
The Urine Metabolome of R6/2 and zQ175DN Huntington’s Disease Mouse Models

 ,
,  ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. SIL Stock and Working Solution Preparation
2.3. Animals and Study Design
2.3.1. R6/2 Mice
2.3.2. zQ175DN KI Mice
2.3.3. Study Design and Urine Collection
2.4. Mouse Urine Normalization via Specific Gravity
2.5. Urine Sample Preparation
2.6. The Preparation and Analysis of Quality Control Samples and Blanks
2.7. UHPLC–High-Resolution Mass Spectrometry Analysis
2.7.1. HILIC and Reverse-Phase Chromatography
2.7.2. Mass Spectrometry
2.8. Data Analysis
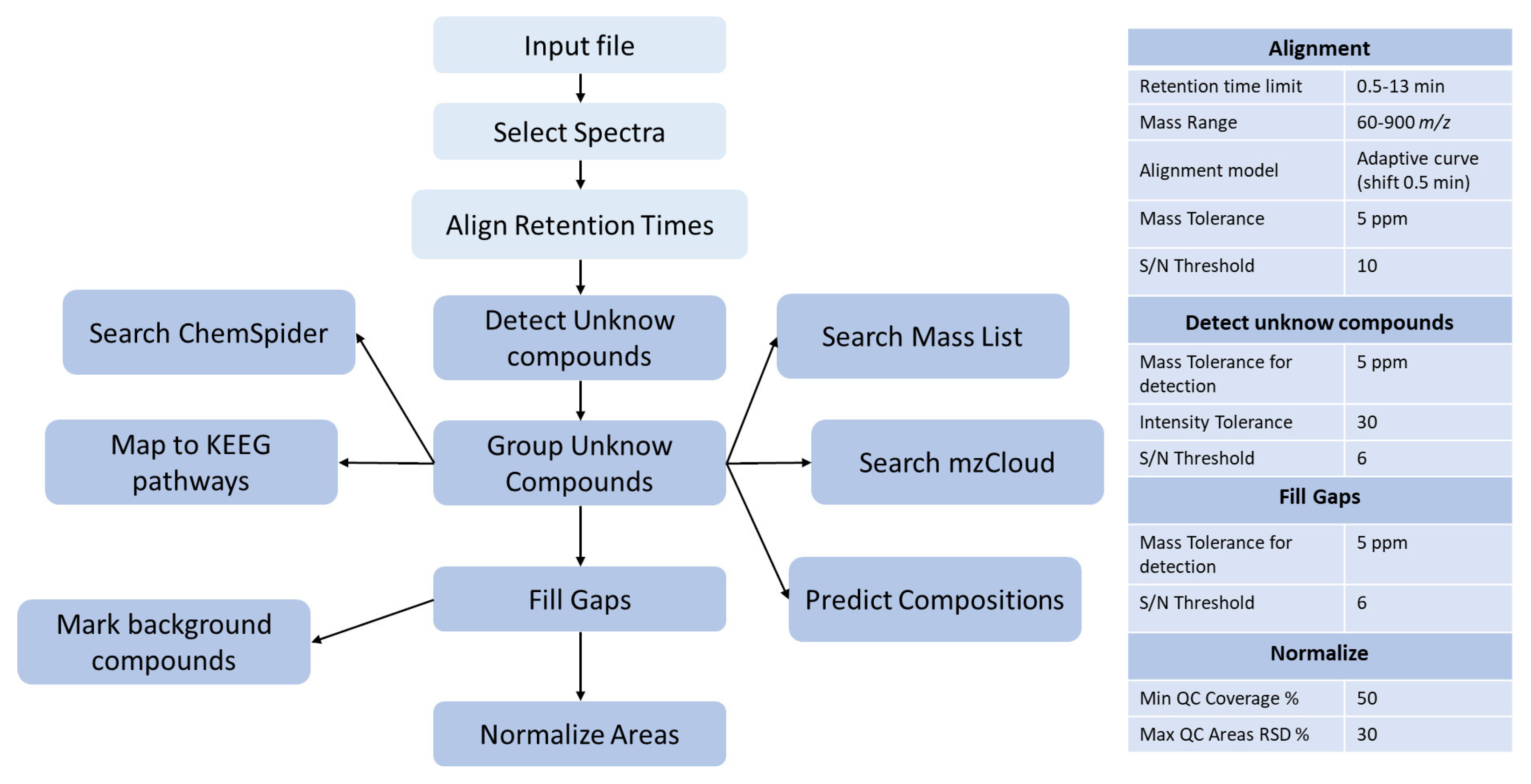
2.8.1. Raw Data Processing Using Compound Discoverer
2.8.2. Raw Data Filtration
2.8.3. Multivariate Statistical Analysis (SIMCA)
2.8.4. Putative Metabolite Identification
2.8.5. Metabolite Confirmation with Synthetic Standards
3. Results
3.1. Mouse Urine Samples’ Normalization via Specific Gravity
3.2. Analytical Quality Assurance
3.3. The Acquisition and Filtering of R6/2 and zQ175DN Datasets
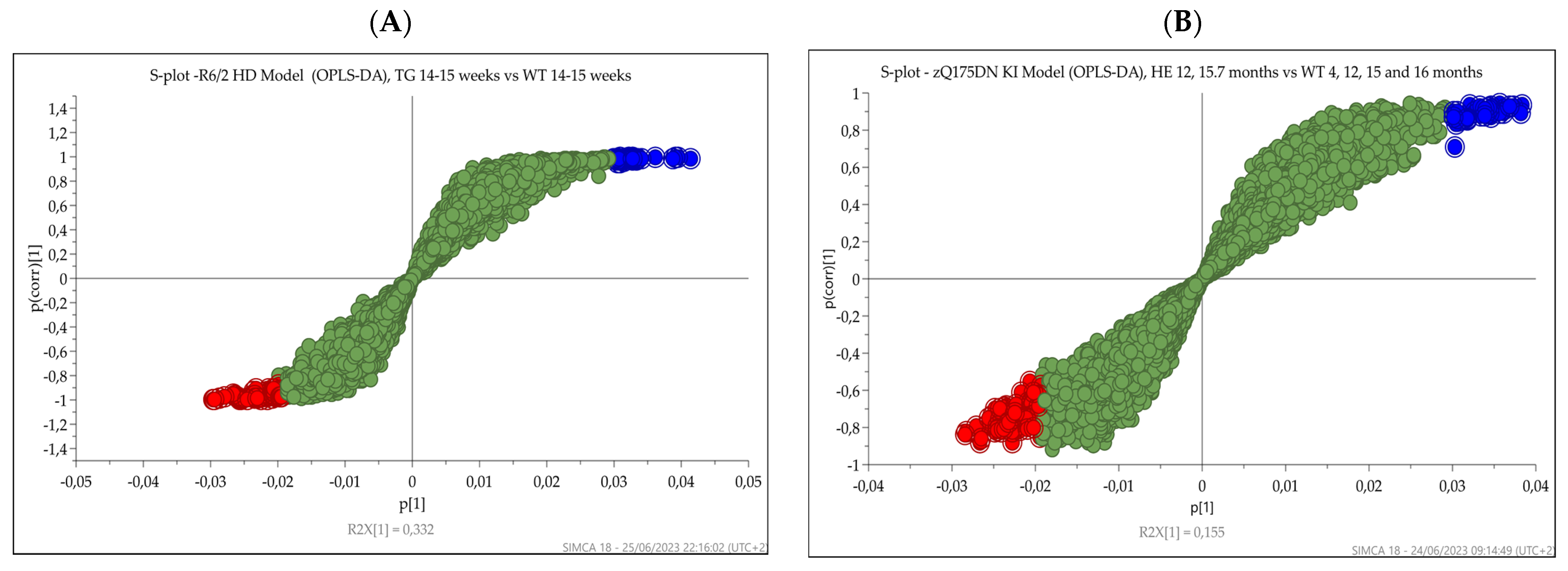
3.4. Multivariate Analysis
3.5. Urine Metabolite Alterations in 3-Month-Old R6/2 versus Wild-Type Mice
3.6. Urine Metabolite Alterations in zQ175DN versus Age-Matched Wild-Type Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saudou, F.; Humbert, S. The biology of huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.A.; Tabrizi, S.J. Huntington’s disease: From molecular pathogenesis to clinical treatment. Lancet Neurol. 2011, 10, 83–98. [Google Scholar] [CrossRef]
- Bates, G.P.; Jones, L. Huntington’s Disease; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Rubinsztein, D.C.; Leggo, J.; Coles, R.; Almqvist, E.; Biancalana, V.; Cassiman, J.J.; Chorai, K.; Connarty, M.; Crauford, D.; Curtis, A.; et al. Phenotypic characterization of individuals with 30-40 CAG repeats in the Huntington disease (HD) gene reveals HD cases with 36 repeats and apparently normal elderly individuals with 36-39 repeats. Am. J. Hum. Genet. 1996, 59, 16–22. [Google Scholar] [PubMed]
- The Huntington’s Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Figiel, M.; Szlachcic, W.J.; Switonski, P.M.; Gabka, A.; Krzyzosiak, W.J. Mouse models of polyglutamine diseases: Review and data table. Part I. Mol. Neurobiol. 2012, 46, 393–429. [Google Scholar] [CrossRef] [PubMed]
- Heng, M.Y.; Detloff, P.J.; Albin, R.L. Rodent genetic models of Huntington disease. Neurobiol. Dis. 2008, 32, 1–9. [Google Scholar] [CrossRef]
- Pouladi, M.A.; Morton, A.J.; Hayden, M.R. Choosing an animal model for the study of Huntington’s disease. Nat. Rev. Neurosci. 2013, 14, 708–721. [Google Scholar] [CrossRef]
- Mangiarini, L.; Sathasivam, K.; Seller, M.; Cozens, B.; Harper, A.; Hetherington, C.; Lawton, M.; Trottier, Y.; Lehrach, H.; Davies, S.W.; et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell 1996, 87, 493–506. [Google Scholar] [CrossRef]
- Menalled, L.; El-Khodor, B.F.; Patry, M.; Suarez-Farinas, M.; Orenstein, S.; Zahasky, B.; Leahy, C.; Wheeler, V.X.; Yang, W.; MacDonald, M.; et al. Systematic behavioral evaluation of Huntington’s disease transgenic and knock-in mouse models. Neurobiol. Dis. 2009, 35, 319–336. [Google Scholar] [CrossRef]
- Hult, S.; Soylu, R.; Björklund, T.; Belgardt, B.F.; Mauer, J.; Brüning, J.C.; Kirik, D.; Petersén, Å. Mutant huntingtin causes metabolic imbalance by disruption of hypothalamic neurocircuits. Cell Metab. 2011, 13, 428–439. [Google Scholar] [CrossRef]
- Björkqvist, M.; Petersen, A.; Bacos, K.; Isaacs, J.; Norlen, P.; Gil, J.; Popovic, N.; Sundler, F.; Bates, G.P.; Tabrizi, S.J.; et al. Progressive alterations in the hypothalamic-pituitary-adrenal axis in the R6/2 transgenic mouse model of Huntington’s disease. Hum. Mol. Genet. 2006, 15, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Van Wamelen, D.J.; Aziz, N.A.; Roos, R.A.C.; Swaab, D.F. Hypothalamic Alterations in Huntington’s Disease Patients: Comparison with Genetic Rodent Models. J. Neuroendocrinol. 2014, 26, 761–775. [Google Scholar] [CrossRef]
- Dufour, B.D.; McBride, J.L. Corticosterone dysregulation exacerbates disease progression in the R6/2 transgenic mouse model of Huntington’s disease. Exp. Neurol. 2016, 283 Pt A, 308–317. [Google Scholar] [CrossRef]
- Southwell, A.L.; Smith-Dijak, A.; Kay, C.; Sepers, M.; Villanueva, E.B.; Parsons, M.P.; Xie, Y.; Anderson, L.; Felczak, B.; Waltl, S.; et al. An enhanced Q175 knock-in mouse model of Huntington disease with higher mutant huntingtin levels and accelerated disease phenotypes. Hum. Mol. Genet. 2016, 25, 3654–3675. [Google Scholar] [CrossRef] [PubMed]
- Khamis, M.M.; Adamko, D.J.; El-Aneed, A. Mass spectrometric based approaches in urine metabolomics and biomarker discovery. Mass Spectrom. Rev. 2017, 36, 115–134. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Modern analytical techniques in metabolomics analysis. Analyst 2012, 137, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wu, X.; Wang, X. Urine metabolomics. Clin. Chim. Acta 2012, 414, 65–69. [Google Scholar] [CrossRef]
- Petrella, G.; Montesano, C.; Lentini, S.; Ciufolini, G.; Vanni, D.; Speziale, R.; Salonia, A.; Montorsi, F.; Summa, V.; Vago, R. Personalized metabolic profile by synergic use of NMR and HRMS. Molecules 2021, 26, 4167. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S.; et al. HMDB: A knowledgebase for the human metabolome. Nucleic Acids Res. 2009, 37, D603–D610. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]
- Sana, T.R.; Roark, J.C.; Li, X.; Waddell, K.; Fischer, S.M. Molecular formula and METLIN Personal Metabolite Database matching applied to the identification of compounds generated by LC/TOF-MS. J. Biomol. Tech. 2008, 19, 258–266. [Google Scholar] [PubMed]
- Gagnebin, Y.; Tonoli, D.; Lescuyer, P.; Ponte, B.; de Seigneux, S.; Martin, P.Y.; Schappler, J.; Boccard, J.; Rudaz, S. Metabolomic analysis of urine samples by UHPLC-QTOF-MS: Impact of normalization strategies. Anal. Chim. Acta 2017, 955, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Edmands, W.M.B.; Ferrari, P.; Scalbert, A. Normalization to specific gravity prior to analysis improves information recovery from high-resolution mass spectrometry metabolomic profiles of human urine. Anal. Chem. 2014, 86, 10925–10931. [Google Scholar] [CrossRef] [PubMed]
- Croixmarie, V.; Umbdenstock, T.; Cloarec, O.; Moreau, A.; Pascussi, J.M.; Boursier-Neyret, C.; Walther, B. Integrated comparison of drug-related and drug-induced ultra-performance liquid chromatography/mass spectrometry metabonomic profiles using human hepatocyte cultures. Anal. Chem. 2009, 81, 6061–6069. [Google Scholar] [CrossRef] [PubMed]
- Scheltema, R.; Decuypere, S.; Dujardin, J.; Watson, D.; Jansen, R.; Breitling, R. Simple data-reduction method for high-resolution LC-MS data in metabolomics. Bioanalysis 2009, 1, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Saleh, N.; Moutereau, S.; Durr, A.; Krystkowiak, P.; Azulay, J.-P.; Tranchant, C.; Broussolle, E.; Morin, F.; Bachoud-Lévi, A.-C.; Maison, P. Neuroendocrine Disturbances in Huntington’s Disease. PLoS ONE 2009, 4, e4962. [Google Scholar] [CrossRef]
- Nagaraj, H.R.; Portero-Otin, M.; Monnier, V.M. Pyrraline ether crosslinks as a basis for protein crosslinking by the advanced Maillard reaction in aging and diabetes. Arch. Biochem. Biophys. 1996, 325, 152–158. [Google Scholar] [CrossRef]
- Hegele, J.; Rarisod, V.; Richoz, J.; Orster, A.F.; Maurer, S.; Krause, R.; Henle, T.; Bütler, T.; Delatour, T. Evaluating the extent of protein damage in dairy products. Ann. N. Y. Acad. Sci. 2008, 1126, 300–306. [Google Scholar] [CrossRef]
- Björkqvist, M.; Fex, M.; Renström, E.; Wierup, N.; Petersén, A.; Gil, J.; Bacos, K.; Popovic, N.; Li, J.Y.; Sundler, F.; et al. The R6/2 transgenic mouse model of Huntington’s disease develops diabetes due to deficient beta-cell mass and exocytosis. Hum. Mol. Genet. 2005, 14, 565–574. [Google Scholar] [CrossRef]
- Smith, A.; Taneda, S.; Richey, P.L.; Miyata, S.; Yan, S.-D.; Stern, D.; Sayre, L.M.; Monnier, V.M.; Perry, G. Advanced Maillard reaction end products are associated with Alzheimer disease pathology. Proc. Natl. Acad. Sci. USA 1994, 91, 5710–5714. [Google Scholar] [CrossRef]
- Armstrong, S.D.; Robertson, D.H.L.; Cheetham, S.A.; Hurst, J.L.; Beynon, R.J. Structural and functional differences in isoforms of mouse major urinary proteins: A male-specific protein that preferentially binds a male pheromone. Biochem. J. 2005, 391, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Röck, F.; Hadeler, K.-P.; Rammensee, H.-G.; Overath, P. Quantitative analysis of mouse urine volatiles: In search of MHC-dependent differences. PLoS ONE 2007, 5, e429. [Google Scholar] [CrossRef] [PubMed]
- Lou, M.F. Isolation and identification of L-/3-aspartyl-L-lysine and L-y-glutamyl-L-ornithine from normal human urine. Biochemistry 1975, 14, 3503–3508. [Google Scholar] [CrossRef]
- Dlasková, Z.; Navrátil, T.; Heyrovský, M.; Pelclová, D.; Novotný, L. Voltammetric determination of thiodiglycolic acid in urine. Anal. Bioanal. Chem. 2003, 375, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and other tryptophan metabolites: Rhythms Outside the Animal World and Some Novel, Presumably Universal Pathways. In Comparative Aspects of Circadian Rhythms; Fanjul-Moles, M.L., Ed.; Rans World Research Network: Kerala, India, 2008; pp. 1–17. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Measured Mass | Formula | p-Value a | Fold Change a TG vs. WT | Identification | Pathway |
|---|---|---|---|---|---|---|
| ESI- | ||||||
| 7.06 | 246.9715 | C12H8S3 | 2.00 × 10−11 | nf in TG | Terthiophene b | unknown |
| 9.24 | 362.9720 | - | 2.82 × 10−14 | nf in TG | unknown | unknown |
| 5.79 | 422.0610 | - | 7.93 × 10−15 | nf in TG | unknown | unknown |
| 8.76 | 377.1973 | C21H30O6 | 4.68 × 10−6 | 9 | 18,19-dihydroxycorticosterone b | cortiscosterone dysregulation |
| 9.65 | 361.2023 | C21H30O5 | 3.82 × 10−5 | 132 | 18-hydroxycorticosterone b | cortiscosterone dysregulation |
| 9.55 | 365.2336 | C21H34O5 | 3.05 × 10−13 | 4 | Tetrahydrocortisol b | cortiscosterone dysregulation |
| 9.79 | 363.2180 | C21H32O5 | 2.03 × 10−5 | 8 | hydroxy-dehydrotetrahydrocorticosterone b | cortiscosterone dysregulation |
| 9.65 | 408.2117 | C17H32O9N2 | 2.65 × 10−5 | 17 | unknown | unknown |
| ESI+ | ||||||
| 9.19 | 379.2112 | C21H30O6 | 2.35 × 10−6 | 218 | Hydroxycorticosterone b | cortiscosterone dysregulation |
| 9.95 | 361.2005 | C21H29O5 | 6.69 × 10−4 | nf in WT | hydroxy-dehydrocorticosterone b | cortiscosterone dysregulation |
| 6.48 | 255.1336 | C12H18O4N2 | <1.00 × 10−4 | 10 | Pyrraline c | nonenzymatic glycation |
| 2.65 | 255.1336 | C12H18O4N2 | 4.00 × 10−10 | 214 | unknown | unknown |
| 4.84 | 239.1388 | C12H18O4N2 | 2.61 × 10−4 | 3.4 | unknown | unknown |
| 6.71 | 144.0841 | C7H13NS | 9.27 × 10−17 | nf in TG | 2-(sec-butyl)-4,5-dihydrothiazole c | pheromone |
| 2.49 | 304.1500 | C12H21O6N3 | 2.57 × 10−13 | 44 | N-acetyl-aspartyl-lysine b | unknown |
| 6.73 | 382.0769 | C16H15O10N | 5.30 × 10−3 | 1.95 | xanthurenic acid glucuronide b | Trp metabolism |
| 7.51 | 206.0448 | C10H7O4N | <1.00 × 10−4 | 0.12 | xanthurenic acid c | Trp metabolism |
| 8.17 | 188.0739 | C8H14O2NS | 2.93 × 10−6 | 0.001 | N-[(1E)-3-methyl-1,3-butadien-1-yl]-L-cysteine b | unknown |
| RT (min) | Measured Mass | Formula | p-Value a | Fold Change a HE vs. WT | Identification | Pathway |
|---|---|---|---|---|---|---|
| ESI- | ||||||
| 4.82 | 148.9914 | C4H5O4S | 6.37 × 10−6 | 2.7 | thiodiglycolic acid b | oxidative stress |
| 4.43 | 192.0336 | C6H10O4NS | 1.11 × 10−6 | 2.5 | S-2-carboxyethyl-L-cysteine/ 2-((2-Hydroxyethyl)amino)-3-mercapto-4-oxobutanoic acid b | oxidative stress |
| 4.31 | 305.0453 | C10H13O7N2S | 9.35 × 10−6 | nf in WT | deaminated glutathione b | waste compound |
| 9.03 | 378.0837 | C17H16O9N | 1.18 × 10−4 | 3.3 | indole-3-pyruvic acid glucuronide b | Trp metabolism |
| 8.56 | 313.0567 | C13H13O9 | 5.97 × 10−5 | 2.9 | unknown | Unknown |
| 9.59 | 435.1303 | C21H23O10 | 4.51 × 10−4 | 3.7 | unknown | Unknown |
| 9.56 | 421.1510 | C21H25O9 | 3.37 × 10−4 | 4.3 | unknown | Unknown |
| 9.19 | 315.1242 | C18H19O5 | 3.29 × 10−3 | 3.9 | unknown | Unknown |
| ESI+ | ||||||
| 5.10 | 350.1012 | C12H20O7N3S | 4.00 × 10−7 | 4.4 | N-acetylglutathione c | oxidative stress |
| 4.94 | 398.1663 | C16H24O7N5 | 5.62 × 10−6 | 3.9 | carnosine-propanal-aspartic b | oxidative stress |
| 1.32 | 255.0642 | C7H15O6N2S | 2.46 × 10−7 | 1.3 | gamma-glutamyltaurine b | dipeptide |
| 1.65 | 130.1226 | C7H16ON | 8.97 × 10−5 | 0.3 | N-methyl hexanamide b | fatty amides |
| 0.91 | 114.0913 | C4H8ON3 | 2.75 × 10−5 | 0.3 | creatinine c | others |
| 1.18 | 116.1069 | C6H14NO | 1.53 × 10−5 | 0.3 | trimethylaminoacetone c | others |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Speziale, R.; Montesano, C.; Di Pietro, G.; Cicero, D.O.; Summa, V.; Monteagudo, E.; Orsatti, L. The Urine Metabolome of R6/2 and zQ175DN Huntington’s Disease Mouse Models. Metabolites 2023, 13, 961. https://doi.org/10.3390/metabo13080961
Speziale R, Montesano C, Di Pietro G, Cicero DO, Summa V, Monteagudo E, Orsatti L. The Urine Metabolome of R6/2 and zQ175DN Huntington’s Disease Mouse Models. Metabolites. 2023; 13(8):961. https://doi.org/10.3390/metabo13080961
Chicago/Turabian StyleSpeziale, Roberto, Camilla Montesano, Giulia Di Pietro, Daniel Oscar Cicero, Vincenzo Summa, Edith Monteagudo, and Laura Orsatti. 2023. "The Urine Metabolome of R6/2 and zQ175DN Huntington’s Disease Mouse Models" Metabolites 13, no. 8: 961. https://doi.org/10.3390/metabo13080961
APA StyleSpeziale, R., Montesano, C., Di Pietro, G., Cicero, D. O., Summa, V., Monteagudo, E., & Orsatti, L. (2023). The Urine Metabolome of R6/2 and zQ175DN Huntington’s Disease Mouse Models. Metabolites, 13(8), 961. https://doi.org/10.3390/metabo13080961