

The Effect of MSTN Mutation on Bile Acid Metabolism and Lipid Metabolism in Cattle
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Sample Collection
2.2. Fatty Acid Content Analysis in Liver
2.3. Targeted Metabolomic Analysis of Bile Acids in Liver Tissues and Ileal Contents
2.4. Non-Targeted Lipid Metabolic Analysis of the Liver
2.5. Real-Time PCR
2.6. Western Blot
2.7. Statistical Analysis
3. Results
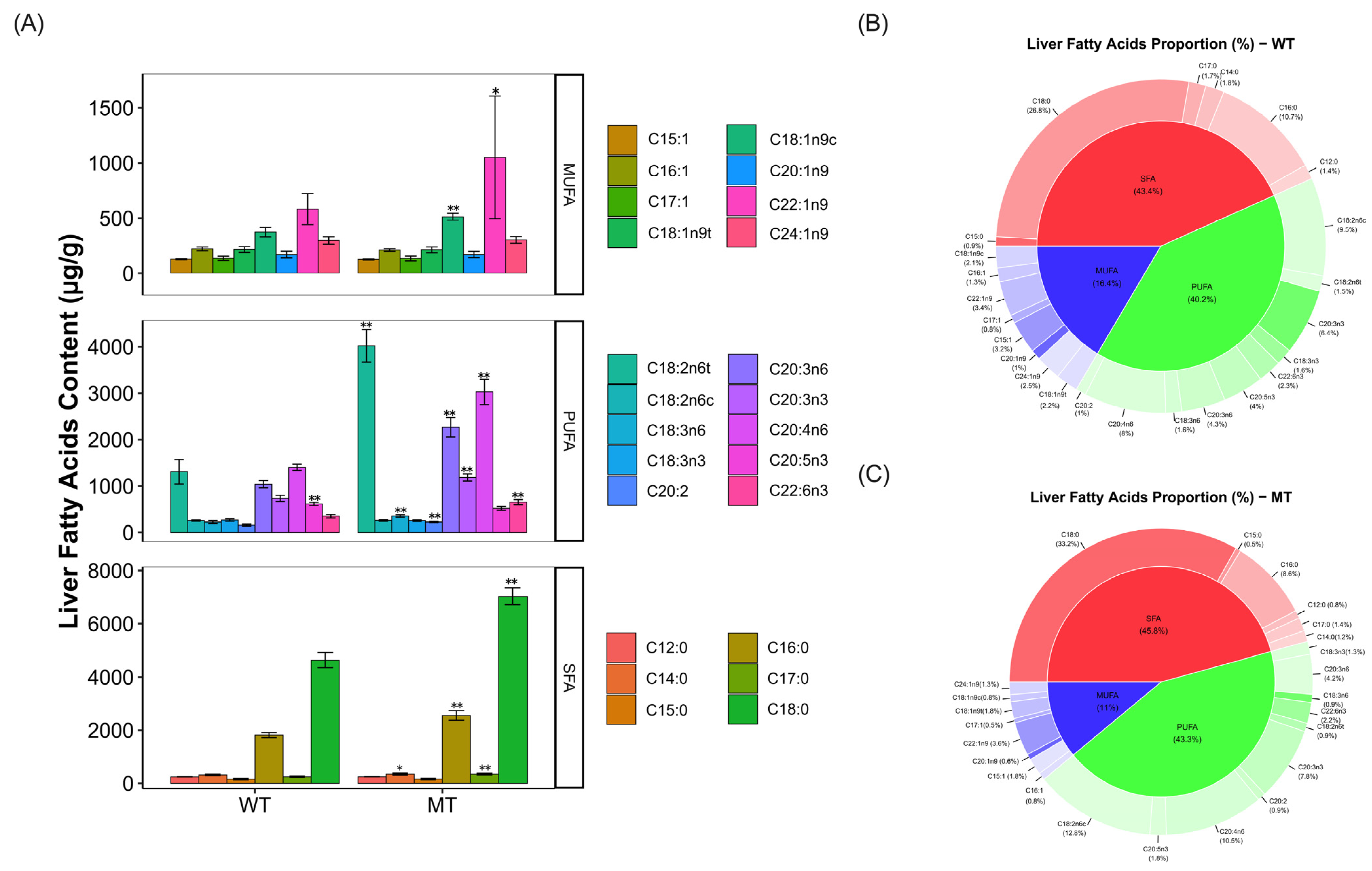
3.1. Analysis of Fatty Acid Content and Composition in Livers of the Cattle
3.2. Analysis of Bile Acid Content and Proportion in Liver and Ileal Contents of the Cattle
3.3. Non-Targeted Analysis of Liver Lipid Metabolism
3.4. Analysis of Gene Expression Related to Bile Acid Metabolism and Lipid Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McPherron, A.C.; Lawler, A.M.; Lee, S.-J. Regulation of skeletal muscle mass in mice by a new TGF-p superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lee, S.-J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Luo, Z.; Han, S.; Li, Z.; Choe, H.; Paek, H.; Quan, B.; Kang, J.; Yin, X. Analysis of meat color, meat tenderness and fatty acid composition of meat in second filial hybrid offspring of MSTN mutant pigs. Meat Sci. 2022, 193, 108929. [Google Scholar] [CrossRef] [PubMed]
- Budsuren, U.; Ulaangerel, T.; Shen, Y.; Liu, G.; Davshilt, T.; Yi, M.; Bold, D.; Zhang, X.; Bai, D.; Dorjgotov, D.; et al. MSTN Regulatory Network in Mongolian Horse Muscle Satellite Cells Revealed with miRNA Interference Technologies. Genes 2022, 13, 1836. [Google Scholar] [CrossRef]
- Bagatoli, A.; Gasparino, E.; Soares, M.; Amaral, R.; Macedo, F.; Voltolini, D.; Del Vesco, A. Expression of calpastatin and myostatin genes associated with lamb meat quality. Genet. Mol. Res. GMR 2013, 12, 6168–6175. [Google Scholar] [CrossRef]
- Gim, G.; Uhm, K.; Kwon, D.; Kim, M.; Jung, D.; Kim, D.; Yi, J.; Ha, J.; Yum, S.; Son, W.; et al. Germline transmission of MSTN knockout cattle via CRISPR-Cas9. Theriogenology 2022, 192, 22–27. [Google Scholar] [CrossRef]
- Konovalova, E.; Romanenkova, O.; Zimina, A.; Volkova, V.; Sermyagin, A. Genetic Variations and Haplotypic Diversity in the Myostatin Gene of Different Cattle Breeds in Russia. Animals 2021, 11, 2810. [Google Scholar] [CrossRef]
- Jakaria, J.; Ladhunka Nur Aliyya, W.; Ismail, R.; Yuni Siswanti, S.; Fakhrul Ulum, M.; Priyanto, R. Discovery of SNPs and indel 11-bp of the myostatin gene and its association with the double-muscled phenotype in Belgian blue crossbred cattle. Gene 2021, 784, 145598. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, L.; Su, G.; Wei, Z.; Liu, X.; Song, L.; Hai, C.; Wu, D.; Hao, Z.; Wu, Y.; et al. Growth Traits and Sperm Proteomics Analyses of Myostatin Gene-Edited Chinese Yellow Cattle. Life 2022, 12, 627. [Google Scholar] [CrossRef]
- Zhu, L.; Bai, C.; Wang, X.; Wei, Z.; Gu, M.; Zhou, X.; Su, G.; Liu, X.; Yang, L.; Li, G. Myostatin Knockout Limits Exercise-Induced Reduction in Bovine Erythrocyte Oxidative Stress by Enhancing the Efficiency of the Pentose Phosphate Pathway. Animals 2022, 12, 927. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, X.; Wei, Z.; Yang, M.; Zhou, X.; Lei, J.; Bai, C.; Su, G.; Liu, X.; Yang, L.; et al. Myostatin Deficiency Enhances Antioxidant Capacity of Bovine Muscle via the SMAD-AMPK-G6PD Pathway. Oxidative Med. Cell. Longev. 2022, 2022, 3497644. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Zhou, X.; Zhu, L.; Gao, Y.; Gao, L.; Bai, C.; Yang, L.; Li, G. viaMyostatin Mutation Promotes Glycolysis by Increasing Phosphorylation of Phosphofructokinase Activation of PDE5A-cGMP-PKG in Cattle Heart. Front. Cell Dev. Biol. 2021, 9, 774185. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Qian, L.; Jiang, S.; Sun, Y.; Wang, Q.; Ma, D.; Xiao, G.; Li, B.; Xie, S.; Gao, T.; et al. Loss-of-function myostatin mutation increases insulin sensitivity and browning of white fat in Meishan pigs. Oncotarget 2017, 8, 34911–34922. [Google Scholar] [CrossRef] [Green Version]
- Trefts, E.; Gannon, M.; Wasserman, D. The liver. Curr. Biol. CB 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Russell, D. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, J.; Soroka, C. Bile formation and secretion: An update. J. Hepatol. 2021, 75, 190–201. [Google Scholar] [CrossRef]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Guzior, D.; Quinn, R. Review: Microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef]
- de Aguiar Vallim, T.; Tarling, E.; Edwards, P. Pleiotropic roles of bile acids in metabolism. Cell Metab. 2013, 17, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Zheng, X.; Ma, X.; Jiang, R.; Zhou, W.; Zhou, S.; Zhang, Y.; Lei, S.; Wang, S.; Kuang, J.; et al. Theabrownin from Pu-erh tea attenuates hypercholesterolemia via modulation of gut microbiota and bile acid metabolism. Nat. Commun. 2019, 10, 4971. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, J.; Zhang, Z.; Xie, H.; Zhang, C.; Bai, Y.; Cao, H.; Che, Q.; Guo, J.; Su, Z. Effect of different bile acids on the intestine through enterohepatic circulation based on FXR. Gut Microbes 2021, 13, 1949095. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, S.; Ahmad, T.; Argueta, D.; Perez, P.; Zhao, C.; Schwartz, G.; DiPatrizio, N.; Haeusler, R. Bile acid composition regulates GPR119-dependent intestinal lipid sensing and food intake regulation in mice. Gut 2020, 69, 1620–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, B.; Sedgeman, L.; Williams, K.; Morand, P.; Cheng, A.; Jarrett, K.; Chan, A.; Brearley-Sholto, M.; Wahlström, A.; Ashby, J.; et al. FXR activation protects against NAFLD via bile-acid-dependent reductions in lipid absorption. Cell Metab. 2021, 33, 1671–1684.e1674. [Google Scholar] [CrossRef]
- Wu, D.; Gu, M.; Wei, Z.; Bai, C.; Su, G.; Liu, X.; Zhao, Y.; Yang, L.; Li, G. Myostatin Knockout Regulates Bile Acid Metabolism by Promoting Bile Acid Synthesis in Cattle. Animals 2022, 12, 205. [Google Scholar] [CrossRef]
- Gao, L.; Yang, M.; Wei, Z.; Gu, M.; Yang, L.; Bai, C.; Wu, Y.; Li, G. MSTN Mutant Promotes Myogenic Differentiation by Increasing Demethylase Expression via the SMAD2/SMAD3 Pathway. Int. J. Biol. Sci. 2020, 16, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- GB/T 19477-2018; Operating procedure of livestock and poultry slaughtering—Cattle. Standards Press of China: Beijing, China, 2018.
- Want, E.J.; Coen, M.; Masson, P.; Keun, H.C.; Pearce, J.T.; Reily, M.D.; Robertson, D.G.; Rohde, C.M.; Holmes, E.; Lindon, J.C.; et al. Ultra performance liquid chromatography-mass spectrometry profiling of bile acid metabolites in biofluids: Application to experimental toxicology studies. Anal. Chem. 2010, 82, 5282–5289. [Google Scholar] [CrossRef]
- Smith, C.; Want, E.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Kuhl, C.; Tautenhahn, R.; Böttcher, C.; Larson, T.; Neumann, S. CAMERA: An integrated strategy for compound spectra extraction and annotation of liquid chromatography/mass spectrometry data sets. Anal. Chem. 2012, 84, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. metaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [Green Version]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Fiems, L. Double Muscling in Cattle: Genes, Husbandry, Carcasses and Meat. Animals 2012, 2, 472–506. [Google Scholar] [CrossRef] [Green Version]
- Fakhfakh, R.; Michaud, A.; Tremblay, J. Blocking the myostatin signal with a dominant negative receptor improves the success of human myoblast transplantation in dystrophic mice. Mol. Ther. J. Am. Soc. Gene Ther. 2011, 19, 204–210. [Google Scholar] [CrossRef]
- Fiems, L.; Vanacker, J.; De Boever, J.; van Caelenbergh, W.; Aerts, J.; De Brabander, D. Effect of energy restriction and re-alimentation in Belgian Blue double-muscled beef cows on digestibility and metabolites. J. Anim. Physiol. Anim. Nutr. 2007, 91, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Fiems, L.; Cottyn, B.; Boucqué, C.V.; Bogaerts, D.; Van Eenaeme, C.; Vanacker, J. Effect of beef type, body weight and dietary protein content on voluntary feed intake, digestibility, blood and urine metabolites and nitrogen retention. J. Anim. Physiol. Anim. Nutr. 1997, 77, 1–9. [Google Scholar] [CrossRef]
- Bittante, G.; Cecchinato, A.; Tagliapietra, F.; Verdiglione, R.; Simonetto, A.; Schiavon, S. Crossbred young bulls and heifers sired by double-muscled Piemontese or Belgian Blue bulls exhibit different effects of sexual dimorphism on fattening performance and muscularity but not on meat quality traits. Meat Sci. 2018, 137, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Fiems, L.; De Campeneere, S.; Van Caelenbergh, W.; De Boever, J.; Vanacker, J. Carcass and meat quality in double-muscled Belgian Blue bulls and cows. Meat Sci. 2003, 63, 345–352. [Google Scholar] [CrossRef]
- Allais, S.; Levéziel, H.; Payet-Duprat, N.; Hocquette, J.-F.; Lepetit, J.; Rousset, S.; Denoyelle, C.; Bernard-Capel, C.; Journaux, L.; Bonnot, A. The two mutations, Q204X and nt821, of the myostatin gene affect carcass and meat quality in young heterozygous bulls of French beef breeds. J. Anim. Sci. 2010, 88, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Wiener, P.; Woolliams, J.; Frank-Lawale, A.; Ryan, M.; Richardson, R.; Nute, G.; Wood, J.; Homer, D.; Williams, J. The effects of a mutation in the myostatin gene on meat and carcass quality. Meat Sci. 2009, 83, 127–134. [Google Scholar] [CrossRef]
- He, N.; Lang, X.; Wang, C.; Lv, C.; Li, M.; Sun, R.; Zhang, J. Expression of MSTN/Smad signaling pathway genes and its association with meat quality in Tibetan sheep (Ovis aries). Food Sci. Nutr. 2023, 11, 1836–1845. [Google Scholar] [CrossRef]
- Chen, Z.; Herdt, T.H.; Liesman, J.S.; Ames, N.K.; Emery, R.S. Reduction of bovine plasma cholesterol concentration by partial interruption of enterohepatic circulation of bile salts: A novel hypocholesterolemic model. J. Lipid Res. 1995, 36, 1544–1556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, L.; Ke, S.; Chen, X.; Kenéz, Á.; Xu, W.; Wang, D.; Zhang, F.; Li, Y.; Cui, Z.; et al. Yak rumen microbiome elevates fiber degradation ability and alters rumen fermentation pattern to increase feed efficiency. Anim. Nutr. 2022, 11, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Jiao, T.; Liu, W.; Luo, Y.; Wang, J.; Guo, X.; Tong, X.; Lin, Z.; Sun, C.; Wang, K.; et al. Hepatic cytochrome P450 8B1 and cholic acid potentiate intestinal epithelial injury in colitis by suppressing intestinal stem cell renewal. Cell Stem Cell 2022, 29, 1366–1381.e1369. [Google Scholar] [CrossRef] [PubMed]
- Ticho, A.; Malhotra, P.; Dudeja, P.; Gill, R.; Alrefai, W. Intestinal Absorption of Bile Acids in Health and Disease. Compr. Physiol. 2019, 10, 21–56. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Lai, Z.; Yang, H.; Zhang, J.; Qi, W.; Xie, F.; Mao, S. Genome-centric investigation of bile acid metabolizing microbiota of dairy cows and associated diet-induced functional implications. Isme J 2023, 17, 172–184. [Google Scholar] [CrossRef]
- Wilkes, J.; Lloyd, D.; Gekakis, N. Loss-of-function mutation in myostatin reduces tumor necrosis factor alpha production and protects liver against obesity-induced insulin resistance. Diabetes 2009, 58, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Williams, N.G.; Interlichia, J.P.; Jackson, M.F.; Hwang, D.; Cohen, P.; Rodgers, B.D. Endocrine actions of myostatin: Systemic regulation of the IGF and IGF binding protein axis. Endocrinology 2011, 152, 172–180. [Google Scholar] [CrossRef]
- Chiang, J.; Ferrell, J. Bile Acids as Metabolic Regulators and Nutrient Sensors. Annu. Rev. Nutr. 2019, 39, 175–200. [Google Scholar] [CrossRef]
- Fuchs, C.; Trauner, M. Role of bile acids and their receptors in gastrointestinal and hepatic pathophysiology. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 432–450. [Google Scholar] [CrossRef]
- Allen, D.L.; Cleary, A.S.; Speaker, K.J.; Lindsay, S.F.; Uyenishi, J.; Reed, J.M.; Madden, M.C.; Mehan, R.S. Myostatin, activin receptor IIb, and follistatin-like-3 gene expression are altered in adipose tissue and skeletal muscle of obese mice. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E918–E927. [Google Scholar] [CrossRef] [Green Version]
- Grade, C.V.C.; Mantovani, C.S.; Fontoura, M.A.; Yusuf, F.; Brand-Saberi, B.; Alvares, L.E. CREB, NF-Y and MEIS1 conserved binding sites are essential to balance Myostatin promoter/enhancer activity during early myogenesis. Mol. Biol. Rep. 2017, 44, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Simbrunner, B.; Trauner, M.; Reiberger, T. Review article: Therapeutic aspects of bile acid signalling in the gut-liver axis. Aliment. Pharmacol. Ther. 2021, 54, 1243–1262. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.; Ferrell, J. Discovery of farnesoid X receptor and its role in bile acid metabolism. Mol. Cell. Endocrinol. 2022, 548, 111618. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, B.; Jones, S.; Price, R.; Watson, M.; McKee, D.; Moore, L.; Galardi, C.; Wilson, J.; Lewis, M.; Roth, M.; et al. A regulatory cascade of the nuclear receptors FXR, SHP-1, and LRH-1 represses bile acid biosynthesis. Mol. Cell 2000, 6, 517–526. [Google Scholar] [CrossRef]
- Somm, E.; Henry, H.; Bruce, S.; Bonnet, N.; Montandon, S.; Niederländer, N.; Messina, A.; Aeby, S.; Rosikiewicz, M.; Fajas, L.; et al. β-Klotho deficiency shifts the gut-liver bile acid axis and induces hepatic alterations in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E833–E847. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Crudele, A.; Panera, N.; Romito, I.; Meroni, M.; De Stefanis, C.; Palma, A.; Comparcola, D.; Fracanzani, A.; Miele, L.; et al. β-Klotho gene variation is associated with liver damage in children with NAFLD. J. Hepatol. 2020, 72, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Gulfo, J.; Rotondo, F.; Ávalos de León, C.G.; Cornide-Petronio, M.E.; Fuster, C.; Gracia-Sancho, J.; Jiménez-Castro, M.B.; Peralta, C. FGF15 improves outcomes after brain dead donor liver transplantation with steatotic and non-steatotic grafts in rats. J. Hepatol. 2020, 73, 1131–1143. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, X.X.; Scherzer, P.; Wilson, P.; Tallman, J.; Takahashi, H.; Li, J.; Iwahashi, M.; Sutherland, E.; Arend, L.; et al. Farnesoid X receptor modulates renal lipid metabolism, fibrosis, and diabetic nephropathy. Diabetes 2007, 56, 2485–2493. [Google Scholar] [CrossRef] [Green Version]
- Dodson, M.V.; Hausman, G.J.; Guan, L.; Du, M.; Rasmussen, T.P.; Poulos, S.P.; Mir, P.; Bergen, W.G.; Fernyhough, M.E.; McFarland, D.C.; et al. Lipid metabolism, adipocyte depot physiology and utilization of meat animals as experimental models for metabolic research. Int. J. Biol. Sci. 2010, 6, 691–699. [Google Scholar] [CrossRef]
- Liang, Z.; Zhang, J.; Du, M.; Ahmad, A.A.; Wang, S.; Zheng, J.; Salekdeh, G.H.; Yan, P.; Han, J.; Tong, B.; et al. Age-dependent changes of hindgut microbiota succession and metabolic function of Mongolian cattle in the semi-arid rangelands. Front. Microbiol. 2022, 13, 957341. [Google Scholar] [CrossRef]
- Mancin, L.; Wu, G.D.; Paoli, A. Gut microbiota-bile acid-skeletal muscle axis. Trends Microbiol. 2023, 31, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Heymans, C.; Heij, L.R.; Lenaerts, K.; den Dulk, M.; Hadfoune, M.; van Heugten, C.; Spiller, O.B.; Beeton, M.L.; Stock, S.J.; Jobe, A.H.; et al. Prophylactic Intra-Uterine β-Cyclodextrin Administration during Intra-Uterine Ureaplasma parvum Infection Partly Prevents Liver Inflammation without Interfering with the Enterohepatic Circulation of the Fetal Sheep. Nutrients 2020, 12, 1312. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, D.; Wang, S.; Hai, C.; Wang, L.; Pei, D.; Bai, C.; Su, G.; Liu, X.; Zhao, Y.; Liu, Z.; et al. The Effect of MSTN Mutation on Bile Acid Metabolism and Lipid Metabolism in Cattle. Metabolites 2023, 13, 836. https://doi.org/10.3390/metabo13070836
Wu D, Wang S, Hai C, Wang L, Pei D, Bai C, Su G, Liu X, Zhao Y, Liu Z, et al. The Effect of MSTN Mutation on Bile Acid Metabolism and Lipid Metabolism in Cattle. Metabolites. 2023; 13(7):836. https://doi.org/10.3390/metabo13070836
Chicago/Turabian StyleWu, Di, Song Wang, Chao Hai, Linfeng Wang, Dongchao Pei, Chunling Bai, Guanghua Su, Xuefei Liu, Yuefang Zhao, Zhonghua Liu, and et al. 2023. "The Effect of MSTN Mutation on Bile Acid Metabolism and Lipid Metabolism in Cattle" Metabolites 13, no. 7: 836. https://doi.org/10.3390/metabo13070836