
Intestinal Immune Deficiency and Juvenile Hormone Signaling Mediate a Metabolic Trade-off in Adult Drosophila Females


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
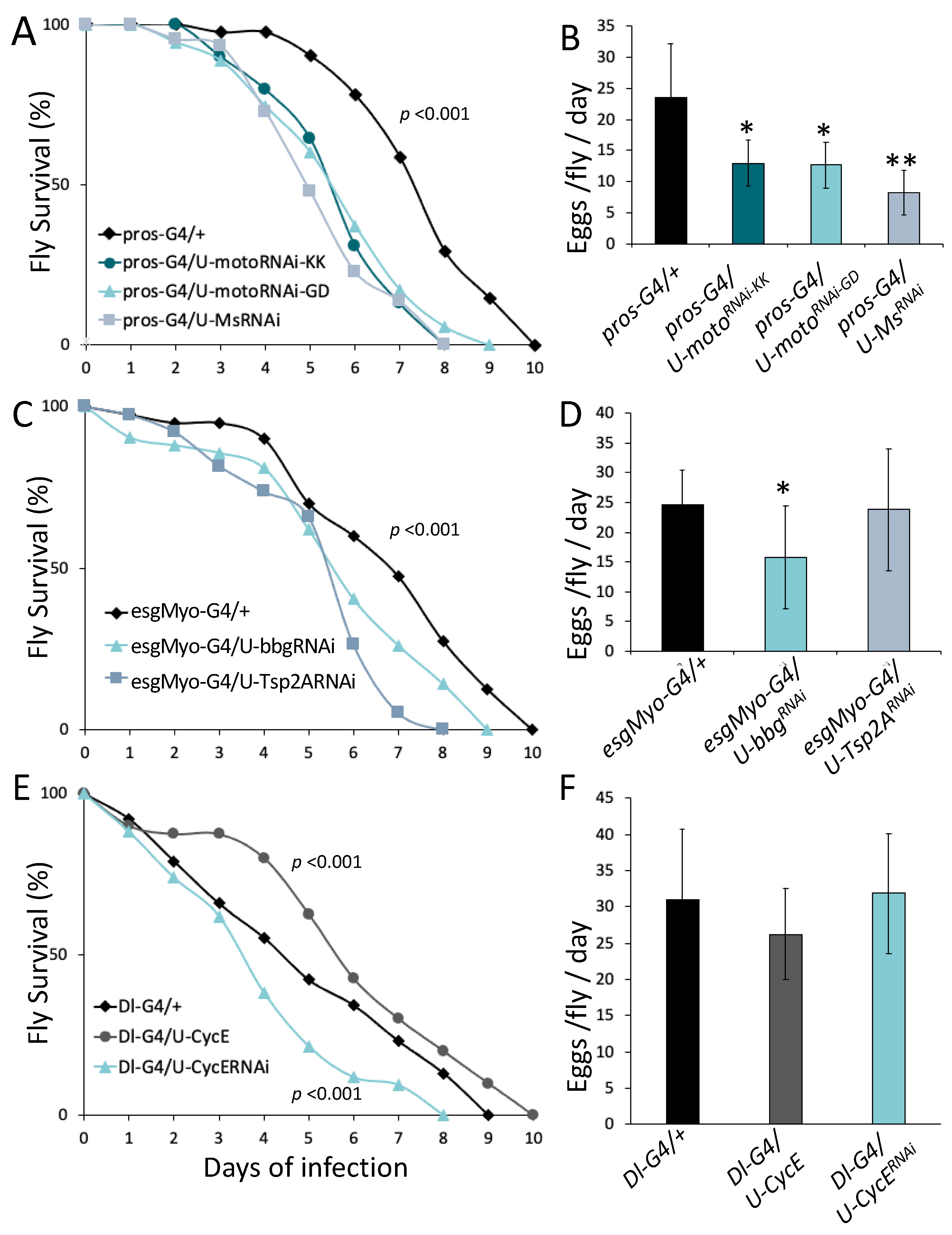
2.1. Immune Activation Reduces Reproductive Rate
2.2. Imd and Duox Signaling in Midgut Enterocytes Compromises Fecundity and Insulin-Mediated Metabolism
2.3. Mating Compromises Female Drosophila Intestinal Host Defense
2.4. JH Synthesis and Signaling Compromises Intestinal Host Defense
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction–Immunity Trade-Offs in Insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuraishi, T.; Hori, A.; Kurata, S. Host-Microbe Interactions in the Gut of Drosophila melanogaster. Front. Physiol. 2013, 4, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Chamy, L.; Matt, N.; Ntwasa, M.; Reichhart, J.-M. The Multilayered Innate Immune Defense of the Gut. Biomed. J. 2015, 38, 276. [Google Scholar] [CrossRef]
- Ha, E.-M.; Oh, C.-T.; Bae, Y.S.; Lee, W.-J. A Direct Role for Dual Oxidase in Drosophila Gut Immunity. Science 2005, 310, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Poidevin, M.; Pradervand, S.; Lemaitre, B. Drosophila Intestinal Response to Bacterial Infection: Activation of Host Defense and Stem Cell Proliferation. Cell Host Microbe 2009, 5, 200–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New Insights into Peritrophic Matrix Synthesis, Architecture, and Function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef]
- Kuraishi, T.; Binggeli, O.; Opota, O.; Buchon, N.; Lemaitre, B. Genetic Evidence for a Protective Role of the Peritrophic Matrix against Intestinal Bacterial Infection in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2011, 108, 15966–15971. [Google Scholar] [CrossRef] [Green Version]
- Hultmark, D. Insect lysozymes. EXS 1996, 75, 87–102. [Google Scholar] [CrossRef]
- LaJeunesse, D.R.; Johnson, B.; Presnell, J.S.; Catignas, K.; Zapotoczny, G. Peristalsis in the Junction Region of the Drosophila Larval Midgut Is Modulated by DH31 Expressing Enteroendocrine Cells. BMC Physiol. 2010, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.; Micchelli, C.A. Quiescent Gastric Stem Cells Maintain the Adult Drosophila Stomach. Proc. Natl. Acad. Sci. USA 2011, 108, 17696–17701. [Google Scholar] [CrossRef] [Green Version]
- Opota, O.; Vallet-Gély, I.; Vincentelli, R.; Kellenberger, C.; Iacovache, I.; Gonzalez, M.R.; Roussel, A.; van der Goot, F.-G.; Lemaitre, B. Monalysin, a Novel ß-Pore-Forming Toxin from the Drosophila Pathogen Pseudomonas Entomophila, Contributes to Host Intestinal Damage and Lethality. PLoS Pathog. 2011, 7, e1002259. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-Z.; Lestradet, M.; Socha, C.; Schirmeier, S.; Schmitz, A.; Spenlé, C.; Lefebvre, O.; Keime, C.; Yamba, W.M.; Bou Aoun, R.; et al. Enterocyte Purge and Rapid Recovery Is a Resilience Reaction of the Gut Epithelium to Pore-Forming Toxin Attack. Cell Host Microbe 2016, 20, 716–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, T.; Maki, K.; Hadano, J.; Fujikawa, T.; Kitazaki, K.; Koshiba, T.; Kawabata, S. Crosslinking of a Peritrophic Matrix Protein Protects Gut Epithelia from Bacterial Exotoxins. PLoS Pathog. 2015, 11, e1005244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apidianakis, Y.; Pitsouli, C.; Perrimon, N.; Rahme, L. Synergy between Bacterial Infection and Genetic Predisposition in Intestinal Dysplasia. Proc. Natl. Acad. Sci. USA 2009, 106, 20883–20888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Patel, P.H.; Kohlmaier, A.; Grenley, M.O.; McEwen, D.G.; Edgar, B.A. Cytokine/Jak/Stat Signaling Mediates Regeneration and Homeostasis in the Drosophila Midgut. Cell 2009, 137, 1343–1355. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hodgson, J.J.; Buchon, N. Drosophila as a Model for Homeostatic, Antibacterial, and Antiviral Mechanisms in the Gut. PLoS Pathog. 2017, 13, e1006277. [Google Scholar] [CrossRef] [Green Version]
- Biteau, B.; Hochmuth, C.E.; Jasper, H. JNK Activity in Somatic Stem Cells Causes Loss of Tissue Homeostasis in the Aging Drosophila Gut. Cell Stem Cell 2008, 3, 442–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamamouna, V.; Panagi, M.; Theophanous, A.; Demosthenous, M.; Michail, M.; Papadopoulou, M.; Teloni, S.; Pitsouli, C.; Apidianakis, Y. Evidence of Two Types of Balance between Stem Cell Mitosis and Enterocyte Nucleus Growth in the Drosophila Midgut. Development 2020, 147, dev189472. [Google Scholar] [CrossRef]
- Vandehoef, C.; Molaei, M.; Karpac, J. Dietary Adaptation of Microbiota in Drosophila Requires NF-ΚB-Dependent Control of the Translational Regulator 4E-BP. Cell Rep. 2020, 31, 107736. [Google Scholar] [CrossRef]
- Bahuguna, S.; Atilano, M.; Glittenberg, M.; Lee, D.; Arora, S.; Wang, L.; Zhou, J.; Redhai, S.; Boutros, M.; Ligoxygakis, P. Bacterial Recognition by PGRP-SA and Downstream Signalling by Toll/DIF Sustain Commensal Gut Bacteria in Drosophila. PLoS Genet. 2022, 18, e1009992. [Google Scholar]
- Raikhel, A.S.; Brown, M.R.; Belles, X. Hormonal Control and Reproduvtie Proceses. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Chapter 3.9; pp. 433–491. [Google Scholar]
- Toivonen, J.M.; Partridge, L. Endocrine Regulation of Aging and Reproduction in Drosophila. Mol. Cell. Endocrinol. 2009, 299, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Rivera-Perez, C.; Abdou, M.; Jia, Q.; He, Q.; Liu, X.; Zyaan, O.; Xu, J.; Bendena, W.G.; Tobe, S.S.; et al. Methyl Farnesoate Plays a Dual Role in Regulating Drosophila Metamorphosis. PLoS Genet. 2015, 11, e1005038. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Karpova, E.K.; Adonyeva, N.V.; Andreenkova, O.V.; Burdina, E.V.; Ilinsky, Y.Y.; Bykov, R.A.; Menshanov, P.N.; Rauschenbach, I.Y. Drosophila Female Fertility and Juvenile Hormone Metabolism Depends on the Type of Wolbachia Infection. J. Exp. Biol. 2019, 222, jeb.195347. [Google Scholar] [CrossRef] [Green Version]
- Hansen, I.A.; Attardo, G.M.; Rodriguez, S.D.; Drake, L.L. Four-Way Regulation of Mosquito Yolk Protein Precursor Genes by Juvenile Hormone-, Ecdysone-, Nutrient-, and Insulin-like Peptide Signaling Pathways. Front. Physiol. 2014, 5, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badisco, L.; van Wielendaele, P.; vanden Broeck, J. Eat to Reproduce: A Key Role for the Insulin Signaling Pathway in Adult Insects. Front. Physiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.M.; Baggott, S.L.; Maingon, R.; Hurd, H. The Costs of Mounting an Immune Response Are Reflected in the Reproductive Fitness of the Mosquito Anopheles gambiae. Oikos 2002, 97, 371–377. [Google Scholar]
- Kurz, C.L.; Charroux, B.; Chaduli, D.; Viallat-Lieutaud, A.; Royet, J. Peptidoglycan Sensing by Octopaminergic Neurons Modulates Drosophila Oviposition. Elife 2017, 6, e21937. [Google Scholar] [CrossRef]
- Kotronarou, K.; Charalambous, A.; Evangelou, A.; Georgiou, O.; Demetriou, A.; Apidianakis, Y. Dietary Stimuli, Intestinal Bacteria and Peptide Hormones Regulate Female Drosophila Defecation Rate. Metabolites 2023, 13, 264. [Google Scholar] [CrossRef]
- Bonnay, F.; Cohen-Berros, E.; Hoffmann, M.; Kim, S.Y.; Boulianne, G.L.; Hoffmann, J.A.; Matt, N.; Reichhart, J.-M. Big Bang Gene Modulates Gut Immune Tolerance in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 2957–2962. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Tang, H.-W.; Hung, R.-J.; Hu, Y.; Ni, X.; Housden, B.E.; Perrimon, N. The Septate Junction Protein Tsp2A Restricts Intestinal Stem Cell Activity via Endocytic Regulation of APKC and Hippo Signaling. Cell Rep. 2019, 26, 670–688.e6. [Google Scholar] [CrossRef] [Green Version]
- Schwenke, R.A.; Lazzaro, B.P. Juvenile Hormone Suppresses Resistance to Infection in Mated Female Drosophila melanogaster. Curr. Biol. 2017, 27, 596–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiAngelo, J.R.; Bland, M.L.; Bambina, S.; Cherry, S.; Birnbaum, M.J. The Immune Response Attenuates Growth and Nutrient Storage in Drosophila by Reducing Insulin Signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 20853–20858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, S.; Purdy, A.E.; Robins, W.P.; Wang, Z.; Mandal, M.; Chang, S.; Mekalanos, J.J.; Watnick, P.I. The Acetate Switch of an Intestinal Pathogen Disrupts Host Insulin Signaling and Lipid Metabolism. Cell Host Microbe 2014, 16, 592–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjieconomou, D.; King, G.; Gaspar, P.; Mineo, A.; Blackie, L.; Ameku, T.; Studd, C.; de Mendoza, A.; Diao, F.; White, B.H.; et al. Enteric Neurons Increase Maternal Food Intake during Reproduction. Nature 2020, 587, 455–459. [Google Scholar] [CrossRef]
- Rahman, M.M.; Franch-Marro, X.; Maestro, J.L.; Martin, D.; Casali, A. Local Juvenile Hormone Activity Regulates Gut Homeostasis and Tumor Growth in Adult Drosophila. Sci. Rep. 2017, 7, 11677. [Google Scholar] [CrossRef] [Green Version]
- Reiff, T.; Jacobson, J.; Cognigni, P.; Antonello, Z.; Ballesta, E.; Tan, K.J.; Yew, J.Y.; Dominguez, M.; Miguel-Aliaga, I. Endocrine Remodelling of the Adult Intestine Sustains Reproduction in Drosophila. Elife 2015, 4, e06930. [Google Scholar] [CrossRef] [Green Version]
- Evangelou, A.; Ignatiou, A.; Antoniou, C.; Kalanidou, S.; Chatzimatthaiou, S.; Shianiou, G.; Ellina, S.; Athanasiou, R.; Panagi, M.; Apidianakis, Y.; et al. Unpredictable Effects of the Genetic Background of Transgenic Lines in Physiological Quantitative Traits. G3 Genes Genomes Genet. 2019, 9, 3877–3890. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.M.; Engström, Y. Immune Response in the Barrier Epithelia: Lessons from the Fruit Fly Drosophila melanogaster. J. Innate Immun. 2012, 4, 273–283. [Google Scholar] [CrossRef]
- Veenstra, J.A.; Agricola, H.-J.; Sellami, A. Regulatory Peptides in Fruit Fly Midgut. Cell Tissue Res. 2008, 334, 499–516. [Google Scholar] [CrossRef]
- Dutta, D.; Dobson, A.J.; Houtz, P.L.; Gläßer, C.; Revah, J.; Korzelius, J.; Patel, P.H.; Edgar, B.A.; Buchon, N. Regional Cell-Specific Transcriptome Mapping Reveals Regulatory Complexity in the Adult Drosophila Midgut. Cell Rep. 2015, 12, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Apidianakis, Y.; Mindrinos, M.N.; Xiao, W.; Lau, G.W.; Baldini, R.L.; Davis, R.W.; Rahme, L.G. Profiling Early Infection Responses: Pseudomonas Aeruginosa Eludes Host Defenses by Suppressing Antimicrobial Peptide Gene Expression. Proc. Natl. Acad. Sci. USA 2005, 102, 2573–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangi, E.; Pitsouli, C.; Rahme, L.G.; Cagan, R.; Apidianakis, Y. Immune Response to Bacteria Induces Dissemination of Ras-activated Drosophila Hindgut Cells. EMBO Rep. 2012, 13, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Panayidou, S.; Georgiades, K.; Christofi, T.; Tamana, S.; Promponas, V.J.; Apidianakis, Y. Pseudomonas Aeruginosa Core Metabolism Exerts a Widespread Growth-Independent Control on Virulence. Sci. Rep. 2020, 10, 9505. [Google Scholar] [CrossRef]
- Kounatidis, I.; Chtarbanova, S.; Cao, Y.; Hayne, M.; Jayanth, D.; Ganetzky, B.; Ligoxygakis, P. NF-ΚB Immunity in the Brain Determines Fly Lifespan in Healthy Aging and Age-Related Neurodegeneration. Cell Rep 2017, 19, 836–848. [Google Scholar] [CrossRef]
- Cao, Y.; Chtarbanova, S.; Petersen, A.J.; Ganetzky, B. Dnr1 Mutations Cause Neurodegeneration in Drosophila by Activating the Innate Immune Response in the Brain. Proc. Natl. Acad. Sci. USA 2013, 110, E1752–E1760. [Google Scholar] [CrossRef] [Green Version]
- White, M.A.; Bonfini, A.; Wolfner, M.F.; Buchon, N. Drosophila melanogaster Sex Peptide Regulates Mated Female Midgut Morphology and Physiology. Proc. Natl. Acad. Sci. USA 2021, 118, e2018112118. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.-H.; Kim, J.-G.; Yang, D.-J.; Kim, Y.-S.; Yoo, M.-A. Age-Related Changes in Drosophila Midgut Are Associated with PVF2, a PDGF/VEGF-like Growth Factor. Aging Cell 2008, 7, 318–334. [Google Scholar] [CrossRef] [Green Version]
- Short, S.M.; Wolfner, M.F.; Lazzaro, B.P. Female Drosophila melanogaster Suffer Reduced Defense against Infection Due to Seminal Fluid Components. J. Insect Physiol. 2012, 58, 1192–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubli, E. Sexual Behavior: Dietary Food Switch Induced by Sex. Curr. Biol. 2010, 20, R474–R476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahme, L.G.; Stevens, E.J.; Wolfort, S.F.; Shao, J.; Tompkins, R.G.; Ausubel, F.M.; Galaktionov, K.; Lee, A.; Eckstein, J.; Draetta, G.; et al. Common virulence factors for bacterial pathogenicity in plants and animals. Science 1995, 268, 1899–1902. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shianiou, G.; Teloni, S.; Apidianakis, Y. Intestinal Immune Deficiency and Juvenile Hormone Signaling Mediate a Metabolic Trade-off in Adult Drosophila Females. Metabolites 2023, 13, 340. https://doi.org/10.3390/metabo13030340
Shianiou G, Teloni S, Apidianakis Y. Intestinal Immune Deficiency and Juvenile Hormone Signaling Mediate a Metabolic Trade-off in Adult Drosophila Females. Metabolites. 2023; 13(3):340. https://doi.org/10.3390/metabo13030340
Chicago/Turabian StyleShianiou, Gavriella, Savvas Teloni, and Yiorgos Apidianakis. 2023. "Intestinal Immune Deficiency and Juvenile Hormone Signaling Mediate a Metabolic Trade-off in Adult Drosophila Females" Metabolites 13, no. 3: 340. https://doi.org/10.3390/metabo13030340