

Human Metabolome Reference Database in a Biracial Cohort across the Adult Lifespan

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Health Status
2.3. Plasma Collection and Metabolomics Assessment
3. Results
3.1. Reference Values
3.2. Metabolites Associated with Age
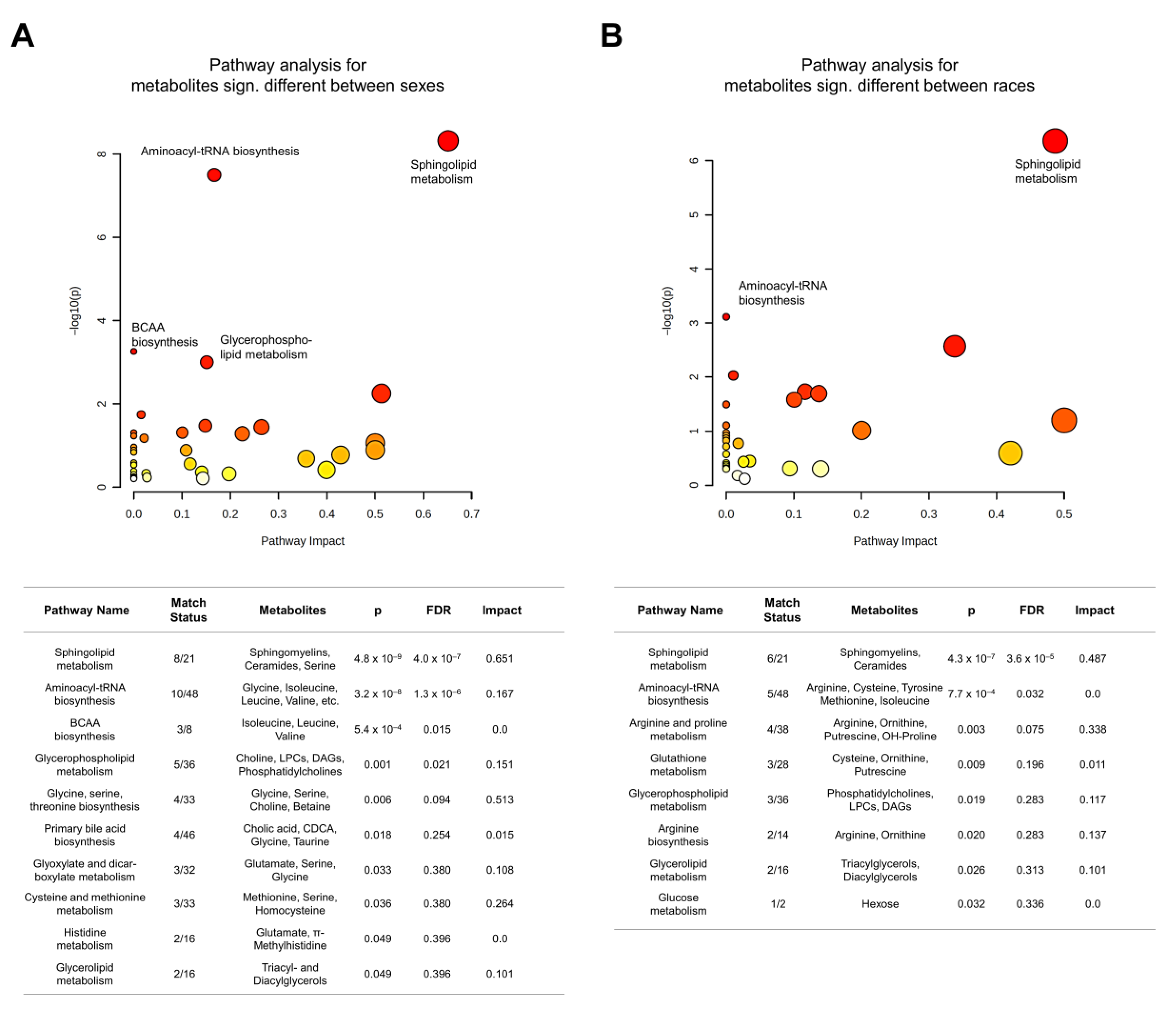
3.3. Metabolite Differences by Sex
3.4. Metabolite Differences by Race
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balashova, E.E.; Maslov, D.L.; Trifonova, O.P.; Lokhov, P.G.; Archakov, A.I. Metabolome Profiling in Aging Studies. Biology 2022, 11, 1570. [Google Scholar] [CrossRef]
- Vardarajan, B.; Kalia, V.; Manly, J.; Brickman, A.; Reyes-Dumeyer, D.; Lantigua, R.; Ionita-Laza, I.; Jones, D.P.; Miller, G.W.; Mayeux, R. Differences in plasma metabolites related to Alzheimer’s disease, APOE epsilon4 status, and ethnicity. Alzheimers Dement 2020, 6, e12025. [Google Scholar]
- Cai, X.; Perttula, K.; Sk, P.; Hubbard, A.; Dk, N.; Rappaport, S. Untargeted Lipidomic Profiling of Human Plasma Reveals Differences due to Race, Gender and Smoking Status. J. Postgenomics Drug Biomark. Dev. 2014, 4, 1–8. [Google Scholar]
- Darst, B.F.; Koscik, R.L.; Hogan, K.J.; Johnson, S.C.; Engelman, C.D. Longitudinal plasma metabolomics of aging and sex. Aging 2019, 11, 1262–1282. [Google Scholar] [CrossRef]
- Bell, J.A.; Ferreira, D.L.S.; Fraser, A.; Soares, A.L.G.; Howe, L.D.; Lawlor, D.A.; Carslake, D.; Smith, G.D.; O’keeffe, L.M. Sex differences in systemic metabolites at four life stages: Cohort study with repeated metabolomics. BMC Med. 2021, 19, 58. [Google Scholar] [CrossRef]
- Hernandes, V.V.; Dordevic, N.; Hantikainen, E.M.; Sigurdsson, B.B.; Smárason, S.V.; Garcia-Larsen, V.; Gögele, M.; Caprioli, G.; Bozzolan, I.; Pramstaller, P.P.; et al. Age, Sex, Body Mass Index, Diet and Menopause Related Metabolites in a Large Homogeneous Alpine Cohort. Metabolites 2022, 12, 205. [Google Scholar] [CrossRef]
- Trabado, S.; Al-Salameh, A.; Croixmarie, V.; Masson, P.; Corruble, E.; Fève, B.; Colle, R.; Ripoll, L.; Walther, B.; Boursier-Neyret, C.; et al. The human plasma-metabolome: Reference values in 800 French healthy volunteers; impact of cholesterol, gender and age. PLoS ONE 2017, 12, e0173615. [Google Scholar] [CrossRef]
- Yin, X.; Prendiville, O.; McNamara, A.E.; Brennan, L. Targeted Metabolomic Approach to Assess the Reproducibility of Plasma Metabolites over a Four Month Period in a Free-Living Population. J. Proteome Res. 2022, 21, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L. The Baltimore Longitudinal Study of Aging (BLSA): A 50-year-long journey and plans for the future. J. Gerontol A Biol. Sci. Med. Sci. 2008, 63, 1416–1419. [Google Scholar] [CrossRef]
- Tumasian, R.A., 3rd; Harish, A.; Kundu, G.; Yang, J.-H.; Ubaida-Mohien, C.; Gonzalez-Freire, M.; Kaileh, M.; Zukley, L.M.; Chia, C.W.; Lyashkov, A.; et al. Skeletal muscle transcriptome in healthy aging. Nat. Commun. 2021, 12, 2014. [Google Scholar] [CrossRef]
- Adam, M.G.; Limonciel, A. The Quantitative Metabolomics Database (QMDB)—A new reference database for human metabolomics. MetaboNews 2022, 12, 6–8. [Google Scholar]
- Kooperberg, C.L.; Stone, C.J. Logspline Density Estimation for Censored Data. J. Comput. Graph. Stat. 1992, 1, 301–328. [Google Scholar]
- Irizarry, R.A.; Wang, C.; Zhou, Y.; Speed, T.P. Gene set enrichment analysis made simple. Stat. Methods Med. Res. 2009, 18, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Almeida, I.; Magalhães, S.; Nunes, A. Lipids: Biomarkers of healthy aging. Biogerontology 2021, 22, 273–295. [Google Scholar] [CrossRef] [PubMed]
- Honarvar, N.M.; Zarezadeh, M.; Molsberry, S.A.; Ascherio, A. Changes in plasma phospholipids and sphingomyelins with aging in men and women: A comprehensive systematic review of longitudinal cohort studies. Ageing Res. Rev. 2021, 68, 101340. [Google Scholar] [CrossRef]
- Lawton, K.A.; Berger, A.; Mitchell, M.; Milgram, K.E.; Evans, A.M.; Guo, L.; Hanson, R.W.; Kalhan, S.C.; Ryals, J.A.; Milburn, M.V. Analysis of the adult human plasma metabolome. Pharmacogenomics 2008, 9, 383–397. [Google Scholar] [CrossRef]
- Menni, C.; Kastenmüller, G.; Petersen, A.K.; Bell, J.T.; Psatha, M.; Tsai, P.-C.; Gieger, C.; Schulz, H.; Erte, I.; John, S.; et al. Metabolomic markers reveal novel pathways of ageing and early development in human populations. Int. J. Epidemiol. 2013, 42, 1111–1119. [Google Scholar] [CrossRef]
- Slade, E.; Irvin, M.R.; Xie, K.; Arnett, D.K.; Claas, S.A.; Kind, T.; Fardo, D.W.; Graf, G.A. Age and sex are associated with the plasma lipidome: Findings from the GOLDN study. Lipids Health. Dis. 2021, 20, 30. [Google Scholar] [CrossRef]
- Di Cesare, F.; Luchinat, C.; Tenori, L.; Saccenti, E. Age- and Sex-Dependent Changes of Free Circulating Blood Metabolite and Lipid Abundances, Correlations, and Ratios. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 77, 918–926. [Google Scholar] [CrossRef]
- Mielke, M.M.; Bandaru, V.V.R.; Han, D.; An, Y.; Resnick, S.M.; Ferrucci, L.; Haughey, N.J. Factors affecting longitudinal trajectories of plasma sphingomyelins: The Baltimore Longitudinal Study of Aging. Aging Cell 2014, 14, 112–121. [Google Scholar] [CrossRef]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef]
- Schwedhelm, E.; Xanthakis, V.; Maas, R.; Sullivan, L.M.; Atzler, D.; Lüneburg, N.; Glazer, N.L.; Riederer, U.; Vasan, R.S.; Böger, R.H. Plasma symmetric dimethylarginine reference limits from the Framingham offspring cohort. Clin. Chem. Lab. Med. 2011, 49, 1907–1910. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brattström, L.; Lindgren, A.; Israelsson, B.; Andersson, A.; Hultberg, B. Homocysteine and cysteine: Determinants of plasma levels in middle-aged and elderly subjects. J. Intern. Med. 1994, 236, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Toroser, D.; Sohal, R.S. Age-associated perturbations in glutathione synthesis in mouse liver. Biochem. J. 2007, 405, 583–589. [Google Scholar] [CrossRef]
- Sorgdrager, F.J.H.; Vermeiren, Y.; Van Faassen, M.; van der Ley, C.; Nollen, E.A.A.; Kema, I.P.; De Deyn, P.P. Age- and disease-specific changes of the kynurenine pathway in Parkinson’s and Alzheimer’s disease. J. Neurochem. 2019, 151, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Ueland, P.M. Choline and betaine in health and disease. J. Inherit. Metab. Dis. 2011, 34, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Davy, K.P.; Seals, D.R. Total blood volume in healthy young and older men. J. Appl. Physiol. (1985) 1994, 76, 2059–2062. [Google Scholar] [CrossRef]
- Samaras, N.; Samaras, D.; Frangos, E.; Forster, A.; Philippe, J. A review of age-related dehydroepiandrosterone decline and its association with well-known geriatric syndromes: Is treatment beneficial? Rejuvenation Res. 2013, 16, 285–294. [Google Scholar] [CrossRef]
- Baxmann, A.C.A.A.; Ahmed, M.S.; Marques, N.A.A.C.; Menon, V.B.; Pereira, A.B.; Kirsztajn, G.M.; Heilberg, I.P. Influence of muscle mass and physical activity on serum and urinary creatinine and serum cystatin C. Clin. J. Am. Soc. Nephrol. 2008, 3, 348–354. [Google Scholar] [CrossRef]
- Sayda, M.H.; Phillips, B.E.; Williams, J.P.; Greenhaff, P.L.; Wilkinson, D.J.; Smith, K.; Atherton, P.J. Associations between Plasma Branched Chain Amino Acids and Health Biomarkers in Response to Resistance Exercise Training Across Age. Nutrients 2020, 12, 3029. [Google Scholar] [CrossRef]
- Larsson, C.A.; Gullberg, B.; Råstam, L.; Lindblad, U. Salivary cortisol differs with age and sex and shows inverse associations with WHR in Swedish women: A cross-sectional study. BMC Endocr. Disord. 2009, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Krumsiek, J.; Mittelstrass, K.; Do, K.T.; Stückler, F.; Ried, J.; Adamski, J.; Peters, A.; Illig, T.; Kronenberg, F.; Friedrich, N.; et al. Gender-specific pathway differences in the human serum metabolome. Metabolomics 2015, 11, 1815–1833. [Google Scholar] [CrossRef]
- Hu, J.; Yao, J.; Dseng, S.; Balasubramanian, R.; Jiménez, M.C.; Li, J.; Guo, X.; Cruz, D.E.; Gao, Y.; Huang, T.; et al. Differences in Metabolomic Profiles Between Black and White Women and Risk of Coronary Heart Disease: An Observational Study of Women From Four US Cohorts. Circ. Res. 2022, 131, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Wagner-Golbs, A.; Neuber, S.; Kamlage, B.; Christiansen, N.; Bethan, B.; Rennefahrt, U.; Schatz, P.; Lind, L. Effects of Long-Term Storage at −80 °C on the Human Plasma Metabolome. Metabolites 2019, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Haid, M.; Muschet, C.; Wahl, S.; Römisch-Margl, W.; Prehn, C.; Möller, G.; Adamski, J. Long-Term Stability of Human Plasma Metabolites during Storage at −80 °C. J. Proteome Res. 2018, 17, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Reis, G.B.; Rees, J.C.; Ivanova, A.A.; Kuklenyik, Z.; Drew, N.M.; Pirkle, J.L.; Barr, J.R. Stability of lipids in plasma and serum: Effects of temperature-related storage conditions on the human lipidome. J. Mass Spectrom. Adv. Clin. Lab 2021, 22, 34–42. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | 20–30 | 30–40 | 40–50 | 50–60 | 60–70 | 70–80 | 80–90 | 90+ |
|---|---|---|---|---|---|---|---|---|
| Sample size, n | ||||||||
| White Men (n = 204) | 10 | 29 | 24 | 30 | 30 | 30 | 30 | 21 |
| White Women (n = 213) | 13 | 30 | 28 | 30 | 30 | 30 | 30 | 22 |
| Black Men (n = 122) | 4 | 7 | 14 | 30 | 30 | 23 | 14 | 1 |
| Black Women (n = 148) | 3 | 5 | 22 | 30 | 30 | 30 | 28 | 3 |
| Age, Mean ± SD (range) | ||||||||
| White Men | 25.2 ± 2.5 (22–29) | 34.8 ± 3.1 (30–39) | 46.9 ± 2.1 (41–49) | 55.7 ± 2.9 (50–59) | 65.5 ± 2.4 (60–69) | 74.0 ± 2.7 (70–79) | 83.7 ± 2.5 (80–89) | 91.4 ± 1.4 (90–94) |
| White Women | 27.0 ± 1.6 (24–29) | 36.1 ± 2.7 (30–39) | 45.5 ± 2.4 (40–49) | 56.2 ± 3.0 (50–59) | 65.6 ± 2.6 (60–69) | 73.4 ± 2.3 (70–78) | 83.4 ± 2.3 (80–88) | 90.8 ± 1.3 (90–94) |
| Black Men | 26.5 ± 1.0 (26–28) | 33.4 ± 3.4 (31–39) | 45.7 ± 2.3 (40–49) | 54.5 ± 2.6 (50–59) | 64.4±2.7 (61–69) | 73.2 ± 2.5 (70–78) | 81.8 ± 2.0 (80–87) | 91 – |
| Black Women | 28 ± 0 28 | 35.8 ± 1.6 (34–38) | 46.1 ± 2.5 (40–49) | 56.3 ± 2.8 (51–59) | 65.3 ± 3.2 (60–69) | 73.1 ± 2.5 (70–79) | 83.0 ± 2.0 (80–87) | 91.7 ± 2.1 (90–94) |
| Body mass index, kg/m2, Mean ± SD | ||||||||
| White Men | 25.0 ± 2.4 | 25.9 ± 3.1 | 27.4 ± 3.7 | 27.1 ± 3.3 | 28.7 ± 3.3 | 26.6 ± 2.5 | 25.8 ± 3.1 | 25.8 ± 3.6 |
| White Women | 25.4 ± 3.0 | 24.1 ± 3.8 | 24.6 ± 3.7 | 24.9 ± 3.7 | 26.1 ± 4.8 | 25.6 ± 3.6 | 24.8 ± 3.3 | 23.8 ± 3.7 |
| Black Men | 25.1 ± 3.6 | 24.7 ± 1.7 | 28.0 ± 3.8 | 29.0 ± 3.9 | 28.4 ± 3.6 | 28.3 ± 3.8 | 27.0 ± 3.9 | 24.7 |
| Black Women | 23.6 ± 2.9 | 28.6 ± 3.4 | 26.1 ± 3.8 | 26.7 ± 3.4 | 29.1 ± 3.7 | 28.6 ± 3.7 | 26.5 ± 3.6 | 26.7 ± 3.3 |
| Gait speed, m/s, Mean ± SD | ||||||||
| White Men | 1.33 ± 0.15 | 1.37 ± 0.23 | 1.38 ± 0.16 | 1.35 ± 0.19 | 1.31 ± 0.19 | 1.26 ± 0.17 | 1.17 ± 0.15 | 0.95 ± 0.14 |
| White Women | 1.35 ± 0.15 | 1.36 ± 0.19 | 1.39 ± 0.17 | 1.33 ± 0.18 | 1.31 ± 0.15 | 1.17 ± 0.16 | 1.14 ± 0.14 | 0.92 ± 0.17 |
| Black Men | 1.32 ± 0.36 | 1.20 ± 0.14 | 1.30 ± 0.30 | 1.20 ± 0.18 | 1.16 ± 0.15 | 1.18 ± 0.25 | 1.08 ± 0.18 | 1.12 |
| Black Women | 1.24 ± 0.08 | 1.42 ± 0.30 | 1.19 ± 0.19 | 1.17 ± 0.15 | 1.09 ± 0.11 | 1.04 ± 0.18 | 0.96 ± 0.13 | 0.80 ± 0.27 |
| White Men | White Women | Black Men | Black Women | |
|---|---|---|---|---|
| Biochemical Classes | p-Value | |||
| Acylcarnitines | 0.057 | 0.007 | 0.004 | 3.9 × 10−6 |
| Amino-acid-related | 0.001 | 0.001 | 3.2 × 10−5 | 1.2 × 10−9 |
| Amino acids | 0.115 | 0.332 | 0.298 | 0.307 |
| Bile acids | 0.985 | 0.991 | 0.197 | 0.983 |
| Biogenic amines | 0.284 | 0.421 | 0.121 | 0.106 |
| Carboxylic acids | 0.041 | 0.132 | 0.042 | 0.022 |
| Ceramides | 3.8 × 10−4 | 2.0 × 10−8 | 0.009 | 2.8 × 10−6 |
| Cholesteryl esters | 0.812 | 0.678 | 0.002 | 0.914 |
| Diglycerides | 0.612 | 0.999 | 0.676 | 0.763 |
| Dihexosylceramides | 3.4 × 10−4 | 0.004 | 0.138 | 0.269 |
| Dihydroceramides | 0.181 | 0.047 | 0.635 | 0.422 |
| Fatty acids | 0.163 | 0.971 | 0.518 | 0.508 |
| Hexosylceramides | 1.7 × 10−7 | 0.003 | 0.886 | 0.393 |
| Hormones and related | 0.119 | 0.228 | 0.097 | 0.063 |
| Indoles and derivatives | 0.541 | 0.859 | 0.439 | 0.083 |
| Lysophosphatidylcholines | 0.897 | 0.990 | 0.896 | 0.591 |
| Phosphatidylcholines | 3.3 × 10−7 | 0.859 | 0.001 | 0.865 |
| Sphingomyelins | 0.020 | 2.7 × 10−5 | 0.897 | 0.014 |
| Triglycerides | 0.999 | 0.999 | 0.999 | 0.999 |
| Trihexosylceramides | 0.004 | 0.120 | 0.013 | 0.014 |
| Classes Associated with Sex | p-Value | Classes Associated with Race | p-Value |
|---|---|---|---|
| Phosphatidylcholines | 2.2 × 10−11 | Lysophosphatidylcholines | 0.045 |
| Sphingomyelins | 2.9 × 10−8 | Phosphatidylcholines | 0.046 |
| Ceramides | 4.2 × 10−4 | Triglycerides | 0.047 |
| Dihexosylceramides | 0.047 | Cholesteryl esters | 0.204 |
| Trihexosylceramides | 0.051 | Dihexosylceramides | 0.275 |
| Amino acids | 0.064 | Diglycerides | 0.315 |
| Amino-acid-related | 0.077 | Amino-acid-related | 0.326 |
| Hexosylceramides | 0.079 | Sphingomyelins | 0.393 |
| Cholesteryl esters | 0.081 | Bile acids | 0.439 |
| Hormones and related | 0.119 | Dihydroceramides | 0.454 |
| Diglycerides | 0.305 | Indoles and derivatives | 0.586 |
| Biogenic amines | 0.417 | Hormones and related | 0.742 |
| Acylcarnitines | 0.590 | Biogenic amines | 0.749 |
| Dihydroceramides | 0.725 | Amino acids | 0.813 |
| Lysophosphatidylchlines | 0.862 | Fatty acids | 0.829 |
| Bile acids | 0.885 | Trihexosylceramides | 0.882 |
| Indoles and derivatives | 0.944 | Acylcarnitines | 0.908 |
| Fatty acids | 0.972 | Carboxylic acids | 0.970 |
| Carboxylic acids | 0.997 | Hexosylceramides | 0.998 |
| Triglycerides | 0.999 | Ceramides | 0.999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Q.; Adam, M.G.; Ozcariz, E.; Fantoni, G.; Shehadeh, N.M.; Turek, L.M.; Collingham, V.L.; Kaileh, M.; Moaddel, R.; Ferrucci, L. Human Metabolome Reference Database in a Biracial Cohort across the Adult Lifespan. Metabolites 2023, 13, 591. https://doi.org/10.3390/metabo13050591
Tian Q, Adam MG, Ozcariz E, Fantoni G, Shehadeh NM, Turek LM, Collingham VL, Kaileh M, Moaddel R, Ferrucci L. Human Metabolome Reference Database in a Biracial Cohort across the Adult Lifespan. Metabolites. 2023; 13(5):591. https://doi.org/10.3390/metabo13050591
Chicago/Turabian StyleTian, Qu, M. Gordian Adam, Enrique Ozcariz, Giovanna Fantoni, Nader M. Shehadeh, Lisa M. Turek, Victoria L. Collingham, Mary Kaileh, Ruin Moaddel, and Luigi Ferrucci. 2023. "Human Metabolome Reference Database in a Biracial Cohort across the Adult Lifespan" Metabolites 13, no. 5: 591. https://doi.org/10.3390/metabo13050591
APA StyleTian, Q., Adam, M. G., Ozcariz, E., Fantoni, G., Shehadeh, N. M., Turek, L. M., Collingham, V. L., Kaileh, M., Moaddel, R., & Ferrucci, L. (2023). Human Metabolome Reference Database in a Biracial Cohort across the Adult Lifespan. Metabolites, 13(5), 591. https://doi.org/10.3390/metabo13050591