
Maternal Magnolol Supplementation during Pregnancy and Lactation Promotes Antioxidant Capacity, Improves Gut Health, and Alters Gut Microbiota and Metabolites of Weanling Piglets

 ,
,
Abstract
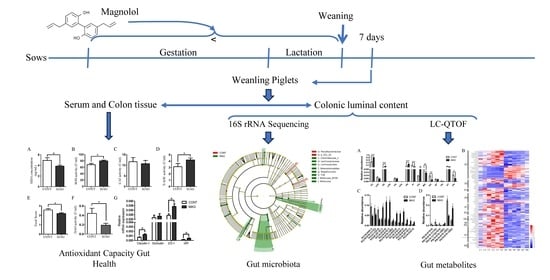
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Data Recording
2.3. Blood and Tissue Sample
2.4. H&E Staining
2.5. Measurement of Serum Oxidative Status
2.6. Measurement of Serum Diamine Oxidase (DAO)
2.7. Quantitative Real-Time PCR
2.8. The 16S rRNA High-Throughput Sequencing
2.9. LC-QTOF Analysis of Piglet Colonic Content Metabolites
2.10. Statistical Analyses
3. Results
3.1. Gestation and Lactation Performance of Sows
3.2. Reproductive and Litter Performance of Sows
3.3. Organ Weight, Length, and Morphology of Intestine of Weanling Piglets
3.4. Serum Antioxidant Capacity, Diarrhea, and Intestinal Barrier Function of Weanling Piglets
3.5. Diversity of Colon Microbiota in Weanling Piglets
3.6. Taxonomic Differences of Colon Microbiota in Weanling Piglets
3.7. Predictive Analysis of Microbiota Function in Weanling Piglets
3.8. Colonic Luminal Metabolomic Analysis of Weanling Piglets
3.9. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Yang, S.; Zhang, X.; Liu, X.; Wu, Z.; Qi, Y.; Guan, W.; Ren, M.; Zhang, S. Maternal Nutrition During Late Gestation and Lactation: Association With Immunity and the Inflammatory Response in the Offspring. Front. Immunol. 2021, 12, 758525. [Google Scholar] [CrossRef]
- Feyera, T.; Hojgaard, C.K.; Vinther, J.; Bruun, T.S.; Theil, P.K. Dietary supplement rich in fiber fed to late gestating sows during transition reduces rate of stillborn piglets. J. Anim. Sci. 2017, 95, 5430–5438. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Chen, X.; Liu, Z.; Diao, X. Effects of soybean hulls and corn stalk on the performance, colostrum composition and faecal microflora of pregnant sows. J. Anim. Physiol. Anim. Nutr. 2022, 107, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhang, W.; Zhao, H.; Tian, Z.; Ren, M.; Chen, F.; Guan, W.; Zhang, S. Dietary Supplementation of Enzymatically Treated Artemisia annua L. Improves Lactation Performance, Alleviates Inflammatory Response of Sows Reared Under Heat Stress, and Promotes Gut Development in Preweaning Offspring. Front. Vet. Sci. 2022, 9, 843673. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Everard, A.; Duparc, T. Gut microbiota, enteroendocrine functions and metabolism. Curr. Opin. Pharmacol. 2013, 13, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wei, H.; Ao, J.; Long, G.; Peng, J. Inclusion of Konjac Flour in the Gestation Diet Changes the Gut Microbiota, Alleviates Oxidative Stress, and Improves Insulin Sensitivity in Sows. Appl. Environ. Microbiol. 2016, 82, 5899–5909. [Google Scholar] [CrossRef]
- Bi, Y.; Tu, Y.; Zhang, N.; Wang, S.; Zhang, F.; Suen, G.; Shao, D.; Li, S.; Diao, Q. Multiomics analysis reveals the presence of a microbiome in the gut of fetal lambs. Gut 2021, 70, 853–864. [Google Scholar] [CrossRef]
- Grzeskowiak, L.; Saliu, E.M.; Martinez-Vallespin, B.; Wessels, A.G.; Manner, K.; Vahjen, W.; Zentek, J. Fiber Composition in Sows’ Diets Modifies Clostridioides difficile Colonization in Their Offspring. Curr. Microbiol. 2022, 79, 154. [Google Scholar] [CrossRef]
- Ma, C.; Azad, M.A.K.; Tang, W.; Zhu, Q.; Wang, W.; Gao, Q.; Kong, X. Maternal probiotics supplementation improves immune and antioxidant function in suckling piglets via modifying gut microbiota. J. Appl. Microbiol. 2022, 133, 515–528. [Google Scholar] [CrossRef]
- Dabulici, C.M.; Sarbu, I.; Vamanu, E. The Bioactive Potential of Functional Products and Bioavailability of Phenolic Compounds. Foods 2020, 9, 953. [Google Scholar] [CrossRef]
- Patra, A.K.; Amasheh, S.; Aschenbach, J.R. Modulation of gastrointestinal barrier and nutrient transport function in farm animals by natural plant bioactive compounds—A comprehensive review. Crit. Rev. Food. Sci. Nutr. 2019, 59, 3237–3266. [Google Scholar] [CrossRef]
- Shen, C.C.; Ni, C.L.; Shen, Y.C.; Huang, Y.L.; Kuo, C.H.; Wu, T.S.; Chen, C.C. Phenolic constituents from the stem bark of Magnolia officinalis. J. Nat. Prod. 2009, 72, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Z.; Huang, X.; Shi, W.; Zhang, R.; Chen, M.; Huang, H.; Wu, L. Insights on the Multifunctional Activities of Magnolol. Biomed. Res. Int. 2019, 2019, 1847130. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Jiang, Z.; Yang, B.; Hu, Z. Magnolol exhibits anti-inflammatory and neuroprotective effects in a rat model of intracerebral haemorrhage. Brain Behav. Immun. 2019, 77, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Xie, K.; Yi, J.; Song, Z.; Zhang, H.; He, X. The effects of magnolol supplementation on growth performance, meat quality, oxidative capacity, and intestinal microbiota in broilers. Poult. Sci. 2022, 101, 101722. [Google Scholar] [CrossRef]
- Peng, W.; Zhou, N.; Song, Z.; Zhang, H.; He, X. Magnolol as a Protective Antioxidant Alleviates Rotenone-Induced Oxidative Stress and Liver Damage through MAPK/mTOR/Nrf2 in Broilers. Metabolites 2023, 13, 84. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, H.; Du, E.; Jin, F.; Zheng, C.; Fan, Q.; Zhao, N.; Guo, W.; Zhang, W.; Huang, S.; et al. Effects of magnolol on egg production, egg quality, antioxidant capacity, and intestinal health of laying hens in the late phase of the laying cycle. Poult. Sci. 2021, 100, 835–843. [Google Scholar] [CrossRef]
- Fan, Q.; Chen, F.; Zhang, W.; Du, E.; Zhao, N.; Huang, S.; Guo, W.; Yan, X.; Chen, M.; Wei, J. Maternal magnolol supplementation alters placental morphology, promotes placental angiogenesis during mid-gestation and improves offspring growth in a pregnant mouse model. Reprod. Biol. 2021, 21, 100567. [Google Scholar] [CrossRef]
- Xu, B.; Qin, W.; Xu, Y.; Yang, W.; Chen, Y.; Huang, J.; Zhao, J.; Ma, L. Dietary Quercetin Supplementation Attenuates Diarrhea and Intestinal Damage by Regulating Gut Microbiota in Weanling Piglets. Oxid. Med. Cell. Longev. 2021, 2021, 6221012. [Google Scholar] [CrossRef]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.R.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 133462. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Kon, R.; Tsubota, Y.; Minami, M.; Kato, S.; Matsunaga, Y.; Kimura, H.; Murakami, Y.; Fujikawa, T.; Sakurai, R.; Tomimoto, R.; et al. CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon. Int. J. Mol. Sci. 2018, 19, 170. [Google Scholar] [CrossRef]
- Pop, M.; Walker, A.W.; Paulson, J.; Lindsay, B.; Antonio, M.; Hossain, M.A.; Oundo, J.; Tamboura, B.; Mai, V.; Astrovskaya, I.; et al. Diarrhea in young children from low-income countries leads to large-scale alterations in intestinal microbiota composition. Genome Biol. 2014, 15, R76. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, H.D.; Catalano, P.; Zhang, C.; Desoye, G.; Mathiesen, E.R.; Damm, P. Gestational diabetes mellitus. Nat. Rev. Dis. Primers 2019, 5, 47. [Google Scholar] [CrossRef]
- Szalabska-Rapala, K.; Borymska, W.; Kaczmarczyk-Sedlak, I. Effectiveness of Magnolol, a Lignan from Magnolia Bark, in Diabetes, Its Complications and Comorbidities—A Review. Int. J. Mol. Sci. 2021, 22, 10050. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhao, R.; Liang, J.C.; Chen, Y. The antidiabetic and hepatoprotective effects of magnolol on diabetic rats induced by high-fat diet and streptozotocin. Yao Xue Xue Bao 2014, 49, 476–481. [Google Scholar]
- Wang, J.; Wang, H. Oxidative Stress in Pancreatic Beta Cell Regeneration. Oxid. Med. Cell. Longev. 2017, 2017, 1930261. [Google Scholar] [CrossRef]
- Robertson, R.P.; Harmon, J.S. Diabetes, glucose toxicity, and oxidative stress: A case of double jeopardy for the pancreatic islet beta cell. Free Radic. Biol. Med. 2006, 41, 177–184. [Google Scholar] [CrossRef]
- Suh, K.S.; Chon, S.; Jung, W.W.; Choi, E.M. Magnolol protects pancreatic β-cells against methylglyoxal-induced cellular dysfunction. Chem. Biol. Interact. 2017, 277, 101–109. [Google Scholar] [CrossRef]
- Kim, Y.S.; Dong, H.J.; Nan, H.K.; Yun, M.L.; Jin, S.K. Effect of magnolol on TGF-β1 and fibronectin expression in human retinal pigment epithelial cells under diabetic conditions. Eur. J. Pharmacol. 2007, 562, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Parray, H.A.; Lone, J.; Park, J.P.; Choi, J.W.; Yun, J.W. Magnolol promotes thermogenesis and attenuates oxidative stress in 3T3-L1 adipocytes. Nutrition 2018, 50, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Pearodwong, P.; Tretipskul, C.; Soede, N.M.; Tummaruk, P. Factors affecting estrus and ovulation time in weaned sows with induced ovulation by GnRH administration in different seasons. J. Vet. Med. Sci. 2019, 81, 1567–1574. [Google Scholar] [CrossRef] [PubMed]
- Cicalau, G.I.P.; Babes, P.A.; Calniceanu, H.; Popa, A.; Ciavoi, G.; Iova, G.M.; Ganea, M.; Scrobota, I. Anti-Inflammatory and Antioxidant Properties of Carvacrol and Magnolol, in Periodontal Disease and Diabetes Mellitus. Molecules 2021, 26, 6899. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, Y.; Liu, M.; Huang, Y.K.; Zhang, J.; Yao, Q.; Zhao, Y.L.; Xiong, J.J. Evaluating Intestinal Permeability by Measuring Plasma Endotoxin and Diamine Oxidase in Children with Acute Lymphoblastic Leukemia Treated with High-dose Methotrexate. Anticancer. Agents Med. Chem. 2016, 16, 387–392. [Google Scholar] [CrossRef]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight Junction Proteins and Signaling Pathways in Cancer and Inflammation: A Functional Crosstalk. Front. Physiol. 2018, 9, 1942. [Google Scholar] [CrossRef]
- Liu, W.; Hu, D.; Huo, H.; Zhang, W.; Adiliaghdam, F.; Morrison, S.; Ramirez, J.M.; Gul, S.S.; Hamarneh, S.R.; Hodin, R.A. Intestinal Alkaline Phosphatase Regulates Tight Junction Protein Levels. J. Am. Coll. Surg. 2016, 222, 1009–1017. [Google Scholar] [CrossRef]
- Alvarenga, L.; Cardozo, L.; Lindholm, B.; Stenvinkel, P.; Mafra, D. Intestinal alkaline phosphatase modulation by food components: Predictive, preventive, and personalized strategies for novel treatment options in chronic kidney disease. EPMA J. 2020, 11, 565–579. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Carlsson, A.H.; Yakymenko, O.; Olivier, I.; Hakansson, F.; Postma, E.; Keita, A.V.; Soderholm, J.D. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice DSS colitis. Scand. J. Gastroenterol. 2013, 48, 1136–1144. [Google Scholar] [CrossRef]
- Lenoir, M.; Martin, R.; Torres-Maravilla, E.; Chadi, S.; Gonzalez-Davila, P.; Sokol, H.; Langella, P.; Chain, F.; Bermudez-Humaran, L.G. Butyrate mediates anti-inflammatory effects of Faecalibacterium prausnitzii in intestinal epithelial cells through Dact3. Gut Microbes 2020, 12, 1–16. [Google Scholar] [CrossRef]
- Duncan, S.H.; Barcenilla, A.; Stewart, C.S.; Pryde, S.E.; Flint, H.J. Acetate utilization and butyryl coenzyme A (CoA):acetate-CoA transferase in butyrate-producing bacteria from the human large intestine. Appl. Environ. Microbiol. 2002, 68, 5186–5190. [Google Scholar] [CrossRef]
- Keren, N.; Konikoff, F.M.; Paitan, Y.; Gabay, G.; Reshef, L.; Naftali, T.; Gophna, U. Interactions between the intestinal microbiota and bile acids in gallstones patients. Environ. Microbiol. Rep. 2015, 7, 874–880. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, X.; Wang, J.; Liao, S.; Duan, Y.; Li, H.; Song, Z.; He, X.; Fan, Z. Effects of Dietary Isomaltooligosaccharide Levels on the Gut Microbiota, Immune Function of Sows, and the Diarrhea Rate of Their Offspring. Front. Microbiol. 2020, 11, 588986. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, S.; Li, S.; Zhao, L.; Hao, Y.; Qin, J.; Zhang, L.; Zhang, C.; Bian, W.; Zuo, L.; et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut 2020, 69, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Stachulski, A.V.; Knausenberger, T.B.; Shah, S.N.; Hoyles, L.; McArthur, S. A host-gut microbial amino acid co-metabolite, p-cresol glucuronide, promotes blood-brain barrier integrity in vivo. Tissue Barriers 2023, 11, 2073175. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhang, K.; Zhang, K.; Wang, G.; Wang, L.; Zhang, J.; Qiu, Z.; Guo, Z.; Song, X.; Li, J. Integrated 16S rDNA Gene Sequencing and Untargeted Metabolomics Analyses to Investigate the Gut Microbial Composition and Plasma Metabolic Phenotype in Calves with Dampness-Heat Diarrhea. Front. Vet. Sci. 2022, 9, 703051. [Google Scholar] [CrossRef]
- Callens, N.; Brugger, B.; Bonnafous, P.; Drobecq, H.; Gerl, M.J.; Krey, T.; Roman-Sosa, G.; Rumenapf, T.; Lambert, O.; Dubuisson, J.; et al. Morphology and Molecular Composition of Purified Bovine Viral Diarrhea Virus Envelope. PLoS Pathog. 2016, 12, e1005476. [Google Scholar] [CrossRef]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell. Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Y.; Song, X.; Jia, Q.; Zhang, X.; Qian, Y.; Qiu, J. Analysis of glycerophospholipid metabolism after exposure to PCB153 in PC12 cells through targeted lipidomics by UHPLC-MS/MS. Ecotoxicol. Environ. Saf. 2019, 169, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Olson, A.; Diebel, L.N.; Liberati, D.M. Exogenous phosphatidylcholine supplementation improves intestinal barrier defense against Clostridium difficile toxin. J. Trauma Acute Care Surg. 2014, 77, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Carlin, S.; Kennelly, J.P.; Fedoruk, H.; Quiroga, A.; Leonard, K.A.; Nelson, R.; Thiesen, A.; Buteau, J.; Lehner, R.; Jacobs, R. De novo phosphatidylcholine synthesis in the small intestinal epithelium is required for normal dietary lipid handling and maintenance of the mucosal barrier. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2022, 1867, 159109. [Google Scholar] [CrossRef]
- Holthuis, J.C.; Menon, A.K. Lipid landscapes and pipelines in membrane homeostasis. Nature 2014, 510, 48–57. [Google Scholar] [CrossRef]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063.e8. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Y.; Ma, F.; Sun, M.; Song, Y.; Xu, D.; Mu, G.; Tuo, Y. Lactobacillus plantarum Y44 alleviates oxidative stress by regulating gut microbiota and colonic barrier function in Balb/C mice with subcutaneous D-galactose injection. Food Funct. 2021, 12, 373–386. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment | p-Value | |
|---|---|---|---|
| CONT | MAG | ||
| No. of sows | 25 | 25 | - |
| Sow BFT, mm | |||
| Breeding (day 0) | 9.51 ± 2.27 | 9.2 ± 1.51 | 0.581 |
| Gestation (day 110) | 19.85 ± 3.26 | 21.14 ± 2.39 | 0.083 |
| Weaning | 16.61 ± 2.43 | 18.62 ± 2.19 * | 0.0016 |
| Loss lactation | 3.57 ± 1.83 | 2.66 ± 1.63 | 0.068 |
| Feed intake, kg | |||
| Gestation ADFI (day 110) | 2.86 ± 0.38 | 2.84 ± 0.42 | 0.921 |
| Lactation ADFI (day 20) | 6.05 ± 0.61 | 6.21 ± 0.57 | 0.847 |
| WOI, days | 5.50 ± 0.55 | 4.75 ± 0.71 | 0.074 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| CONT | MAG | ||
| Sow reproductive performance, n | 22 | 21 | - |
| Birth weight, kg | 1.25 ± 0.43 | 1.28 ± 0.39 | 0.716 |
| Total born piglets, n | 9.23 ± 2.83 | 9.81 ± 3.06 | 0.219 |
| Piglets born alive, n | 9.00 ± 2.81 | 9.43 ± 3.22 | 0.534 |
| Born alive piglet weight, kg | 1.36 ± 0.33 | 1.31 ± 0.36 | 0.287 |
| Litter performance, n | 15 | 15 | - |
| Litter size at cross-fostering (CF), n | 12.1 ± 1.33 | 11.9 ± 0.53 | 0.919 |
| Piglet CF weight, kg | 1.41 ± 0.54 | 1.39 ± 0.32 | 0.663 |
| Piglet lactation weight day 21, kg | 6.03 ± 1.33 | 6.09 ± 1.30 | 0.812 |
| Piglet postweaning weight day 7, kg | 7.49 ± 1.16 | 7.25 ± 1.33 | 0.711 |
| Preweaning mortality rate, % | 2.09 ± 0.08 | 1.57 ± 0.06 * | 0.047 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| CONT | MAG | ||
| No. of piglets | 6 | 6 | - |
| Body weight, kg | 7.79 ± 0.26 | 7.85 ± 0.25 | 0.740 |
| Heart, g | 50.0 ± 7.07 | 47.5 ± 7.58 | 0.624 |
| Liver, g | 166.67 ± 18.07 | 174.32 ± 8.84 | 0.263 |
| Spleen, g | 18.33 ± 6.06 | 17.5 ± 4.18 | 0.792 |
| Lung, g | 120.83 ± 25.77 | 142.5 ± 27.88 | 0.109 |
| Kidney, g | 39.17 ± 5.85 | 43.33 ± 5.16 | 0.341 |
| Duodenum, cm | 27.25 ± 2.75 | 33.75 ± 4.95 * | 0.050 |
| Jejunum + Ileum, cm | 723.5 ± 101.02 | 856 ± 60.42 * | 0.041 |
| Colon, cm | 94.83 ± 12.86 | 90.17 ± 9.39 | 0.740 |
| Day Postweaning | Number of Diarrhea Piglets/Total Piglets | χ2 | p-Value | |
|---|---|---|---|---|
| CONT | MAG | |||
| Day 1 | 6/140 | 2/111 | 0.514 | 0.473 |
| Day 2 | 5/140 | 3/111 | 0.000 | 0.988 |
| Day 3 | 8/140 | 1/111 | 2.652 | 0.103 |
| Day 4 | 9/127 | 1/106 | 3.567 | 0.059 |
| Day 5 | 7/127 | 1/106 | 2.205 | 0.138 |
| Day 6 | 8/119 | 0/104 * | 5.033 | 0.025 |
| Day 7 | 9/119 | 1/104 | 3.815 | 0.051 |
| Total in 1 week | 52/912 | 9/753 * | 22.158 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Q.; Du, E.; Chen, F.; Tao, W.; Zhao, N.; Huang, S.; Guo, W.; Huang, J.; Wei, J. Maternal Magnolol Supplementation during Pregnancy and Lactation Promotes Antioxidant Capacity, Improves Gut Health, and Alters Gut Microbiota and Metabolites of Weanling Piglets. Metabolites 2023, 13, 797. https://doi.org/10.3390/metabo13070797
Fan Q, Du E, Chen F, Tao W, Zhao N, Huang S, Guo W, Huang J, Wei J. Maternal Magnolol Supplementation during Pregnancy and Lactation Promotes Antioxidant Capacity, Improves Gut Health, and Alters Gut Microbiota and Metabolites of Weanling Piglets. Metabolites. 2023; 13(7):797. https://doi.org/10.3390/metabo13070797
Chicago/Turabian StyleFan, Qiwen, Encun Du, Fang Chen, Wenjing Tao, Na Zhao, Shaowen Huang, Wanzheng Guo, Jing Huang, and Jintao Wei. 2023. "Maternal Magnolol Supplementation during Pregnancy and Lactation Promotes Antioxidant Capacity, Improves Gut Health, and Alters Gut Microbiota and Metabolites of Weanling Piglets" Metabolites 13, no. 7: 797. https://doi.org/10.3390/metabo13070797
APA StyleFan, Q., Du, E., Chen, F., Tao, W., Zhao, N., Huang, S., Guo, W., Huang, J., & Wei, J. (2023). Maternal Magnolol Supplementation during Pregnancy and Lactation Promotes Antioxidant Capacity, Improves Gut Health, and Alters Gut Microbiota and Metabolites of Weanling Piglets. Metabolites, 13(7), 797. https://doi.org/10.3390/metabo13070797