiev, M.I.; Paek, K.Y. Biotechnological production of eleutherosides: Current state and perspectives. Appl. Microbiol. Biotechnol. 2014, 98, 7319–7329. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Kim, Y.S.; Park, S.Y.; Paek, K.Y. Biotechnological production of caffeic acid derivatives from cell and organ cultures of Echinacea species. Appl. Microbiol. Biotechnol. 2014, 98, 7707–7717. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Kim, Y.S.; Park, S.Y.; Paek, K.Y. Hypericins: Biotechnological production from cell and organ cultures. Appl. Microbiol. Biotechnol. 2014, 98, 9187–9198. [Google Scholar] [CrossRef]
- Laszlo, M.; Kretovics, J.; Danos, B.; Szokan, G.; Liszt, K.; Hollosy, F.; Toth, Z.; Gyurjan, I. The production of secondary metabolites by plant cells of Rubia tinctorum cultivated in bioreactors. Plant Med. Suppl. 1992, 58, 613. [Google Scholar] [CrossRef]
- Busto, V.D.; Rodriguez-Talou, J.; Giulietti, A.M.; Merchuk, J.C. Effect of shear stress on anthraquinones production in Rubia tinctorum suspension cultures. Biotechnol. Prog. 2008, 24, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Busto, V.D.; Calabro-Lopez, A.; Rodriguez-Talou, J.; Giulietti, A.M.; Merchuk, J.C. Anthraquinones production in Rubia tinctorum cell suspension cultures: Down scale of shear effects. Biochem. Eng. J. 2013, 77, 119–128. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Major Anthraquinone Compounds | References |
|---|---|---|---|
| R. peregrina L. | Wild madder | Pseudopurpurin | [6] |
| Rubia tinctorum L. | Dyer’s madder or European madder | Alizarin, alizarin-2-methyl ether, anthraflavin, anthragallol, danthron, lucidin, lucidin perimerveroside, lucidin, lucidin-ω-ethyl-ether, munjistin, munjistin ethyl ether, nordamnacanthal, purpurin, pseudopurpurin, quinizarin, ruberythric acid, xanthopurpurin | [7,8,9] |
| R. cordifolia L. | Indian madder | Alizarin, lucidin, munjinstin, pseudopurpurin, purpurin, rubiadin, tectoquinone, xnathopurpurin | [6,9,10] |
| R. akane Makino | Japanese madder | Purpurin, ruberythric acid | [6,11] |
| Species | Medium, Growth Regulators and Parameters Studied | Metabolites and Yield | References |
|---|---|---|---|
| Rubia cordifolia and R. akane | MS medium with kinetin (5 ppm) + 2,4-D or NAA (2 ppm) | AQs—Not reported | [24] |
| Rubia cordifolia | MS medium 0.5 mg L−1 BA and 2 mg L−1 NAA | AQs—0.62–1.22% DW | [25] |
| Rubia cordifolia | Transformed with 35S-rolB and 35S-rolC genes and compared transgenic callus with non-transformed callus cultures; additionally, callus cultures were treated with (1, 10, 100 μM) MJ or SA or ethephone | AQs—4.33% DW | [26] |
| Rubia cordifolia | Studied the effect of Ca2+ channel blockers (verapamil and L-type Ca2+) and inhibitors of protein phosphatases (protein phosphatase 1 and 2A) on growth and anthraquinone production in non-transformed and rolB and rolC transformed callus | AQs—3.02% DW | [27] |
| Rubia cordifolia | Studied the effect of rol genes on AQ content and showed that induction of AQ production in transgenic cultures does not involve the activation of Ca2+-dependent NADPH oxidase pathway | AQs—3.17% DW | [28] |
| Rubia cordifolia | Studied the effect of octadecanoid pathway inhibitors (diethyldithiocarbamate, propyl gallate, salicylhydroxamic acid, and piroxicam) on the synthesis of AQs | AQs—5.00% DW | [29] |
| Rubia cordifolia | Analyzed the chemical composition and pharmacological activity of AQs obtained from callus cultures | AQs—2.14% | [30] |
| Rubia cordifolia | Studied the effect of the rolC gene on the production of reactive oxygen species and AQ | AQs—Not reported | [31] |
| Rubia cordifolia | Studied the effect of rolA, rolB, and rolC genes individually and combined the effect of the above genes on the accumulation of AQs | AQs—5.85% DW | [32] |
| Rubia cordifolia | Studied the expression of the key antioxidant gene in R. cordifolia transformed with Agrobacterium rhizogenes pRiA4-transformed calli | AQs—Not reported | [33] |
| Rubia cordifolia | Transformed calcium-dependent protein kinase gene of Arabidopsis (AtCPK1) into R. cordifolia | AQs—4.36% DW | [34] |
| Rubia cordifolia | Studied the effect of AtCPK1 transgenic R. cordifolia cells on the production of reactive oxygen species | AQs—Not reported | [35] |
| Rubia cordifolia | Studied the expression of peroxidase genes that were isolated from R. cordifolia, namely RcPrx01-RcPrx07, in R. cordifolia in aerial organs as well as in cells transformed with the rolB and rolC genes | AQs—Not reported | [36] |
| Rubia cordifolia | Studied the effect of rolB gene in the suppression of reactive oxygen species in R. cordifolia rolB-expressing cells. | AQs—Not reported | [37] |
| Rubia cordifolia | Immobilization of suspension cultures with loofa sponge, sisal, and jute mixture (2 g each) and re-cultured in MS liquid medium | AQs—1.32 and 0.55 mg g−1 DW | [38] |
| Rubia cordifolia | Transformation with the AtCPK1 gene (calcium-dependent protein kinase) and obtained increased AQs accumulation | AQs—4.64% DW | [39] |
| Rubia cordifolia | Studied the effect of rolA gene expression in long-term cultured callus cultures and obtained increased AQs accumulation | AQs—45 mg g−1 DW | [40] |
| Rubia cordifolia | Studied the effect of 2, 4, 6, 8, 10, 12, 14, and 16 Gy of gamma irradiation and callus was cultured on MS + 1 mg L−1 IAA + 1 mg L−1 NAA + 1 mg L−1 BA and the callus cultures irradiated at 8 Gy showed optimum accumulation of AQs | AQs—26.86 and 44.85 mg g−1 DW | [41] |
| Species | Culture Medium, Growth Regulators, Additives, and Other Parameters | Metabolites and Yield or Productivity | References |
|---|---|---|---|
| Rubia cordifolia | MS medium and studied the effect of auxins IAA or NAA or 2,4-D; concentrations of NAA (0.2–5.0 mg L−1); ratio of NH4 nitrogen and NO3 nitrogen (only NO3, 1:1, 2:1, 3:1, only NH4); sucrose, fructose, glucose, galactose, raffinose, lactose, rhamonose; sucrose concentration (2–7%), myo-inositol concentration (10–250 mg L−1) on the synthesis of anthraquinones | AQs—50 μmol g−1 day−1 | [43] |
| Rubia cordifolia | MS + 2.0 mg L−1 NAA + 0.2 mg L−1 KN +sucrose (5%) +10 mg L−1 myo-inositol and studied effect of light on AQ production | AQs—50 μmol g−1 day−1 | [44] |
| Rubia tinctorum and R. akane | Callus induction: MS + 5 μM NAA + 0.1 μM KN for R. tinctorum; MS + 0.5 μM 2,4-D for R. akane. Cell suspension cultures were established by using LS +0.5 μM NAA + 0.1 μM KN | AQs—Not reported | [45] |
| Rubia akane | SH + 2 mg L−1 2,4-D + 3% sucrose; Various elicitors chitosan, alginate, kappa carrageenan, yeast extract, gum Arabic, lichenan, xylan, nigeran were tested at 0, 20, 40, and 60 mg L−1. | AQs—320 mg g−1 DW | [46] |
| Rubia tinctorum | B5 medium + 2% sucrose + 2 mg L−1 2,4-D + 0.5 mg L−1 NAA, 0.5 mg l−1 IAA, and 0.5 mg L−1 KN and cells were elicited with an autoclave extract of Pythium aphanidermatum | AQs—Not reported | [47] |
| Rubia tinctorum | B5 medium + 10 μM 2,4-D + 1.5% (1-13C)- or (U-13C6) glucose and studied the 13C labeling pattern of CoA, pyruvate, phosphoenol pyruvate, and others to study the biosynthesis of anthraquinones | AQs—Not reported | [48] |
| Rubia tinctorum | B5 medium + 2% sucrose + 2 mg L−1 2,4-D + 0.5 mg L−1 NAA, 0.5 mg L−1 IAA and 0.5 mg L−1 KN and cells were elicited 200 mg L−1 chitosan. They studied signal transduction pathways | AQs—Not reported | [49,50,51,52] |
| Rubia tinctorum | MS + 3% sucrose +1 mg L−1 IAA + 0.2 mg L−1 NAA +0.2 mg L−1 KN; Elicitors 3, 7, 15, and 30 μL mL−1 JA or 13, 27, 67.5 and 100 μL mL−1 SA or 80 mg mL−1 polysaccharides isolated from fungus Coriolus versicolor | AQs—70 to 262 mg g−1 DW | [53] |
| Rubia tinctorum | B5 medium + 2% sucrose + 2 mg L−1 2,4-D + 0.5 mg L−1 NAA, 0.5 mg L−1 IAA and 0.5 mg L−1 KN; Effect of addition of 0.25 mM proline or 100 μM aminoinda-2-phosphonic acid on anthraquinone production | AQs—1.5 μmol g−1 FW | [54] |
| Rubia tinctorum | B5 medium + 2% sucrose + 2 mg L−1 2,4-D + 0.5 mg L−1 NAA, 0.5 mg L−1 IAA, and 0.5 mg L−1 KN; Studied the effect of the addition of glutamate and proline analogs (azatidine-2-carboxylic acid and thiazolidine-4-carboxylic acid) on the pentose phosphate pathway, the proline cycle and anthraquinone production | AQs—Not reported | [55] |
| Species and Type of Root Cultures | Culture Medium | Growth Regulators, Additives, and Other Parameters | Metabolites and Yield or Productivity | References |
|---|---|---|---|---|
| Rubia cordifolia var. pratensis; hairy roots | NN | They tested the effect of auxins viz. 0.5, 1.0 mg L−1 NAA, and 0.1 mg L−1 IAA on the growth and accumulation of metabolites. | AQs—Not reported | [62] |
| Rubia peregrina; hairy roots | B5 | Gamborg B5 medium containing 30 g L−1 sucrose, 10 g L−1 agar | AQs—2.12 mg g−1 DW | [63,64] |
| Rubia akane; hairy roots | MS | MS + 3% sucrose studied the accumulation of alizarin and purpurin | AQs—3.9 and 4.5 mg g−1 DW alizarin and purpurin | [65] |
| Rubia akane; hairy roots | Various media | Studied the effect of half and full strength of B5, half and full strength of MS, and a half and full strength of SH media; 0.1, 0.5, 1.0 mg L−1 IAA, IBA and NAA | AQs—5.9 and 7.2 mg g−1 DW alizarin and purpurin | [66] |
| R. tinctorum; hairy roots | MS | Studied the effect of 0.5, 5 μM IAA, NAA or 2,4-D, and 0.5 μM KN; and 3,6, 9, 12, 15, and 18% sucrose on the growth of roots and AQs production | AQs—700 μg g−1 FW | [67] |
| Rubia tinctorum; hairy roots | MS | Studied the effect of fructose galactose, glucose, lactose, maltose, and sucrose (2%); altered nitrogen (1.90 g L−1 KNO3 + 1.65 g L−1 NH4NO3) and (6.07 g L−1 KNO3) on AQs accumulation | AQs—0.72 mg g−1 day−1 | [68] |
| Rubia tinctorum; hairy roots; | 1/2 B5 | 2% sucrose + 2 mg L−1 2,4-D + 0.5 mg L−1 NAA, 0.5 mg L−1 IAA and 0.5 mg L−1 KN and cells were elicited by 100 μM MJ | AQs—33.4 μmol L−1 day−1 | [69] |
| Rubia tinctorum; adventitious roots | MS | Studied the effect of Methyl jasmonate (10 and 100 μM) and caffeic acid (1 and 2 mM) on the growth of roots and AQs | AQs—Not reported | [70] |
| Rubia tinctorum; adventitious roots | MS | MS +2% sucrose and studied the effect of L-phenylalanine (50 and 100 μM) and salicylic acid (20 and 40 μM) on the accumulation of AQs | AQs—31.47 mg g−1 DW | [71] |
| The Type of Bioreactor Used | Species and Type of Culture | Culture Medium and Growth Regulators/Additives | Optimization of Factors | Metabolites and Yield or Productivity | References |
|---|---|---|---|---|---|
| New Brunswick fermenters; 1.5 L | Rubia cordifolia, cell suspension culture | MS medium + 1 mg L−1 IAA + 1 mg L−1 NAA + 1 mg L−1 BA and 3% sucrose | The effect of helical and Rushton turbine impellers on the accumulation of anthraquinones | AQs—37.96 and 78.93 mg g−1 DW of alizarin and purpurin | [41] |
| Stirred tank bioreactors | Rubia tinctorum, cell suspension culture | - | - | AQs—Not reported | [78] |
| New Brunswick fermenters; 1.5 and 5 L | Rubia tinctorum, cell suspension culture | B5 medium + 2 mg L−1 NAA + 0.1 mg L−1 IAA + 0.2 mg L−1 KN + 2% sucrose | The effect of hydrodynamic stress on cell viability, biomass, and anthraquinone production | AQs—70.7 μmol g−1 day−1 | [79] |
| Stirred tank bioreactors | Rubia tinctorum, cell suspension culture | B5 medium + 2 mg L−1 NAA + 0.1 mg L−1 IAA + 0.2 mg L−1 KN + 2% sucrose | The effect of turbulence and light irradiation on cell viability, biomass, and anthraquinone production | AQs—681.3 μmol L−1 | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Anthraquinone Production from Cell and Organ Cultures of Rubia Species: An Overview. Metabolites 2023, 13, 39. https://doi.org/10.3390/metabo13010039
Murthy HN, Joseph KS, Paek KY, Park SY. Anthraquinone Production from Cell and Organ Cultures of Rubia Species: An Overview. Metabolites. 2023; 13(1):39. https://doi.org/10.3390/metabo13010039
Chicago/Turabian StyleMurthy, Hosakatte Niranjana, Kadanthottu Sebastian Joseph, Kee Yoeup Paek, and So Young Park. 2023. "Anthraquinone Production from Cell and Organ Cultures of Rubia Species: An Overview" Metabolites 13, no. 1: 39. https://doi.org/10.3390/metabo13010039
APA StyleMurthy, H. N., Joseph, K. S., Paek, K. Y., & Park, S. Y. (2023). Anthraquinone Production from Cell and Organ Cultures of Rubia Species: An Overview. Metabolites, 13(1), 39. https://doi.org/10.3390/metabo13010039