
Concentrations of Plasma Amino Acids and Neurotransmitters in Participants with Functional Gut Disorders and Healthy Controls

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Diet Record and Sample Collection
2.3. Proteogenic and Non-Proteogenic Amino Acid Analysis
2.4. Neurotransmitter Analysis
2.5. Statistical Analysis
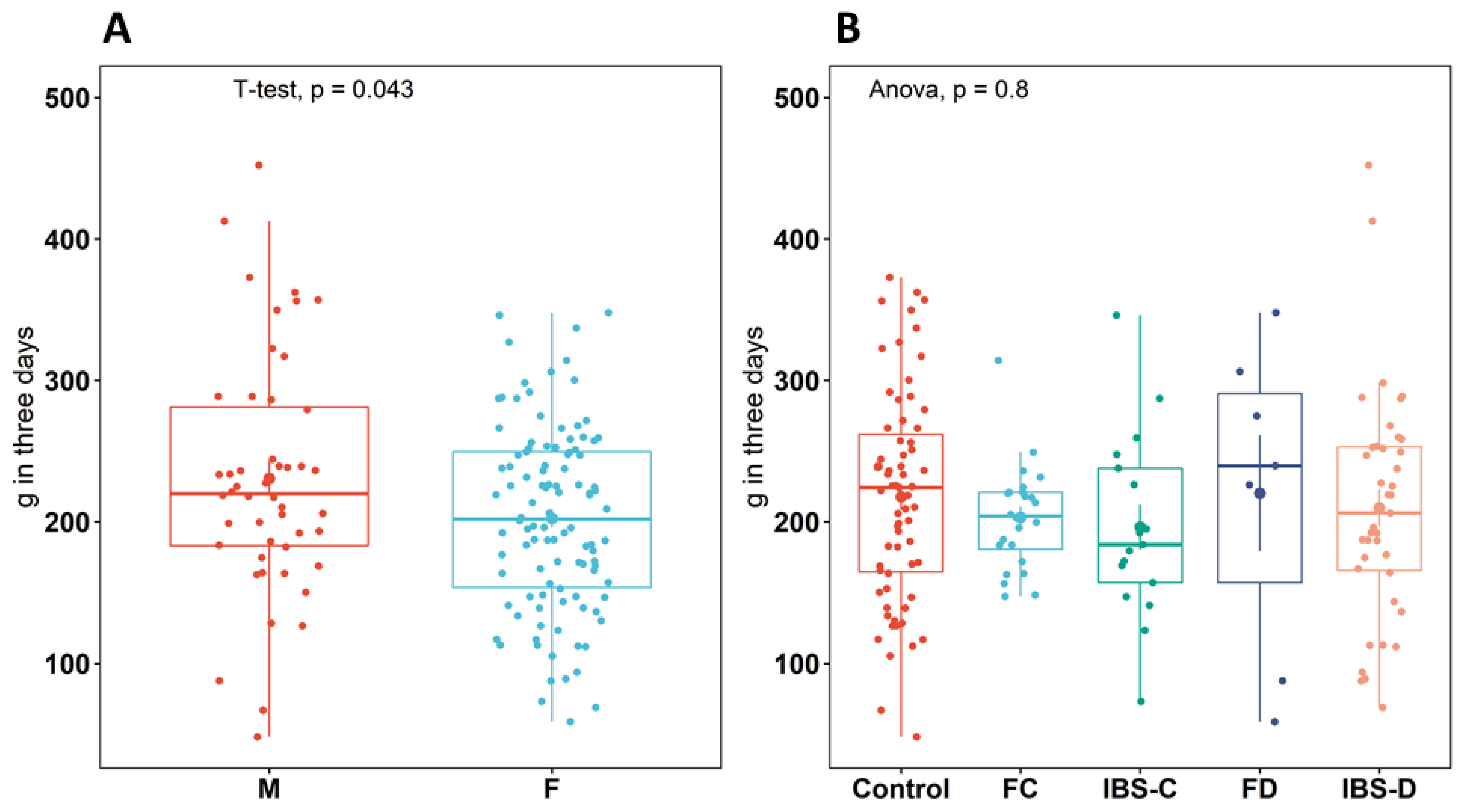
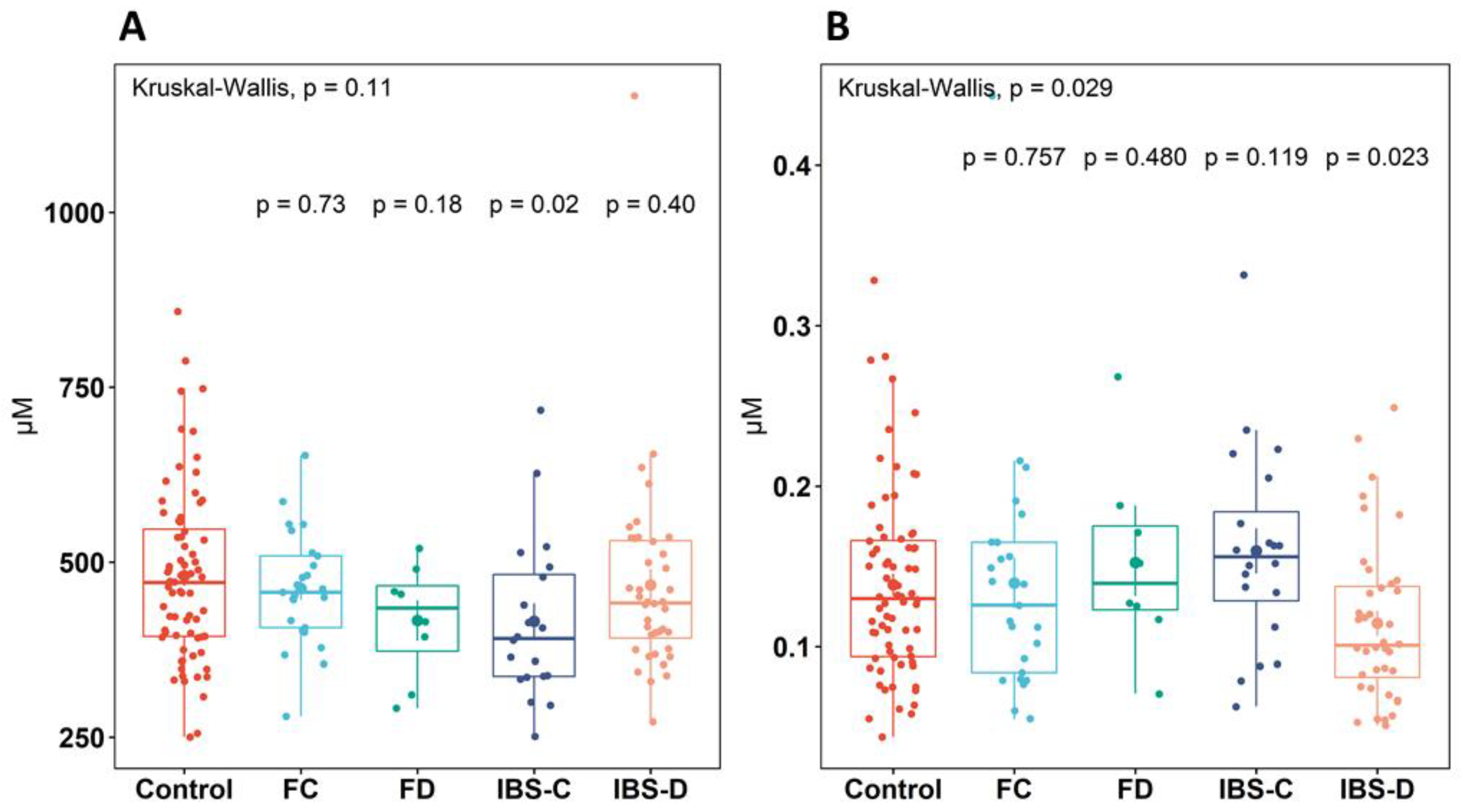
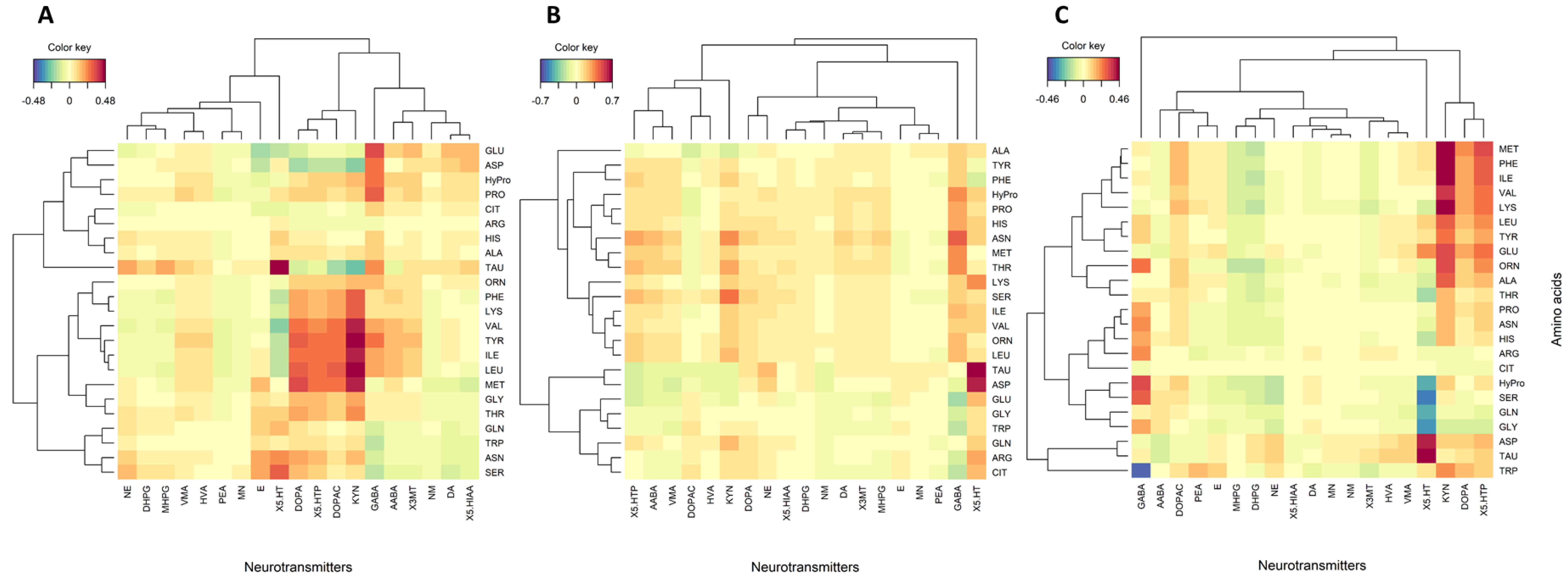
3. Results
3.1. Plasma Amino Acid and Neurotransmitter Concentrations
3.2. Correlations between Amino Acids and Neurotransmitters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Preter, V.; Verbeke, K. Metabolomics as a Diagnostic Tool in Gastroenterology. World J. Gastrointest. Pharm. 2013, 4, 97–107. [Google Scholar] [CrossRef]
- Solon-Biet, S.M.; Cogger, V.C.; Pulpitel, T.; Wahl, D.; Clark, X.; Bagley, E.E.; Gregoriou, G.C.; Senior, A.M.; Wang, Q.-P.; Brandon, A.E.; et al. Branched-chain amino acids impact health and lifespan indirectly via amino acid balance and appetite control. Nat. Metab. 2019, 1, 532–545. [Google Scholar] [CrossRef]
- Alqudah, A.; Wedyan, M.; Qnais, E.; Jawarneh, H.; McClements, L. Plasma Amino Acids Metabolomics’ Important in Glucose Management in Type 2 Diabetes. Front. Pharmacol. 2021, 12, 695418. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kido, J.; Matsumoto, S.; Shimizu, K.; Nakamura, K. Associations among amino acid, lipid, and glucose metabolic profiles in childhood obesity. BMC Pediatr. 2019, 19, 273. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Hu, C.-A.A. Therapeutic Potential of Amino Acids in Inflammatory Bowel Disease. Nutrients 2017, 9, 920. [Google Scholar] [CrossRef] [Green Version]
- Clarke, G.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. Irritable bowel syndrome: Towards biomarker identification. Trends Mol. Med. 2009, 15, 478–489. [Google Scholar] [CrossRef]
- Keshteli, A.H.; Madsen, K.L.; Mandal, R.; Boeckxstaens, G.E.; Bercik, P.; De Palma, G.; Reed, D.E.; Wishart, D.; Vanner, S.; Dieleman, L.A. Comparison of the metabolomic profiles of irritable bowel syndrome patients with ulcerative colitis patients and healthy controls: New insights into pathophysiology and potential biomarkers. Aliment. Pharmacol. Ther. 2019, 49, 723–732. [Google Scholar] [CrossRef]
- Dalangin, R.; Kim, A.; Campbell, R. The Role of Amino Acids in Neurotransmission and Fluorescent Tools for Their Detection. Int. J. Mol. Sci. 2020, 21, 6197. [Google Scholar] [CrossRef]
- Berstad, A.; Raa, J.; Valeur, J. Tryptophan: ‘Essential’ for the pathogenesis of irritable bowel syndrome? Scand J. Gastroenterol. 2014, 49, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Raskov, H.; Burcharth, J.; Pommergaard, H.C.; Rosenberg, J. Irritable bowel syndrome, the microbiota and the gut-brain axis. Gut Microbes 2016, 7, 365–383. [Google Scholar] [CrossRef] [Green Version]
- Gros, M.; Gros, B.; Mesonero, J.E.; Latorre, E. Neurotransmitter Dysfunction in Irritable Bowel Syndrome: Emerging Approaches for Management. J. Clin. Med. 2021, 10, 3429. [Google Scholar] [CrossRef]
- Rhoads, J.M.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino Acids 2009, 37, 111–122. [Google Scholar] [CrossRef]
- Branco, A.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of Histamine in Modulating the Immune Response and Inflammation. Mediat. Inflamm 2018, 2018, 9524075. [Google Scholar] [CrossRef] [Green Version]
- Markus, C.R. Dietary amino acids and brain serotonin function; implications for stress-related affective changes. Neuromol. Med. 2008, 10, 247–258. [Google Scholar] [CrossRef]
- Davenport, M.; Poles, J.; Leung, J.M.; Wolff, M.J.; Abidi, W.M.; Ullman, T.; Mayer, L.; Cho, I.; Loke, P. Metabolic alterations to the mucosal microbiota in inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 723–731. [Google Scholar] [CrossRef] [Green Version]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Faure, C.; Patey, N.; Gauthier, C.; Brooks, E.M.; Mawe, G.M. Serotonin Signaling Is Altered in Irritable Bowel Syndrome With Diarrhea but Not in Functional Dyspepsia in Pediatric Age Patients. Gastroenterology 2010, 139, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Portune, K.J.; Benítez-Páez, A.; Del Pulgar, E.M.G.; Cerrudo, V.; Sanz, Y. Gut microbiota, diet, and obesity-related disorders-The good, the bad, and the future challenges. Mol. Nutr. Food Res. 2017, 61, 1600252. [Google Scholar] [CrossRef] [Green Version]
- Heenan, P.; Creemers, R.H.; Sharma, S.; Keenan, J.; Bayer, S.; Young, W.; Cooney, J.; Armstrong, K.; Fraser, K.; Skidmore, P.M.; et al. Cohort Profile: The Christchurch IBS cOhort to investigate Mechanisms FOr gut Relief and improved Transit (COMFORT). Inflamm. Intest. Dis. 2020, 5, 132–143. [Google Scholar] [CrossRef]
- Simon, M.H.; Heenan, P.E.; Frampton, C.; Bayer, S.; Keenan, J.I.; Boer de, N.K.; Roy, N.C.; Talley, N.J.; Gearry, R.B. Economic living standard and abdominal pain mediate the association between functional gastrointestinal disorders and depression or anxiety. Neurogastroenterol. Motil. 2022, 35, e14465. [Google Scholar]
- James, S.C.; Fraser, K.; Young, W.; Heenan, P.E.; Gearry, R.B.; Keenan, J.I.; Talley, N.J.; Joyce, S.A.; McNabb, W.C.; Roy, N.C. Concentrations of Fecal Bile Acids in Participants with Functional Gut Disorders and Healthy Controls. Metabolites 2021, 11, 612. [Google Scholar] [CrossRef]
- Milan, A.M.; D’Souza, R.F.; Pundir, S.; A Pileggi, C.; Barnett, M.; Markworth, J.F.; Cameron-Smith, D.; Mitchell, C. Older adults have delayed amino acid absorption after a high protein mixed breakfast meal. J. Nutr. Health Aging 2015, 19, 839–845. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. The vegan package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Korkmaz, S.; Goksuluk, D.; Zararsiz, G. MVN: An R package for assessing multivariate normality. R J. 2014, 6, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- González, I.; Déjean, S.; Martin, P.G.; Baccini, A. CCA: An R package to extend canonical correlation analysis. J. Stat. Softw. 2008, 23, 1–14. [Google Scholar] [CrossRef] [Green Version]
- González, I.; Cao, K.A.L.; Davis, M.J.; Déjean, S. Visualising associations between paired ‘omics’ data sets. BioData Min. 2012, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Lê Cao, K.-A.; González, I.; Déjean, S. IntegrOmics: An R package to unravel relationships between two omics datasets. Bioinformatics 2009, 25, 2855–2856. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, P.; Eugene, M.C.; Clarke, G.; Scully, P.; Barry, S.; Eamonn, M.M.Q.; Shanahan, F.; Cryan, J.; Timothy, G.D. Tryptophan catabolism in females with irritable bowel syndrome: Relationship to interferon-gamma, severity of symptoms and psychiatric co-morbidity. Neurogastroenterol. Motil. 2008, 20, 1291–1297. [Google Scholar] [CrossRef]
- Atkinson, W.; Lockhart, S.; Whorwell, P.; Keevil, B.; Houghton, L. Altered 5-Hydroxytryptamine Signaling in Patients With Constipation- and Diarrhea-Predominant Irritable Bowel Syndrome. Gastroenterology 2006, 130, 34–43. [Google Scholar] [CrossRef]
- Coates, M.D.; Mahoney, C.R.; Linden, D.R.; Sampson, J.E.; Chen, J.; Blaszyk, H.; Crowell, M.; Sharkey, K.; Gershon, M.D.; Mawe, G.M. Molecular defects in mucosal serotonin content and decreased serotonin reuptake transporter in ulcerative colitis and irritable bowel syndrome. Gastroenterology 2004, 126, 1657–1664. [Google Scholar] [CrossRef]
- Mujagic, Z.; Kasapi, M.; Jonkers, D.M.; Garcia-Perez, I.; Vork, L.; Weerts, Z.Z.R.; Serrano-Contreras, J.I.; Zhernakova, A.; Kurilshikov, A.; Scotcher, J.; et al. Integrated fecal microbiome–metabolome signatures reflect stress and serotonin metabolism in irritable bowel syndrome. Gut Microbes 2022, 14, 2063016. [Google Scholar] [CrossRef]
- Thijssen, A.Y.; Mujagic, Z.; Jonkers, D.M.A.E.; Ludidi, S.; Keszthelyi, D.; Hesselink, M.A.; Clemens, C.H.M.; Conchillo, J.M.; Kruimel, J.W.; Masclee, A.A.M. Alterations in serotonin metabolism in the irritable bowel syndrome. Aliment. Pharmacol. Ther. 2016, 43, 272–282. [Google Scholar] [CrossRef]
- Wang, W.W.; Qiao, S.Y.; Li, D.F. Amino acids and gut function. Amino Acids 2009, 37, 105–110. [Google Scholar] [CrossRef]
- Li, N.; Lewis, P.; Samuelson, D.; Liboni, K.; Neu, J. Glutamine regulates Caco-2 cell tight junction proteins. Am. J. Physiol. Liver Physiol. 2004, 287, G726–G733. [Google Scholar] [CrossRef] [Green Version]
- Coeffier, M.; Miralles-Barrachina, O.; Le Pessot, F.; Lalaude, O.; Daveau, M.; Lavoinne, A.; Lerebours, E.; Déchelotte, P. Influence of glutamine on cytokine production by human gut in vitro. Cytokine 2001, 13, 148–154. [Google Scholar] [CrossRef]
- Fernstrom, J.D. Large neutral amino acids: Dietary effects on brain neurochemistry and function. Amino Acids 2013, 45, 419–430. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Healthy Control | FC | FD | IBS-C | IBS-D | |
|---|---|---|---|---|---|
| Female count | 37 | 17 | 7 | 19 | 32 |
| Male count | 35 | 8 | 1 | 1 | 8 |
| Metabolite | Healthy Control | FC | FD | IBS-C | IBS-D |
|---|---|---|---|---|---|
| Neurotransmitters, nM | |||||
| Phenethylamine (PEA) | 2.0 ± 1.11 | 2.5 ± 1.51 | 2.0 ± 0.63 | 1.7 ± 0.68 | 2.1 ± 0.96 |
| 3,4-Dihydroxyphenylalanine (L-DOPA) | 28.5 ± 11.12 | 25.5 ± 9.11 | 25.3 ± 5.16 | 27.7 ± 7.61 | 26.6 ± 9.05 |
| Dopamine (DA) | 4.4 ± 7.61 | 5.5 ± 8.96 | 14.9 ± 29.09 | 5.9 ± 7.92 | 4.7 ± 6.49 |
| 3-Methoxytyramine (3-MT) | 0.6 ± 0.38 | 0.7 ± 0.56 | 1.1 ± 1.07 | 0.6 ± 0.64 | 0.6 ± 0.67 |
| 3,4-Dihydroxyphenylacetic acid (L-DOPAC) | 103.3 ± 65.42 | 116.2 ± 99.20 | 120.9 ± 38.60 | 165.4 ± 142.19 | 125.8 ± 103.86 |
| Homovanillic acid (HVA) | 140.5 ± 60.87 | 131.5 ± 72.71 | 158.5 ± 88.98 | 136.9 ± 65.50 | 149.8 ± 67 |
| Norepinephrine (NE) | 3.7 ± 1.34 | 3.7 ± 1.95 | 4.7 ± 2.08 | 3.7 ± 1.49 | 3.6 ± 1.48 |
| 3,4-Dihydroxyphenylglycol (DHPG) | 10.9 ± 3.00 | 10.9 ± 4.01 | 11.4 ± 3.44 | 11.2 ± 3.53 | 10.8 ± 2.95 |
| 3-Methoxy-4-hydroxyphenylglycol (MHPG) | 34.3 ± 7.19 | 33.4 ± 10.25 | 36.6 ± 7.83 | 33.2 ± 7.93 | 33.1 ± 7.75 |
| Normetanephrine (NM) | 2.1 ± 0.61 | 2.5 ± 1.43 | 2.3 ± 0.39 | 2.0 ± 0.83 | 2.0 ± 0.68 |
| Epinephrine (E) | 0.3 ± 0.20 | 0.2 ± 0.13 | 0.2 ± 0.11 | 0.2 ± 0.14 | 0.2 ± 0.21 |
| Metanephrine (MN) | 0.8 ± 0.20 | 0.8 ± 0.26 | 0.8 ± 0.18 | 0.7 ± 0.17 | 0.7 ± 0.17 |
| Vanillylmandelic acid (VMA) | 61.4 ± 18.72 | 63.2 ± 18.31 | 64.2 ± 22.9 | 59.9 ± 22.81 | 59.4 ± 19.98 |
| Kynurenine (KYN) | 8604.6 ± 2727.89 | 7700.1 ± 1601.01 | 7434.1 ± 2017.47 | 8439.3 ± 1392.45 | 8312.5 ± 2520.13 |
| 5-Hydroxytryptophan (5-HTP) | 49.3 ± 18.44 | 46.9 ± 23.27 | 45.7 ± 17.18 | 57.0 ± 21.24 | 48.0 ± 16.57 |
| Serotonin (5-HT) | 585.7 ± 565.47 | 544.0 ± 446.18 | 318.8 ± 377.10 | 620.6 ± 748.40 | 481.0 ± 426.04 |
| 5-Hydroxyindolacetic acid (5-HIAA) | 161.0 ± 125.69 | 176.7 ± 146.02 | 148.7 ± 111.75 | 166.9 ± 131.64 | 213.0 ± 220.91 |
| Alpha-aminobutyric acid (AABA) | 138.6 ± 57.90 | 139.7 ± 78.24 | 152.3 ± 58.71 | 159.7 ± 62.91 | 114.6 ± 42.94 |
| Gamma-aminobutyric acid (GABA) | 747.0 ± 230.49 | 746.1 ± 266.11 | 839.2 ± 321.81 | 794.2 ± 321.22 | 758.7 ± 281.63 |
| Amino acids (including non-proteogenic), μM | |||||
| Aspartic acid (ASP) | 6.0 ± 3.71 | 6.2 ± 3.31 | 6.1 ± 2.66 | 6.2 ± 3.90 | 6.1 ± 2.67 |
| Glutamic acid (GLU) | 54.5 ± 22.33 | 57.5 ± 24.42 | 43.5 ± 16.65 | 46.6 ± 25.90 | 58.1 ± 28.40 |
| Hydroxy-proline (HyPRO) * | 13.1 ± 5.15 | 11.4 ± 3.38 | 14.3 ± 3.10 | 11.9 ± 4.19 | 12.7 ± 3.94 |
| Serine (SER) | 113.3 ± 22.47 | 115.0 ± 19.25 | 124.5 ± 25.23 | 119.6 ± 23.75 | 116.4 ± 25.22 |
| Asparagine (ASN) | 56.7 ± 12.41 | 53.8 ± 10.78 | 54.4 ± 7.42 | 60.0 ± 10.58 | 56.6 ± 10.43 |
| Glycine (GLY) | 249.0 ± 60.87 | 258.7 ± 74.34 | 303.0 ± 90.04 | 246.4 ± 69.17 | 259.4 ± 58.67 |
| Glutamine (GLN) | 585.5 ± 77.60 | 607.7 ± 69.08 | 607.8 ± 49.99 | 584.3 ± 83.22 | 601.9 ± 69.84 |
| Histidine (HIS) | 59.3 ± 9.70 | 57.2 ± 7.91 | 58.3 ± 8.02 | 60.5 ± 8.91 | 61.5 ± 10.26 |
| Taurine (TAU) *, ** | 127.2 ± 44.00 | 133.7 ± 51.34 | 148.9 ± 62.27 | 140.9 ± 54.92 | 140.3 ± 49.42 |
| Citrulline (CIT) * | 35.3 ± 7.99 | 37.6 ± 10.54 | 35.0 ± 5.22 | 35.7 ± 8.56 | 35.0 ± 8.05 |
| Threonine (THR) | 112.0 ± 23.80 | 116.4 ± 20.89 | 115.9 ± 18.48 | 125.9 ± 33.50 | 116.2 ± 29.05 |
| Alanine (ALA) | 402.5 ± 105.12 | 409.2 ± 87.15 | 392.1 ± 98.77 | 370.4 ± 85.40 | 425.3 ± 114.54 |
| Arginine (ARG) | 69.2 ± 16.73 | 68.8 ± 16.55 | 80.6 ± 21.49 | 68.6 ± 21.36 | 73.5 ± 21.88 |
| Proline (PRO) | 281.0 ± 90.15 | 291.8 ± 100.07 | 306.1 ± 89.29 | 249.8 ± 73.52 | 278.1 ± 107.55 |
| Tyrosine (TYR) | 73.8 ± 15.52 | 80.8 ± 18.78 | 67.4 ± 7.26 | 69.6 ± 16.99 | 75.9 ± 18.11 |
| Valine (VAL) | 255.3 ± 54.75 | 248.9 ± 38.53 | 224.4 ± 33.84 | 226.6 ± 53.71 | 251.7 ± 58.85 |
| Methionine (MET) | 31.1 ± 6.71 | 31.9 ± 5.45 | 28.4 ± 3.07 | 29.7 ± 7.69 | 30.6 ± 6.51 |
| Isoleucine (ILE) | 80.8 ± 25.83 | 75.6 ± 16.43 | 67.6 ± 18.93 | 67.9 ± 23.31 | 78.2 ± 30.87 |
| Leucine (LEU) | 144.4 ± 45.03 | 137.8 ± 28.95 | 124.5 ± 31.52 | 120.8 ± 40.24 | 137.5 ± 57.43 |
| Ornithine (ORN) * | 62.7 ± 22.92 | 63.4 ± 14.28 | 60.3 ± 17.81 | 50.5 ± 16.02 | 63.2 ± 19.51 |
| Phenylalanine (PHE) | 66.6 ± 9.73 | 66.2 ± 8.90 | 60.4 ± 7.22 | 64.5 ± 13.01 | 66.5 ± 10.58 |
| Lysine (LYS) | 94.8 ± 22.37 | 95.3 ± 19.46 | 96.5 ± 11.56 | 91.0 ± 22.23 | 94.0 ± 21.59 |
| Tryptophan (TRP) | 47.6 ± 15.86 | 46.7 ± 12.28 | 44.4 ± 16.30 | 46.5 ± 14.50 | 45.9 ± 17.62 |
| BCAAs (ILE, LEU, VAL) | 480.5 ± 125.60 | 462.3 ± 83.91 | 416.5 ± 84.30 | 415.3 ± 117.27 | 467.4 ± 147.15 |
| EAAs (HIS, THR, VAL, MET, ILE, LEU, PHE, LYS, TRP) | 891.8 ± 213.76 | 876.0 ± 158.80 | 820.4 ± 148.94 | 833.4 ± 217.11 | 882.1 ± 242.78 |
| NEAAs (ALA, ARG, ASN, ASP, GLU, GLN, GLY, PRO, SER, TYR) | 1891.6 ± 426.93 | 1949.4 ± 423.72 | 1985.4 ± 408.80 | 1821.3 ± 413.79 | 1951.3 ± 457.32 |
| LNAAs (HIS, THR, VAL, MET, ILE, LEU, PHE, TYR, TRP) | 870.8 ± 206.92 | 861.4 ± 158.13 | 791.3 ± 144.65 | 812.0 ± 211.88 | 864.0 ± 239.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, S.C.; Fraser, K.; Cooney, J.; Günther, C.S.; Young, W.; Gearry, R.B.; Heenan, P.E.; Trower, T.; Keenan, J.I.; Talley, N.J.; et al. Concentrations of Plasma Amino Acids and Neurotransmitters in Participants with Functional Gut Disorders and Healthy Controls. Metabolites 2023, 13, 313. https://doi.org/10.3390/metabo13020313
James SC, Fraser K, Cooney J, Günther CS, Young W, Gearry RB, Heenan PE, Trower T, Keenan JI, Talley NJ, et al. Concentrations of Plasma Amino Acids and Neurotransmitters in Participants with Functional Gut Disorders and Healthy Controls. Metabolites. 2023; 13(2):313. https://doi.org/10.3390/metabo13020313
Chicago/Turabian StyleJames, Shanalee C., Karl Fraser, Janine Cooney, Catrin S. Günther, Wayne Young, Richard B. Gearry, Phoebe E. Heenan, Tania Trower, Jacqueline I. Keenan, Nicholas J. Talley, and et al. 2023. "Concentrations of Plasma Amino Acids and Neurotransmitters in Participants with Functional Gut Disorders and Healthy Controls" Metabolites 13, no. 2: 313. https://doi.org/10.3390/metabo13020313
APA StyleJames, S. C., Fraser, K., Cooney, J., Günther, C. S., Young, W., Gearry, R. B., Heenan, P. E., Trower, T., Keenan, J. I., Talley, N. J., McNabb, W. C., & Roy, N. C. (2023). Concentrations of Plasma Amino Acids and Neurotransmitters in Participants with Functional Gut Disorders and Healthy Controls. Metabolites, 13(2), 313. https://doi.org/10.3390/metabo13020313