
Genistein Suppresses IL-6 and MMP-13 to Attenuate Osteoarthritis in Obese Diabetic Mice

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
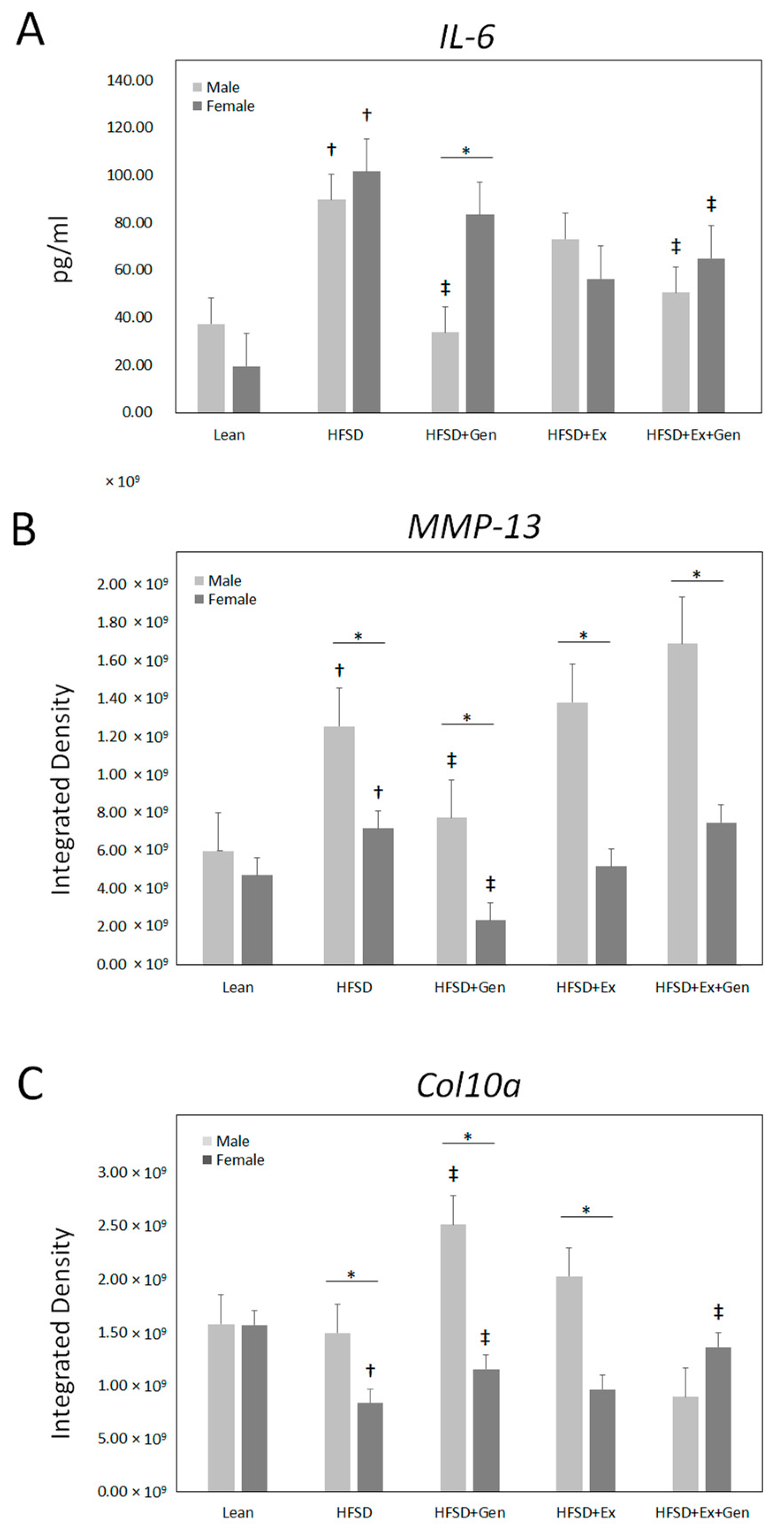
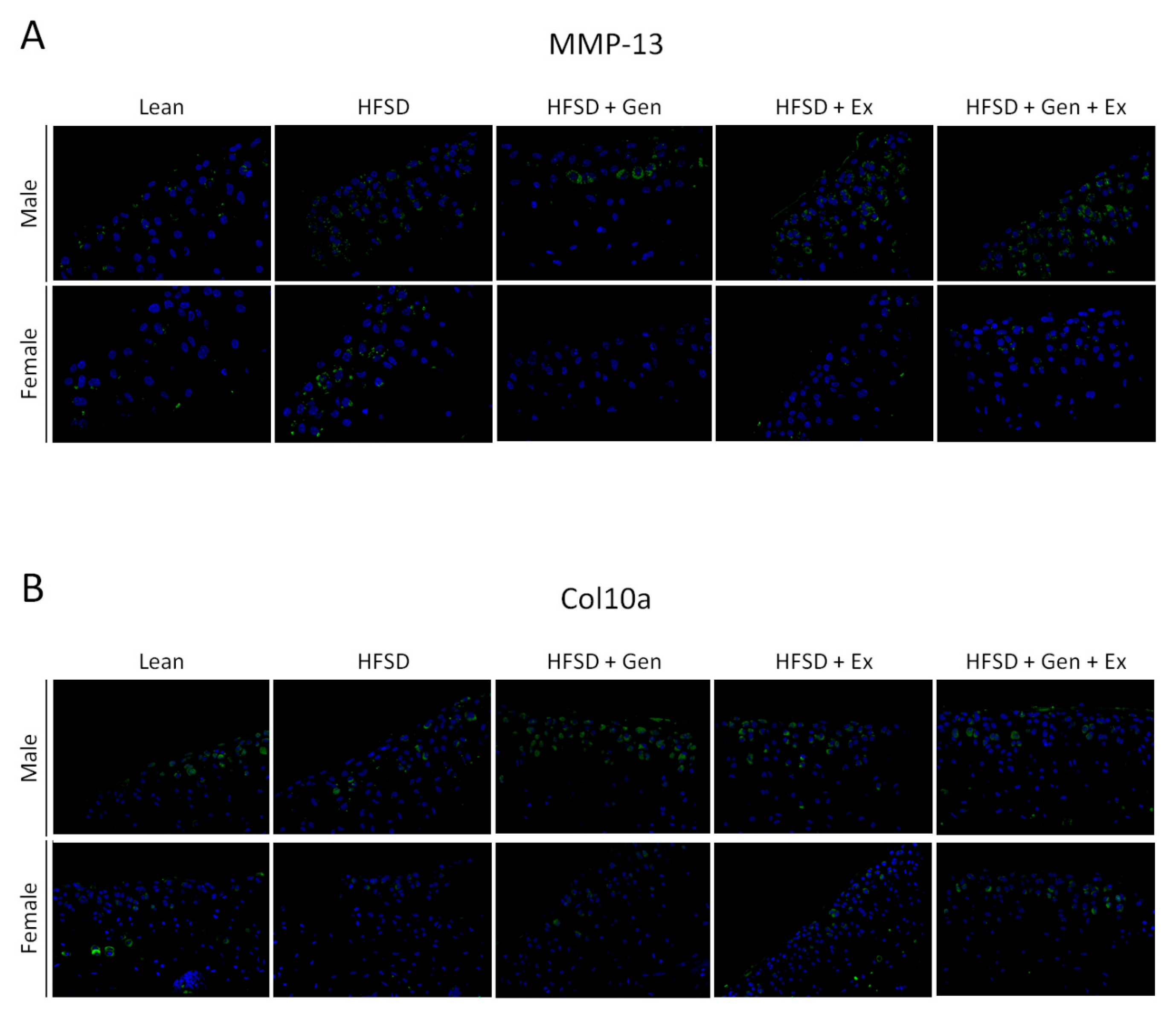
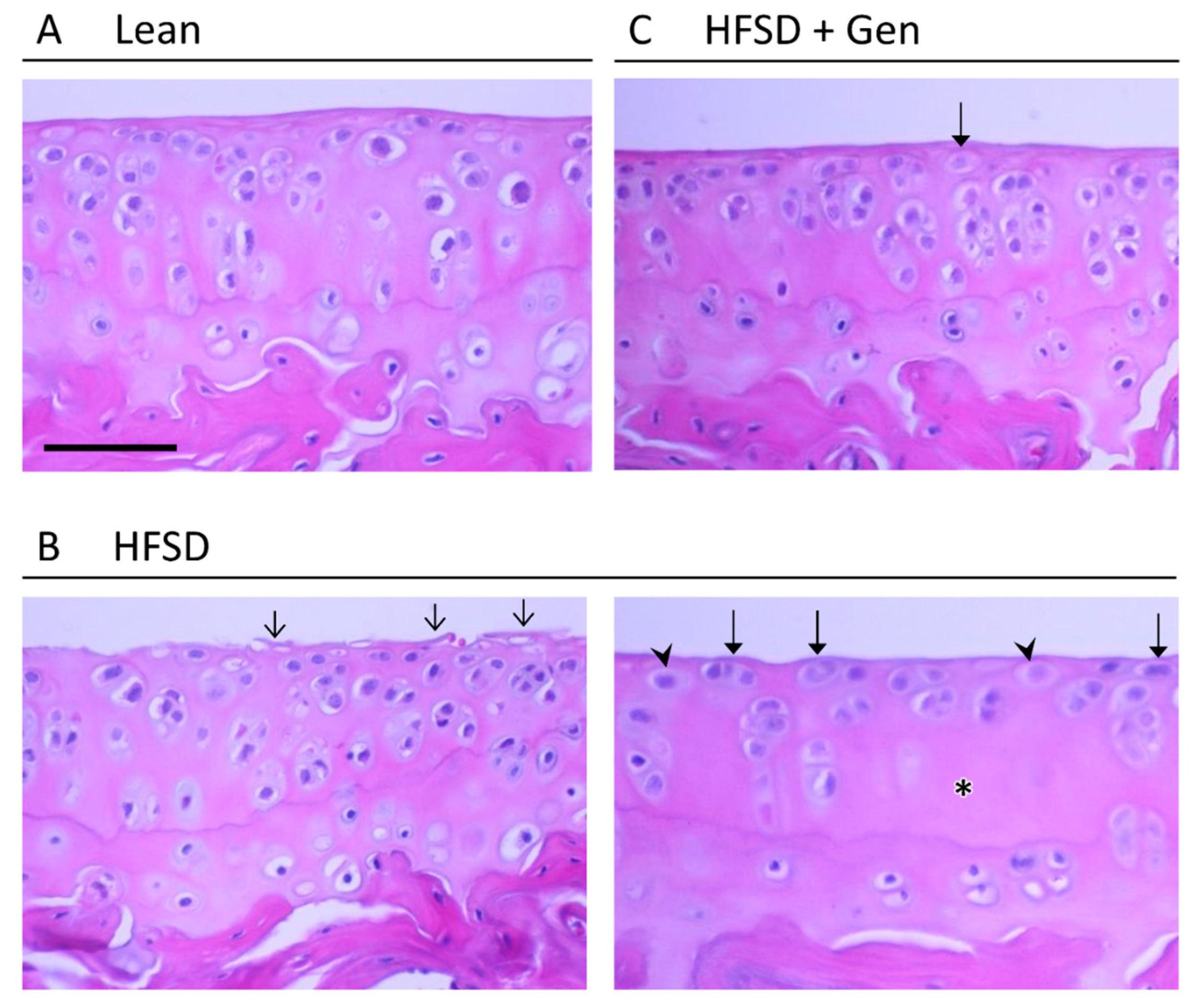
3. Results
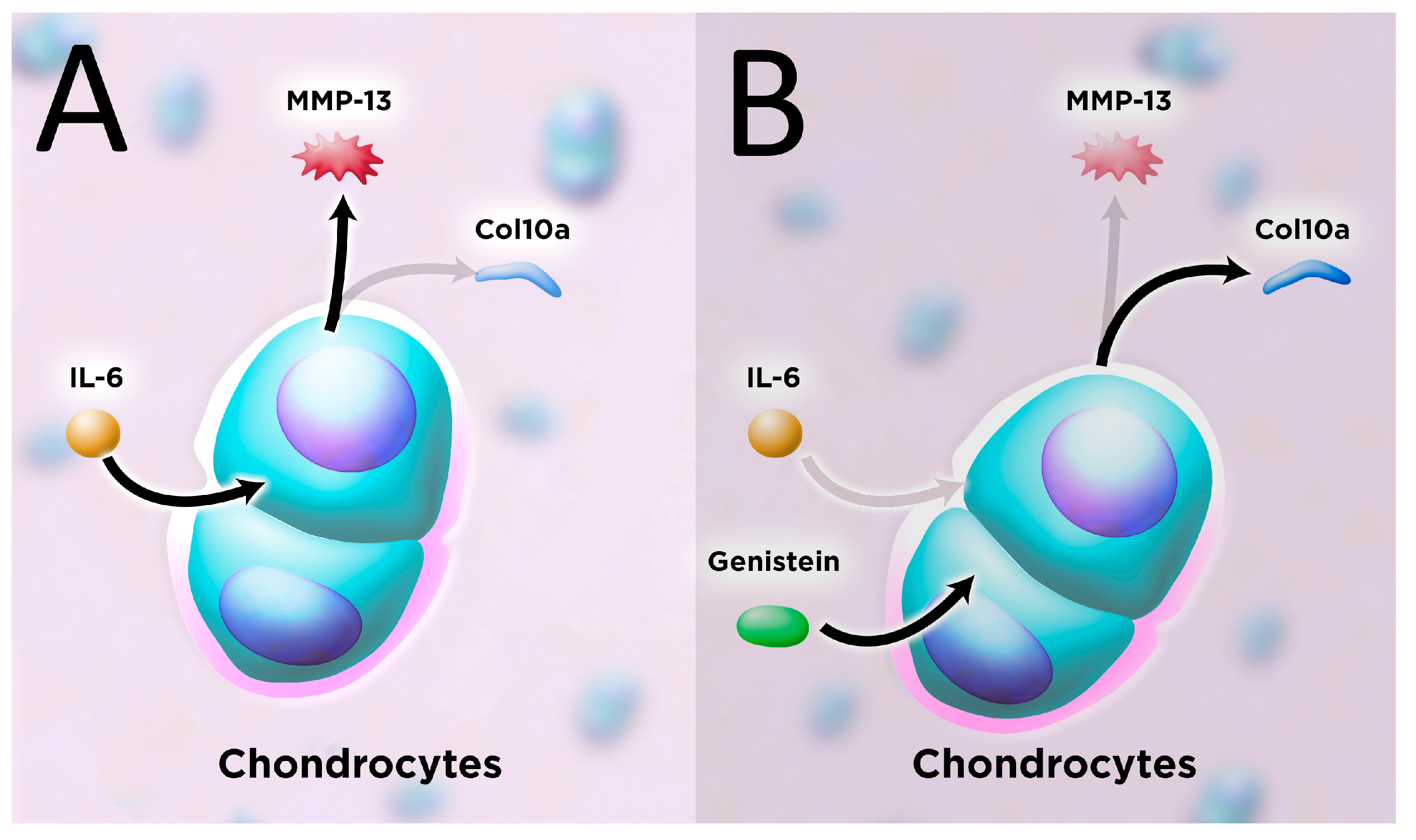
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stürmer, T.; Brenner, H.; Brenner, R.E.; Günther, K.P. Non-insulin dependent diabetes mellitus (NIDDM) and patterns of osteoarthritis. The Ulm osteoarthritis study. Scand. J. Rheumatol. 2001, 30, 169–171. [Google Scholar]
- King, K.B.; Findley, T.W.; Williams, A.E.; Bucknell, A.L. Veterans with diabetes receive arthroplasty more frequently and at a younger age. Clin. Ortho. Rel. Res. 2013, 471, 3049–3054. [Google Scholar]
- Schett, G.; Kleyer, A.; Perricone, C.; Sahinbegovic, E.; Iagnocco, A.; Zwerina, J.; Lorenzini, R.; Aschenbrenner, F.; Berenbaum, F.; D’Agostino, M.A.; et al. Diabetes is an independent predictor for severe osteoarthritis: Results from a longitudinal cohort study. Diabetes Care. 2013, 36, 403–409. [Google Scholar]
- Velasquez, M.T.; Katz, J.D. Osteoarthritis: Another component of metabolic syndrome? Metab. Syndr. Relat. Disord. 2010, 8, 295–305. [Google Scholar]
- Piva, S.R.; Susko, A.M.; Khoja, S.S.; Josbeno, D.A.; Fitzgerald, G.K.; Toledo, F.G. Links between osteoarthritis and diabetes: Implications for management from a physical activity perspective. Clin. Geriatr. Med. 2015, 31, 67–87. [Google Scholar]
- Laiguillon, M.C.; Courties, A.; Houard, X.; Auclair, M.; Sautet, A.; Capeau, J.; Fève, B.; Berenbaum, F.; Sellam, J. Characterization of diabetic osteoarthritic cartilage and role of high glucose environment on chondrocyte activation: Toward pathophysiological delineation of diabetes mellitus-related osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1513–1522. [Google Scholar]
- Courties, A.; Berenbaum, F.; Sellam, J. The phenotypic approach to osteoarthritis: A look at metabolic syndrome-associated osteoarthritis. Jt. Bone Spine 2019, 86, 725–730. [Google Scholar]
- Akbari, M.; Hassan-Zadeh, V. IL-6 signalling pathways and the development of type 2 diabetes. Inflammopharmacology 2018, 26, 685–698. [Google Scholar]
- Spranger, J.; Kroke, A.; Möhlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes: Results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [PubMed]
- Rodrigues, K.F.; Pietrani, N.T.; Bosco, A.A.; Campos, F.M.; Sandrim, V.C.; Gomes, K.B. IL-6, TNF-α, and IL-10 levels/polymorphisms and their association with type 2 diabetes mellitus and obesity in Brazilian individuals. Arch. Endocrinol. Metab. 2017, 61, 438–446. [Google Scholar] [CrossRef]
- Shlopov, B.V.; Gumanovskaya, M.L.; Hasty, K.A. Autocrine regulation of collagenase 3 (matrix metalloproteinase 13) during osteoarthritis. Arthritis Rheumatol. 2000, 43, 195–205. [Google Scholar]
- Sun, G.; Ba, C.L.; Gao, R.; Liu, W.; Ji, Q. Association of IL-6, IL-8, MMP-13 gene polymorphisms with knee osteoarthritis susceptibility in the Chinese Han population. Biosci. Rep. 2019, 39, 2. [Google Scholar] [CrossRef] [PubMed]
- Leyh, M.; Seitz, A.; Dürselen, L.; Springorum, H.R.; Angele, P.; Ignatius, A.; Grifka, J.; Grässel, S. Osteoarthritic cartilage explants affect extracellular matrix production and composition in cocultured bone marrow-derived mesenchymal stem cells and articular chondrocytes. Stem Cell Res. Ther. 2014, 5, 77. [Google Scholar] [CrossRef]
- Miyaji, N.; Nishida, K.; Tanaka, T.; Araki, D.; Kanzaki, N.; Hoshino, Y.; Kuroda, R.; Matsushita, T. Inhibition of knee osteoarthritis progression in mice by administering SRT2014, an activator of silent information regulator 2 ortholog 1. Cartilage 2021, 13, 1366S. [Google Scholar] [CrossRef] [PubMed]
- Wiegertjes, R.; van de Loo, F.A.; Blaney Davidson, E.N. A roadmap to target interleukin-6 in osteoarthritis. Rheumatology 2020, 59, 2681–2694. [Google Scholar] [CrossRef]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a promising target for the treatment of osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef]
- Ryu, J.H.; Yang, S.; Shin, Y.; Rhee, J.; Chun, H.; Chun, J.S. Interleukin-6 plays an essential role in hypoxia-inducible factor 2alpha-induced experimental osteoarthritic cartilage destruction in mice. Arthritis Rheum. 2011, 63, 2732–2743. [Google Scholar] [CrossRef] [PubMed]
- Behloul, N.; Wu, G. Genistein: A promising therapeutic agent for obesity and diabetes treatment. Eur. J. Pharmacol. 2013, 698, 31–38. [Google Scholar]
- Michelin, R.M.; Al-Nakkash, L.; Broderick, T.L.; Plochocki, J.H. Genistein treatment increases bone mass in obese, hyperglycemic mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2016, 15, 63–70. [Google Scholar]
- Hellings, A.; Buchan, L.; Castro, M.; St. Aubin, C.R.; Fisher, A.L.; Al-Nakkash, L.; Broderick, T.L.; Plochocki, J.H. Bone Strength Is Improved with Genistein Treatment in Mice with Diet-Induced Obesity. Curr. Dev. Nutr. 2019, 3, 121. [Google Scholar]
- Li, W.; Cai, L.; Zhang, Y.; Cui, L.; Shen, G. Intra-Articular Resveratrol Injection Prevents Osteoarthritis Progression in a Mouse Model by Activating SIRT1 and Thereby Silencing HIF-2alpha. J. Orthop. Res. 2015, 33, 1061–1070. [Google Scholar] [CrossRef]
- Di Rosa, M.; Castrogiovanni, P.; Musumeci, G. The synovium theory: Can exercise prevent knee osteoarthritis? The role of “mechanokines”, a possible biological key. J. Func. Morphol. Kinesiol. 2019, 4, 11. [Google Scholar] [CrossRef]
- Lu, J.; Feng, X.; Zhang, H.; Wei, Y.; Yang, Y.; Tian, Y.; Bai, L. Maresin-1 suppresses IL-1β-induced MMP-13 secretion by activating the PI3K/AKT pathway and inhibiting the NF-κB pathway in synovioblasts of an osteoarthritis rat model with treadmill exercise. Connect. Tissue Res. 2021, 62, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Buchan, L.; St. Aubin, C.R.; Fisher, A.L.; Hellings, A.; Castro, M.; Al-Nakkash, L.; Broderick, T.L.; Plochocki, J.H. High-fat, high-sugar diet induces splenomegaly that is ameliorated with exercise and genistein treatment. BMC Res. Notes 2018, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- St Aubin, C.R.; Fisher, A.L.; Hernandez, J.A.; Broderick, T.L.; Al-Nakkash, L. Mitigation of MAFLD in high fat-high sucrose-fructose fed mice by a combination of genistein consumption and exercise training. Diabetes Metab. Syndr. Obes. Targets Ther. 2022, 1, 2157–2172. [Google Scholar] [CrossRef]
- Liao, Y.; Ren, Y.; Luo, X.; Mirando, A.J.; Long, J.T.; Leinroth, A.; Ji, R.R.; Hilton, M.J. Interleukin-6 signaling mediates cartilage degradation and pain in posttraumatic osteoarthritis in a sex-specific manner. Sci. Signal. 2022, 15, 7082. [Google Scholar] [CrossRef] [PubMed]
- Galien, R.; Garcia, T. Estrogen receptor impairs interleukin-6 expression by preventing protein binding on the NF-κB site. Nucleic Acids Res. 1997, 25, 2424–2429. [Google Scholar] [CrossRef]
- Zhang, E.; Zhang, H.; Liu, F.; Dong, C.; Yao, Y.; Yun, Z.; Jian, W.; Ma, B. Estrogen exerts anti-inflammatory effects by inhibiting NF-κB pathway through binding with estrogen receptor β on synovicytes of osteoarthritis. Chin. J. Cell. Mol. Immunol. 2016, 32, 1605–1609. [Google Scholar]
- Roman-Blas, J.A.; Castañeda, S.; Largo, R.; Herrero-Beaumont, G. Osteoarthritis associated with estrogen deficiency. Arthritis Res. Ther. 2009, 11, 1–4. [Google Scholar] [CrossRef]
- Wang, W.; Wang, L.; Xu, Z.; Yin, Y.; Su, J.; Niu, X.; Cao, X. Effects of estradiol on reduction of osteoarthritis in rabbits through effect on matrix metalloproteinase proteins. Iran. J. Basic Med. Sci. 2016, 19, 310. [Google Scholar]
- Li, H.; Wang, D.; Yuan, Y.; Min, J. New insights on the MMP-13 regulatory network in the pathogenesis of early osteoarthritis. Arthritis Res. Ther. 2017, 19, 248. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Han, L.; Wang, X.; Li, Y.; Zhu, Y.; Wang, J.; Xue, C. Sialoglycoprotein isolated from eggs of Carassius auratus promotes fracture healing in osteoporotic mice. J. Food Drug Anal. 2018, 26, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Madzuki, I.N.; Lau, S.F.; Mohamad Shalan, N.A.; Mohd Ishak, N.I.; Mohamed, S. Does cartilage ERα overexpression correlate with osteoarthritic chondrosenescence? Indications from Labisia pumila OA mitigation. J. Biosci. 2019, 44, 1. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, L. The protective activity of genistein against bone and cartilage diseases. Front. Pharmacol. 2022, 13, 1016981. [Google Scholar] [CrossRef]
- Yuan, J.; Ding, W.; Wu, N.; Jiang, S.; Li, W. Protective effect of genistein on condylar cartilage through downregulating NF-κB expression in experimentally created osteoarthritis rats. BioMed Res. Int. 2019, 30, 2019. [Google Scholar] [CrossRef]
- Zhang, H.; Ji, L.; Yang, Y.; Wei, Y.; Zhang, X.; Gang, Y.; Lu, J.; Bai, L. The therapeutic effects of treadmill exercise on osteoarthritis in rats by inhibiting the HDAC3/NF-KappaB pathway in vivo and in vitro. Front. Physiol. 2019, 10, 1060. [Google Scholar]
- Helge, J.W.; Stallknecht, B.; Pedersen, B.K.; Galbo, H.; Kiens, B.; Richter, E.A. The effect of graded exercise on IL-6 release and glucose uptake in human skeletal muscle. J. Physiol. 2003, 546, 299–305. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Fischer, C.P. Beneficial health effects of exercise—The role of IL-6 as a myokine. Trends Pharmacol. Sci. 2007, 28, 152–156. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Osteoarthritic Changes |
|---|---|
| Normal | Normal structure and staining |
| Mild | Loss of matrix staining with normal structure |
| Moderate | Loss of matrix staining, mild fibrillations, hypertrophic chondrocytes |
| Severe | Loss of matrix staining, deep fibrillations, hypertrophic chondrocytes, reduced cellularity in radial zone |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, J.; Al-Nakkash, L.; Broderick, T.L.; Castro, M.; Tobin, B.; Plochocki, J.H. Genistein Suppresses IL-6 and MMP-13 to Attenuate Osteoarthritis in Obese Diabetic Mice. Metabolites 2023, 13, 1014. https://doi.org/10.3390/metabo13091014
Lopez J, Al-Nakkash L, Broderick TL, Castro M, Tobin B, Plochocki JH. Genistein Suppresses IL-6 and MMP-13 to Attenuate Osteoarthritis in Obese Diabetic Mice. Metabolites. 2023; 13(9):1014. https://doi.org/10.3390/metabo13091014
Chicago/Turabian StyleLopez, Janelle, Layla Al-Nakkash, Tom L. Broderick, Monica Castro, Brielle Tobin, and Jeffrey H. Plochocki. 2023. "Genistein Suppresses IL-6 and MMP-13 to Attenuate Osteoarthritis in Obese Diabetic Mice" Metabolites 13, no. 9: 1014. https://doi.org/10.3390/metabo13091014
APA StyleLopez, J., Al-Nakkash, L., Broderick, T. L., Castro, M., Tobin, B., & Plochocki, J. H. (2023). Genistein Suppresses IL-6 and MMP-13 to Attenuate Osteoarthritis in Obese Diabetic Mice. Metabolites, 13(9), 1014. https://doi.org/10.3390/metabo13091014