


Metabolomics-Based Analysis of the Effects of Different Cultivation Strategies on Metabolites of Dendrobium officinale Kimura et Migo

Abstract
1. Introduction
2. Results
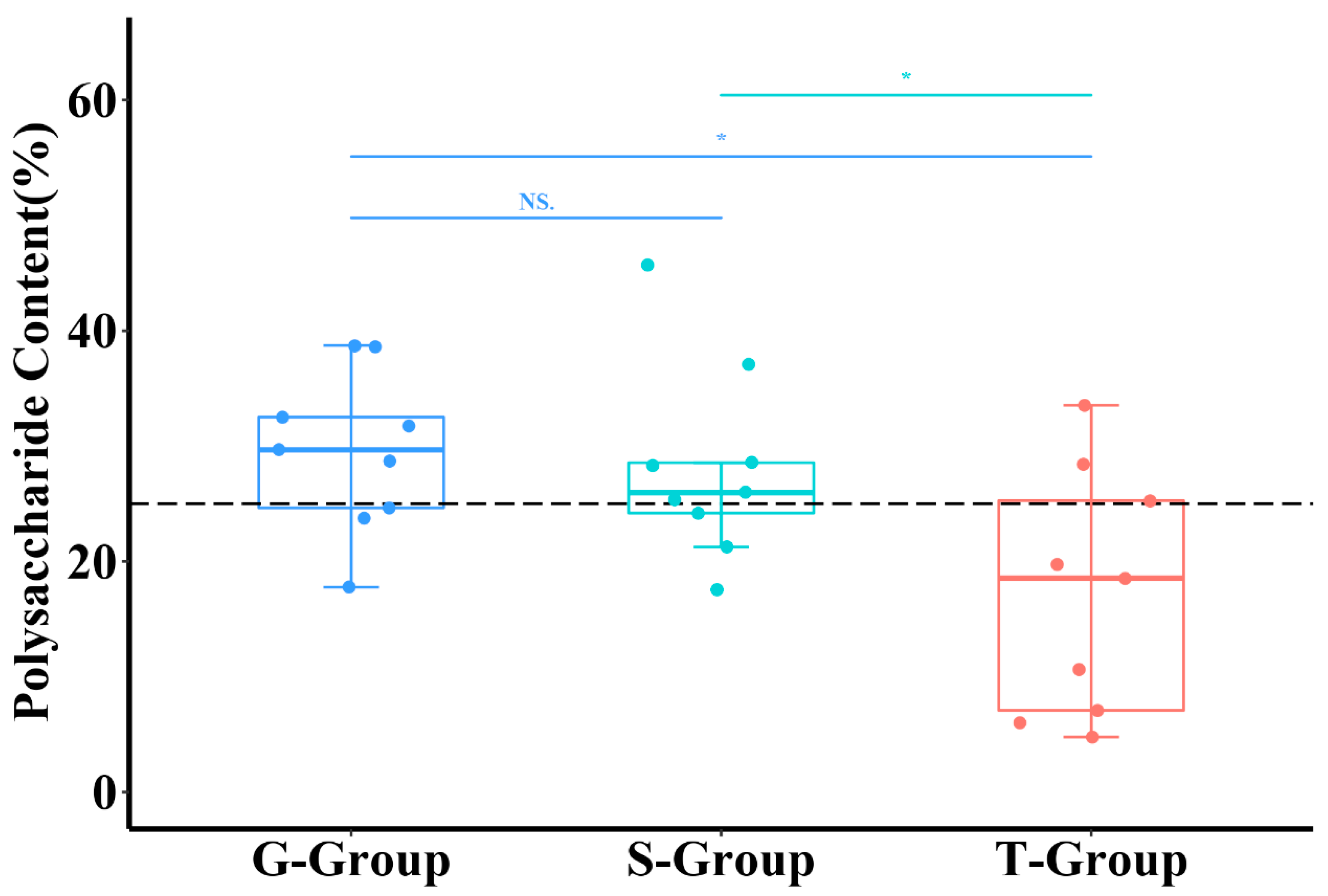
2.1. Effects of Different Cultivation Methods on the Polysaccharide Content of D. officinale
2.2. Metabolomic Determination of D. officinale by UPLC-MS
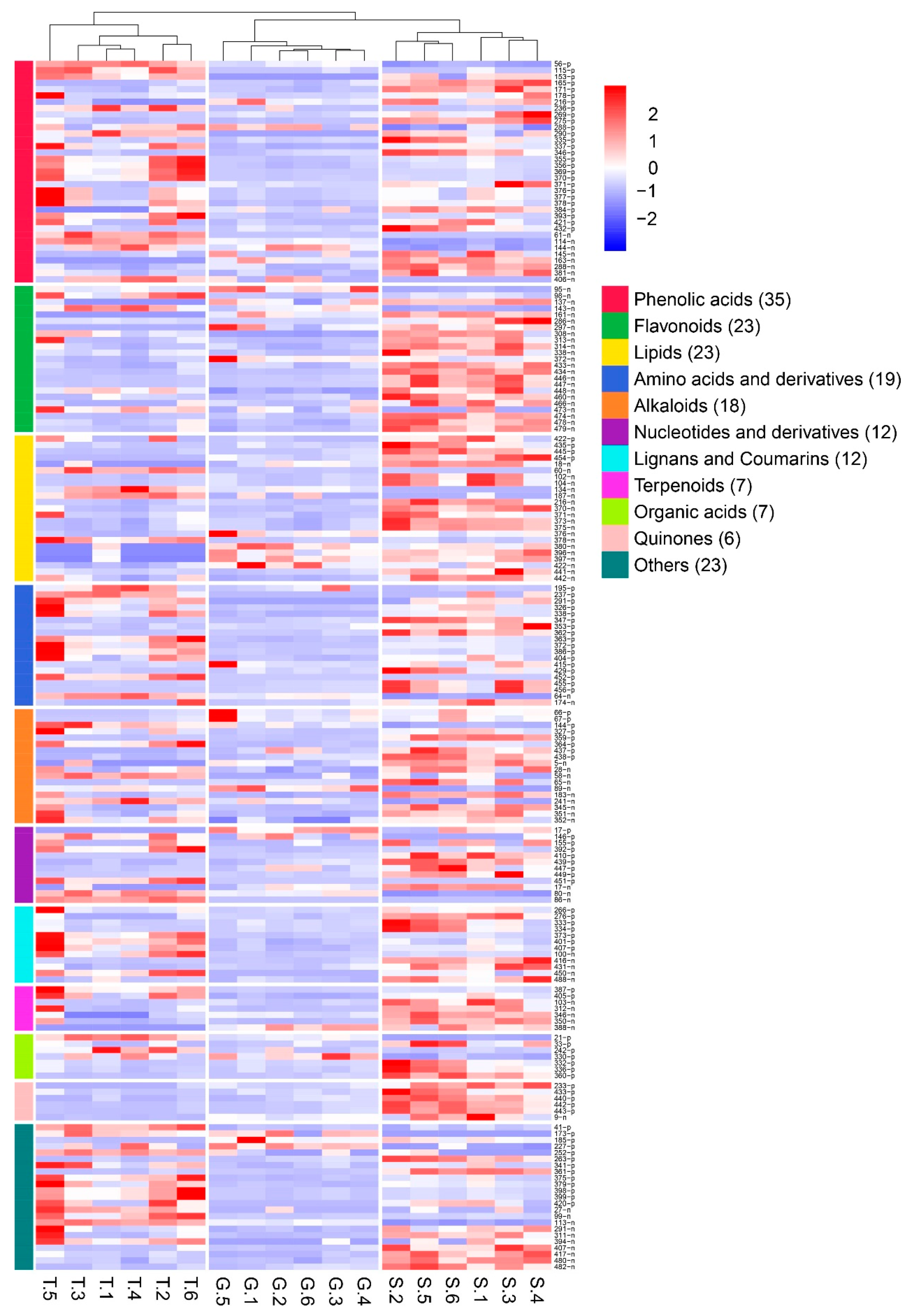
2.3. Effects of Different Cultivation Methods on the Metabolite Profile of D. officinale
2.4. Effects of Different Cultivation Methods on the Metabolites of D. officinale
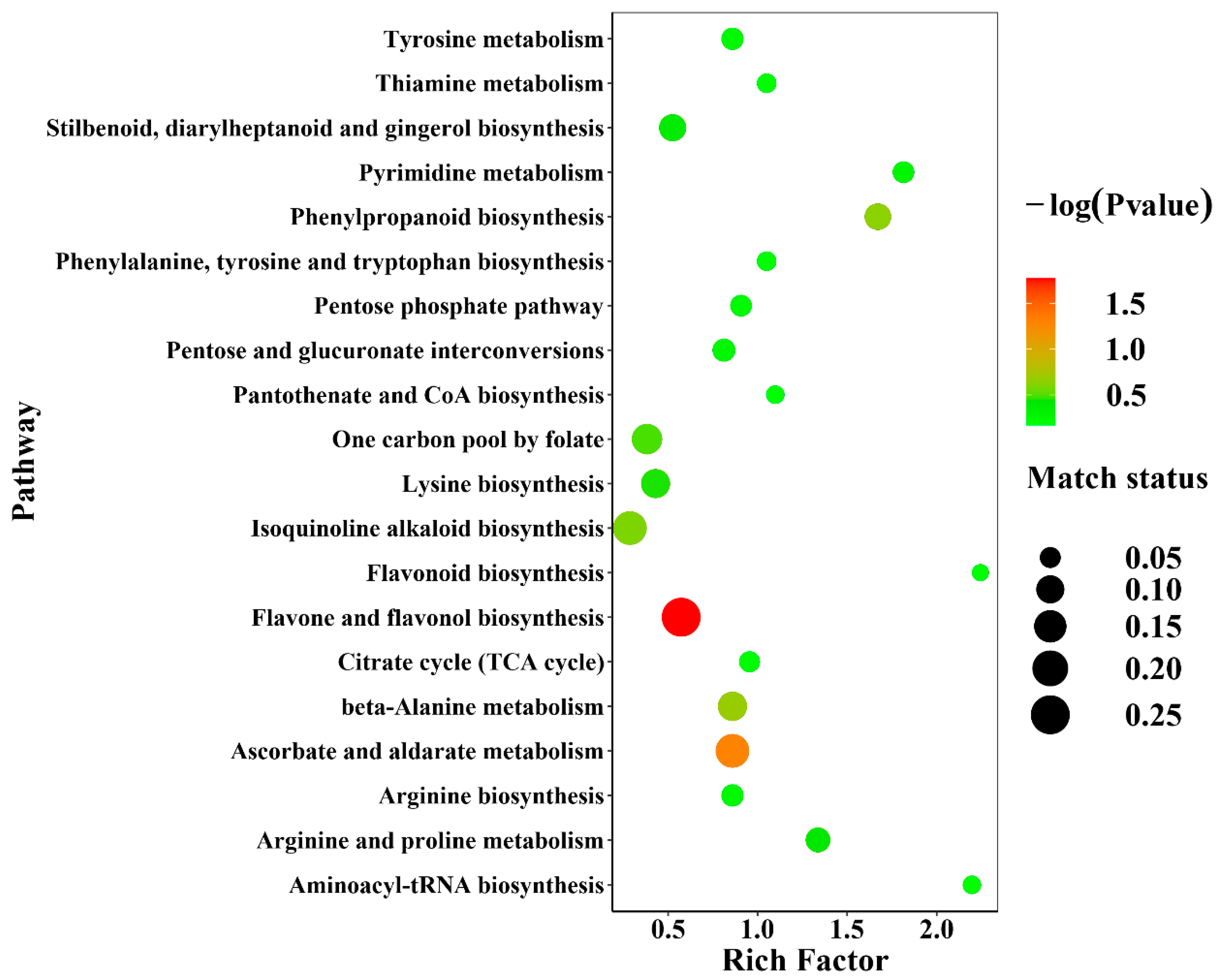
2.5. Effects of Different Cultivation Methods on the Metabolic Pathways of D. officinale
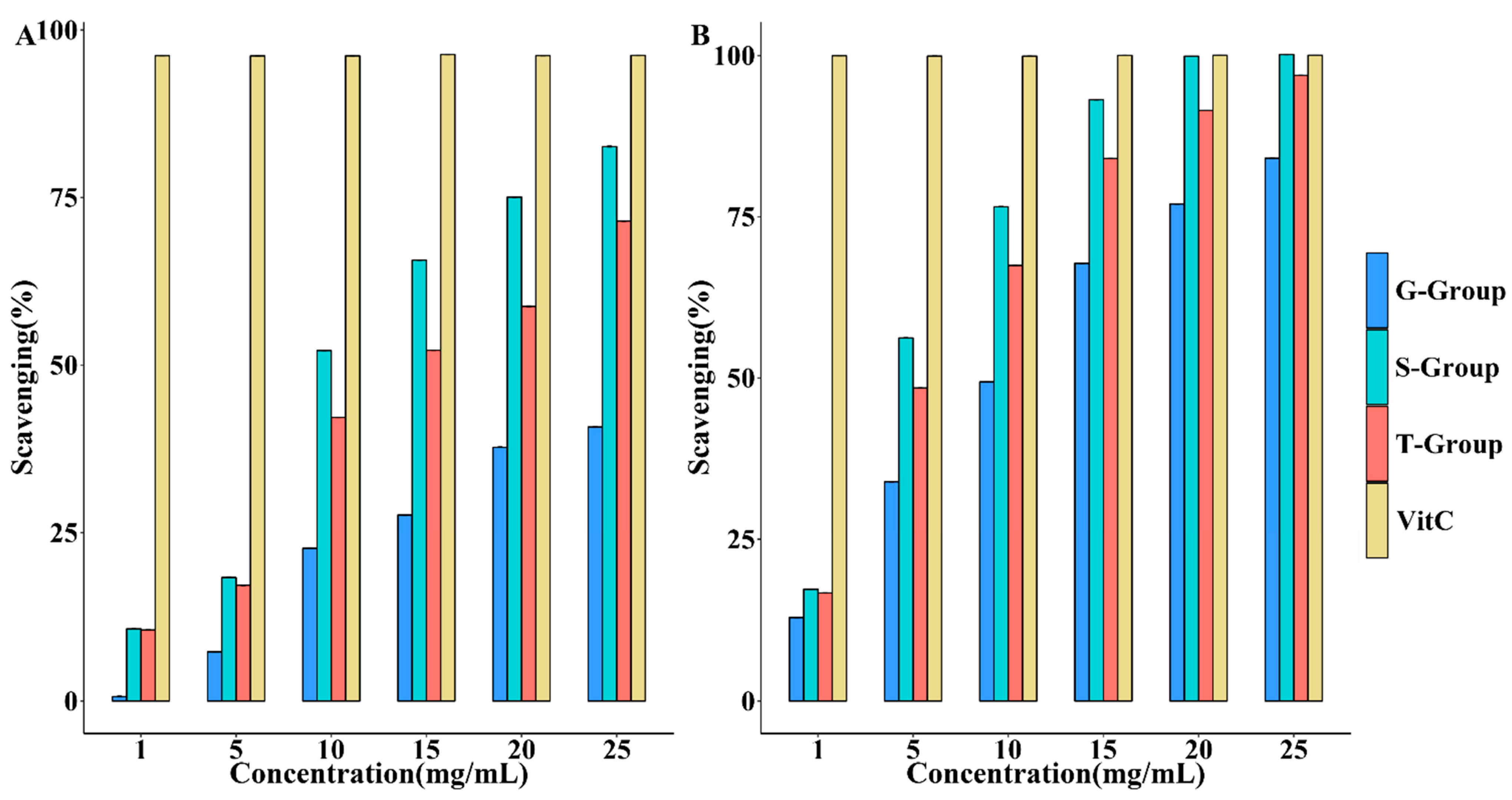
2.6. Effects of Different Cultivation Methods on the Antioxidant Activity of D. officinale
3. Discussion
4. Materials and Methods
4.1. Collection Information of D. officinale Samples
4.2. Instruments and Reagents
4.3. Determination of Polysaccharide Content of D. officinale
4.3.1. Extraction of Polysaccharide
4.3.2. Polysaccharide Assay
4.3.3. Preparation of Glucose Standard Curve
4.4. Determination of the Metabolomics of D. officinale by UPLC -MS/MS
4.5. KEGG Metabolic Pathway Enrichment
4.6. Comparing the Antioxidant Properties of D. officinale Samples
4.6.1. Sample Extraction
4.6.2. DPPH Radical Scavenging Assay
4.6.3. ABTS Radical Scavenging Assay
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.Y.; Guo, Y.; Si, J.P.; Sun, X.B.; Sun, G.B.; Liu, J.J. A polysaccharide of Dendrobium officinale ameliorates H2O2 -induced apoptosis in H9c2 cardiomyocytes via PI3K/AKT and MAPK pathways. Int. J. Biol. Macromol. 2017, 104, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, T.; Yan, M.Q.; Yu, J.J.; Xu, J.B.; Zhuge, R.C.; Yu, H.H.; Chen, S.H.; Lyu, G.Y. Effect of Dendrobii officinalis superfine powder on overeating greasy-induced metabolic hypertension in rats. Zhongguo Zhong Yao Za Zhi = China J. Chin. Mater. Med 2021, 46, 1658–1666. [Google Scholar] [CrossRef]
- Yang, J.; Chen, H.; Nie, Q.; Huang, X.; Nie, S. Dendrobium officinale polysaccharide ameliorates the liver metabolism disorders of type II diabetic rats. Int. J. Biol. Macromol. 2020, 164, 1939–1948. [Google Scholar] [CrossRef]
- Zeng, J.; Li, D.N.; Li, Z.B.; Zhang, J.; Zhao, X.Y. Dendrobium officinale attenuates myocardial fibrosis via inhibiting EMT signaling pathway in HFD/STZ-induced diabetic mice. Biol. Pharm. Bull. 2020, 43, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liang, J.; Zhong, Y.; Xiao, G.; Efferth, T.; Georgiev, M.; Vargas-De-La-Cruz, C.; Bajpai, V.; Caprioli, G.; Liu, J.; et al. Dendrobium officinale Polysaccharide Alleviates Intestinal Inflammation by Promoting Small Extracellular Vesicle Packaging of miR-433-3p. J. Agric. Food Chem. 2021, 69, 13510–13523. [Google Scholar] [CrossRef]
- Liu, C.Z.; Chen, W.; Wang, M.X.; Wang, Y.; Chen, L.Q.; Zhao, F.; Shi, Y.; Liu, H.J.; Dou, X.B.; Liu, C. Dendrobium officinale Kimura et Migo and American ginseng mixture: A Chinese herbal formulation for gut microbiota modulation. Chin. J. Nat. Med. 2020, 18, 446–459. [Google Scholar] [CrossRef]
- Li, L.; Yao, H.; Li, X.; Zhang, Q.; Wu, X.; Wong, T.; Zheng, H.; Fung, H.; Yang, B.; Ma, D.; et al. Destiny of Dendrobium officinale Polysaccharide after Oral Administration: Indigestible and Nonabsorbing, Ends in Modulating Gut Microbiota. J. Agric. Food Chem. 2019, 67, 5968–5977. [Google Scholar] [CrossRef]
- He, T.-B.; Huang, Y.-P.; Yang, L.; Liu, T.-T.; Gong, W.-Y.; Wang, X.-J.; Sheng, J.; Hu, J.-M. Structural characterization and immunomodulating activity of polysaccharide from Dendrobium officinale. Int. J. Biol. Macromol. 2016, 83, 34–41. [Google Scholar] [CrossRef]
- Huang, Y.P.; He, T.B.; Cuan, X.D.; Wang, X.J.; Hu, J.M.; Sheng, J. 1, 4-β-d-Glucomannan from Dendrobium officinale activates NF-кB via TLR4 to regulate the immune response. Molecules 2018, 23, 2658. [Google Scholar] [CrossRef]
- Zhang, K.; Zhou, X.; Wang, J.; Zhou, Y.; Qi, W.; Chen, H.; Nie, S.; Xie, M. Dendrobium officinale polysaccharide triggers mitochondrial disorder to induce colon cancer cell death via ROS-AMPK-autophagy pathway. Carbohydr. Polym. 2021, 264, 118018. [Google Scholar] [CrossRef]
- Guo, Z.; Zhou, Y.; Yang, J.; Shao, X. Dendrobium candidum extract inhibits proliferation and induces apoptosis of liver cancer cells by inactivating Wnt/β-catenin signaling pathway. Biomed. Pharmacother. 2019, 110, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.P.; Yu, W.X.; Zhang, X.F.; Luo, Y.Y.; Lei, Z.X.; Huang, D.D.; Lin, J.; Huang, Y.C.; Huang, S.W.; Nong, F.F.; et al. Isoviolanthin Extracted from Dendrobium officinale Reverses TGF-β1-Mediated Epithelial–Mesenchymal Transition in Hepatocellular Carcinoma Cells via Deactivating the TGF-β/Smad and PI3K/Akt/mTOR Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 1556. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yang, T.; Shen, L.; Zhang, J.; Liu, L. Study on the properties of Dendrobiumofficinale fermentation broth as functional raw material of cosmetics. J. Cosmet. Dermatol. 2022, 21, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Yang, K.; Zhan, L.; Lu, T.; Chen, X.; Cai, X.; Zhou, C.; Li, H.; Qian, L.; Lv, G.; et al. Optimization of Flavonoid Extraction in Dendrobium officinale Leaves and Their Inhibitory Effects on Tyrosinase Activity. Int. J. Anal. Chem. 2019, 2019, 7849198. [Google Scholar] [CrossRef]
- Wang, Q.; Zi, C.T.; Wang, J.; Wang, Y.N.; Huang, Y.W.; Fu, X.Q.; Wang, X.J.; Sheng, J. Dendrobium officinale Orchid Extract Prevents Ovariectomy-Induced Osteoporosis in Vivo and Inhibits RANKL-Induced Osteoclast Differentiation in Vitro. Front. Pharmacol. 2017, 8, 966. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, K.; Jiang, Q.; Dai, D.; Li, H.; Bi, W.; Chen, D.D.Y. Characterization of plant polysaccharides from Dendrobium officinale by multiple chromatographic and mass spectrometric techniques. J. Chromatogr. A 2018, 1547, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Xie, Z.; Lei, Z.; Huang, Y.; Wei, G. Simultaneous identification and determination of flavonoids in Dendrobium officinale. Chem. Cent. J. 2018, 12, 40. [Google Scholar] [CrossRef]
- Yoo, S.R.; Jeong, S.J.; Lee, N.R.; Shin, H.K.; Seo, C.S. Simultaneous determination and anti-inflammatory effects of four phenolic compounds in Dendrobii Herba. Nat. Prod. Res. 2017, 31, 2923–2926. [Google Scholar] [CrossRef]
- Liu, J.L.; Yu, N.J.; Xing, L.H.; Wang, R.; Xu, J.; Peng, D.Y.; Tong, X.H.; Ou, J.M.; He, X.L. Simultaneous determination and analysis of amino acids and nucleosides in Dendrobium by UHPLC-QTRAP-MS/MS. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2020, 45, 3890–3899. [Google Scholar] [CrossRef]
- He, L.; Su, Q.; Bai, L.; Li, M.; Liu, J.; Liu, X.; Zhang, C.; Jiang, Z.; He, J.; Shi, J.; et al. Recent research progress on natural small molecule bibenzyls and its derivatives in Dendrobium species. Eur. J. Med. Chem. 2020, 204, 112530. [Google Scholar] [CrossRef]
- Song, C.; Zhang, Y.; Manzoor, M.A.; Li, G. Identification of alkaloids and related intermediates of Dendrobium officinale by solid-phase extraction coupled with high-performance liquid chromatography tandem mass spectrometry. Front. Plant Sci. 2022, 13, 952051. [Google Scholar] [CrossRef]
- Yang, J.; Kuang, M.T.; Yang, L.; Huang, W.; Hu, J.M. Modern interpretation of the traditional application of Shihu—A comprehensive review on phytochemistry and pharmacology progress of Dendrobium officinale. J. Ethnopharmacol. 2023, 302, 115912. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Lu, J.; Zhang, J.; Wu, J.; Yu, L.; Qin, L.; Zhu, B. Traditional Uses, Phytochemistry, Pharmacology, and Quality Control of Dendrobium officinale Kimura et. Migo. Front. Pharmacol. 2021, 12, 726528. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-H.; Wu, J.-J.; Li, X.-F.; Lu, J.-M.; Wu, W.; Sun, Y.-Q.; Zhu, B.; Qin, L.-P. Isolation, structural properties, bioactivities of polysaccharides from Dendrobium officinale Kimura et. Migo: A review. Int. J. Biol. Macromol. 2021, 184, 1000–1013. [Google Scholar] [CrossRef]
- Hou, B.; Luo, J.; Zhang, Y.; Niu, Z.; Xue, Q.; Ding, X. Iteration expansion and regional evolution: Phylogeography of Dendrobium officinale and four related taxa in southern China. Sci. Rep. 2017, 7, 43525. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.K.; Jin, J.M. China Plant Red Data Book-Rare and Endangered Plants; Science Press: Beijing, China, 1992; Volume 1. [Google Scholar]
- Si, J.P.; Yu, Q.X.; Song, X.S.; Shao, W.J. Artificial cultivation modes for Dendrobium officinale. Zhongguo Zhong Yao Za Zhi = China J. Chin. Mater. Med. 2013, 38, 481–484. [Google Scholar]
- Cheng, J.; Dang, P.-P.; Zhao, Z.; Yuan, L.-C.; Zhou, Z.-H.; Wolf, D.; Luo, Y.-B. An assessment of the Chinese medicinal Dendrobium industry: Supply, demand and sustainability. J. Ethnopharmacol. 2019, 229, 81–88. [Google Scholar] [CrossRef]
- Du, G.; Yang, R.; Yan, F.; Wei, S.; Ren, D.; Li, X. Use of Microscopic Characteristics and Multielemental Fingerprinting Analysis to Trace Three Different Cultivation Modes of Medicinal and Edible Dendrobium officinale in China. Biol. Trace Elem. Res. 2022, 201, 1006–1018. [Google Scholar] [CrossRef]
- Hornyák, M.; Dziurka, M.; Kula-Maximenko, M.; Pastuszak, J.; Szczerba, A.; Szklarczyk, M.; Płażek, A. Photosynthetic efficiency, growth and secondary metabolism of common buckwheat (Fagopyrum esculentum Moench) in different controlled-environment production systems. Sci. Rep. 2022, 12, 257. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, X.; Liao, Y.; Yang, Z. Roles of specialized metabolites in biological function and environmental adaptability of tea plant (Camellia sinensis) as a metabolite studying model. J. Adv. Res. 2021, 34, 159–171. [Google Scholar] [CrossRef]
- Strzemski, M.; Dresler, S.; Sowa, I.; Czubacka, A.; Agacka-Mołdoch, M.; Płachno, B.J.; Granica, S.; Feldo, M.; Wójciak-Kosior, M. The Impact of Different Cultivation Systems on the Content of Selected Secondary Metabolites and Antioxidant Activity of Carlina acaulis Plant Material. Molecules 2019, 25, 146. [Google Scholar] [CrossRef] [PubMed]
- Madhavi, B.G.K.; Khan, F.; Bhujel, A.; Jaihuni, M.; Kim, N.E.; Moon, B.E.; Kim, H.T. Influence of different growing media on the growth and development of strawberry plants. Heliyon 2021, 7, e07170. [Google Scholar] [CrossRef]
- Alami, M.M.; Ouyang, Z.; Zhang, Y.; Shu, S.; Yang, G.; Mei, Z.; Wang, X. The Current Developments in Medicinal Plant Genomics Enabled the Diversification of Secondary Metabolites’ Biosynthesis. Int. J. Mol. Sci. 2022, 23, 15932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Si, J.P.; Wu, L.S.; Guo, Y.Y.; Yu, J.; Wang, L.H. Field experiment of F1 generation and superior families selection of Dendrobium officinale. Zhongguo Zhong Yao Za Zhi 2013, 38, 3861–3865. [Google Scholar] [PubMed]
- Hou, Z.H.; Yin, J.L.; Lu, Y.F.; Song, J.H.; Wang, S.P.; Wei, S.D.; Liu, Z.X.; Zhang, Y.X.; Fang, Z.W. Transcriptomic analysis reveals the temporal and spatial changes in physiological process and gene expression in common buckwheat (Fagopyrum esculentum Moench) grown under drought stress. Agronomy 2019, 9, 569. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, J.; Huang, W.; Song, X.; Niu, J. Transcriptomics and Metabolomics Reveal Purine and Phenylpropanoid Metabolism Response to Drought Stress in Dendrobium sinense, an Endemic Orchid Species in Hainan Island. Front. Genet. 2021, 12, 692702. [Google Scholar] [CrossRef]
- Silveira, N.M.; Ribeiro, R.V.; de Morais, S.F.N.; de Souza, S.C.R.; da Silva, S.F.; Seabra, A.B.; Hancock, J.T.; Machado, E.C. Leaf arginine spraying improves leaf gas exchange under water deficit and root antioxidant responses during the recovery period. Plant Physiol. Biochem. PPB 2021, 162, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Savka, M.A.; Hudson, A.O. The Synthesis and Role of β-Alanine in Plants. Front. Plant Sci. 2019, 10, 921. [Google Scholar] [CrossRef]
- Iwagami, T.; Ogawa, T.; Ishikawa, T.; Maruta, T. Activation of ascorbate metabolism by nitrogen starvation and its physiological impacts in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2022, 86, 476–489. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, S.; Ma, Y.; Zhao, W.; Wang, P.; Zhao, S.; Wang, D. Ascorbic acid prevents yellowing of fresh-cut yam by regulating pigment biosynthesis and energy metabolism. Food Res. Int. 2022, 157, 111424. [Google Scholar] [CrossRef]
- Wołosiak, R.; Drużyńska, B.; Derewiaka, D.; Piecyk, M.; Majewska, E.; Ciecierska, M.; Worobiej, E.; Pakosz, P. Verification of the Conditions for Determination of Antioxidant Activity by ABTS and DPPH Assays—A Practical Approach. Molecules 2022, 27, 50. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Huebbe, P.; Konishi, T.; Rahman, M.M.; Nakahara, M.; Matsugo, S.; Rimbach, G. Free radical scavenging and antioxidant activity of ascorbigen versus ascorbic acid: Studies in vitro and in cultured human keratinocytes. J. Agric. Food Chem. 2008, 56, 11694–11699. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, N.; Russi, S.; Bruno, M.; Maresca, V.; Vaglica, A.; Ilardi, V.; Zanfardino, A.; Di Napoli, M.; Varcamonti, M.; Cianciullo, P.; et al. Dihydrophenanthrenes from a Sicilian Accession of Himantoglossum robertianum (Loisel.) P. Delforge Showed Antioxidant, Antimicrobial, and Antiproliferative Activities. Plants 2021, 10, 2776. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cao, Z.; Zhang, W.; Tickner, J.; Qiu, H.; Wang, C.; Chen, K.; Wang, Z.; Tan, R.; Dong, S.; et al. Lumichrome inhibits osteoclastogenesis and bone resorption through suppressing RANKL-induced NFAT activation and calcium signaling. J. Cell. Physiol. 2018, 233, 8971–8983. [Google Scholar] [CrossRef]
- Kauss, T.; Moynet, D.; Rambert, J.; Al-Kharrat, A.; Brajot, S.; Thiolat, D.; Ennemany, R.; Fawaz, F.; Mossalayi, M.D. Rutoside decreases human macrophage-derived inflammatory mediators and improves clinical signs in adjuvant-induced arthritis. Arthritis Res. Ther. 2008, 10, R19. [Google Scholar] [CrossRef]
- Morquette, A.J.; Lee, J.B.; Grossman, S.K.; Hsu, S. Rutoside and Ascorbic Acid in the Treatment of Schamberg Pigmented Purpuric Dermatosis. Cureus 2021, 13, e14592. [Google Scholar] [CrossRef]
- Chen, X.; Gu, N.; Xue, C.; Li, B.R. Plant flavonoid taxifolin inhibits the growth, migration and invasion of human osteosarcoma cells. Mol. Med. Rep. 2018, 17, 3239–3245. [Google Scholar] [CrossRef]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/ β -catenin signaling pathway. BMC Cancer 2018, 18, 1043. [Google Scholar] [CrossRef]
- Li, J.; Hu, L.; Zhou, T.; Gong, X.; Jiang, R.; Li, H.; Kuang, G.; Wan, J.; Li, H. Taxifolin inhibits breast cancer cells proliferation, migration and invasion by promoting mesenchymal to epithelial transition via β-catenin signaling. Life Sci. 2019, 232, 116617. [Google Scholar] [CrossRef]
- Wang, R.; Zhu, X.; Wang, Q.; Li, X.; Wang, E.; Zhao, Q.; Wang, Q.; Cao, H. The anti-tumor effect of taxifolin on lung cancer via suppressing stemness and epithelial-mesenchymal transition in vitro and oncogenesis in nude mice. Ann. Transl. Med. 2020, 8, 590. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | G vs. S | G vs. T | T vs. S | |||
|---|---|---|---|---|---|---|
| UP | DOWN | UP | DOWN | UP | DOWN | |
| Phenolic acid | 2 | 15 | 2 | 12 | 10 | 13 |
| Flavonoids | 1 | 15 | 3 | 1 | 3 | 10 |
| Lipids | 1 | 13 | 6 | 3 | 3 | 14 |
| Alkaloids | 1 | 8 | 1 | 3 | 4 | 7 |
| Saccharides and alcohols | 2 | 5 | 0 | 3 | 2 | 4 |
| Amino acids and their derivatives | 0 | 10 | 2 | 7 | 3 | 7 |
| Organic acids | 1 | 4 | 0 | 0 | 2 | 3 |
| Lignans and coumarins | 0 | 7 | 0 | 5 | 2 | 5 |
| Others | 0 | 3 | 0 | 4 | 1 | 2 |
| Nucleotides and their derivatives | 0 | 4 | 2 | 3 | 4 | 6 |
| Quinones | 0 | 5 | 2 | 0 | 0 | 6 |
| Terpenoids | 0 | 4 | 1 | 2 | 0 | 4 |
| Vitamins | 0 | 1 | 0 | 2 | 2 | 0 |
| Stilbenes | 0 | 1 | 0 | 1 | 0 | 0 |
| Total | 8 | 95 | 19 | 46 | 36 | 81 |
| Growth Method | Epiphytic Substrates | Time of Harvesting | Growing Years | Elevation (m) | Climate Type | Lightwardness | Longitude and Latitude | Number of Samples | Place of Origin |
|---|---|---|---|---|---|---|---|---|---|
| Tree epiphytic cultivation | Silktree Siris | 201906 | 3 years old | 1122 | Subtropical monsoon climate | Light | E105°27′57”N24°54′3” | 9 | Anlong |
| Stone epiphytic cultivation | Limestone | 201906 | 3 years old | 1172 | Subtropical monsoon climate | Light | E104°57′54”N24°54′18” | 9 | Xingyi |
| Greenhouse cultivation | Wood chips | 201906 | 3 years old | 1160 | - | - | E104°57′49”N24°54′17” | 9 | Xingyi |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Song, Y.; Lu, A.; Qin, L.; Tan, D.; Zhang, Q.; He, Y.; Lu, Y. Metabolomics-Based Analysis of the Effects of Different Cultivation Strategies on Metabolites of Dendrobium officinale Kimura et Migo. Metabolites 2023, 13, 389. https://doi.org/10.3390/metabo13030389
Yang D, Song Y, Lu A, Qin L, Tan D, Zhang Q, He Y, Lu Y. Metabolomics-Based Analysis of the Effects of Different Cultivation Strategies on Metabolites of Dendrobium officinale Kimura et Migo. Metabolites. 2023; 13(3):389. https://doi.org/10.3390/metabo13030389
Chicago/Turabian StyleYang, Da, Yeyang Song, Anjin Lu, Lin Qin, Daopeng Tan, Qianru Zhang, Yuqi He, and Yanliu Lu. 2023. "Metabolomics-Based Analysis of the Effects of Different Cultivation Strategies on Metabolites of Dendrobium officinale Kimura et Migo" Metabolites 13, no. 3: 389. https://doi.org/10.3390/metabo13030389
APA StyleYang, D., Song, Y., Lu, A., Qin, L., Tan, D., Zhang, Q., He, Y., & Lu, Y. (2023). Metabolomics-Based Analysis of the Effects of Different Cultivation Strategies on Metabolites of Dendrobium officinale Kimura et Migo. Metabolites, 13(3), 389. https://doi.org/10.3390/metabo13030389