
From the Sun to the Cell: Examining Obesity through the Lens of Vitamin D and Inflammation
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Adipocyte Dynamics in Obesity: Connections with Cellular Senescence
3.1.1. Senescence-Associated Secretory Phenotype (SASP)
3.1.2. Preadipocyte Changes in Obesity and Senescent AT
3.1.3. Neural and Vascular Dysfunction in Obesity and Senescent AT
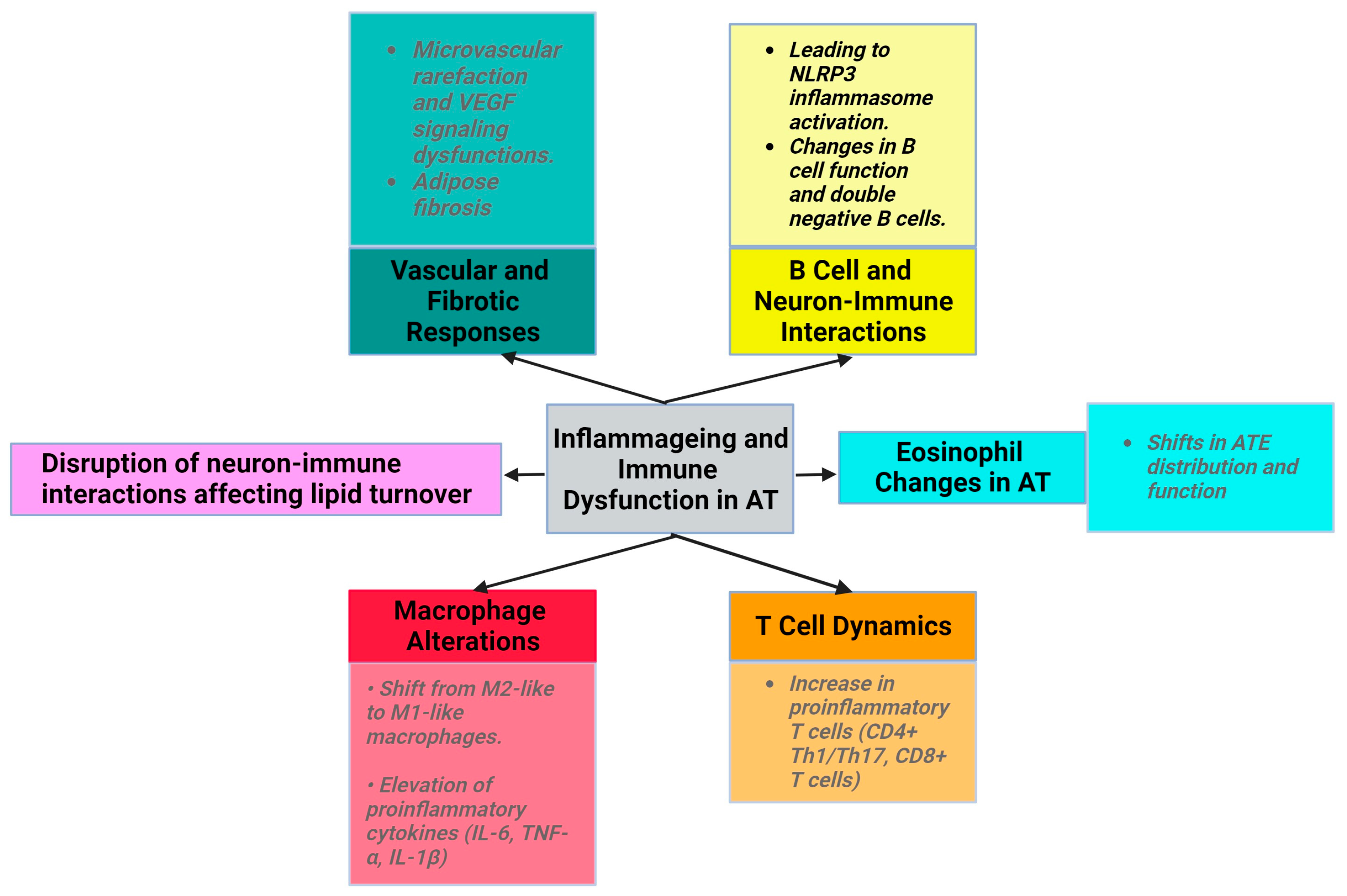
3.1.4. Low-Grade Inflammation in Obesity and Inflammageing
3.1.5. Extracellular Vesicles and Particles (EVPs)
3.2. Physiological Roles of Vitamin D: Bone Health, Immune Function, and Beyond
3.2.1. Vitamin D Deficiency in the Context of Obesity
3.2.2. The Role of Vitamin D in the Senescence Process
3.3. Exploring the Shared Characteristics of Obesity and Ageing with a Focus on Vitamin D
3.3.1. Toll-like Receptors (TLRs)
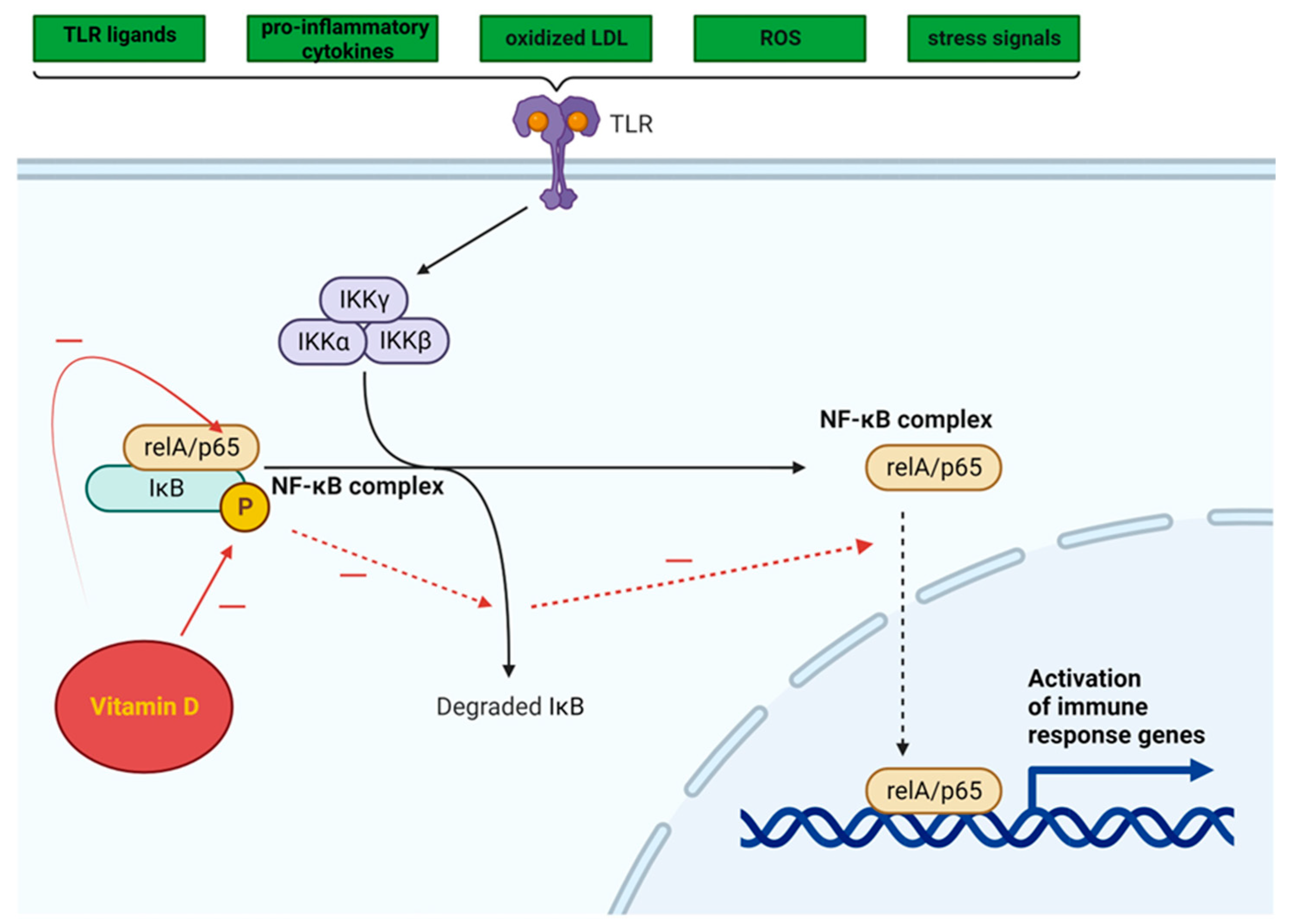
3.3.2. NF-κB Pathway
3.3.3. NLRP3 Inflammasome Activation
3.3.4. Immune Cell Modulation
The Anti-Inflammatory Action of Vitamin D in Cultured Cells
Inconsistent Results in Human Studies
Insights from Meta-Analyses
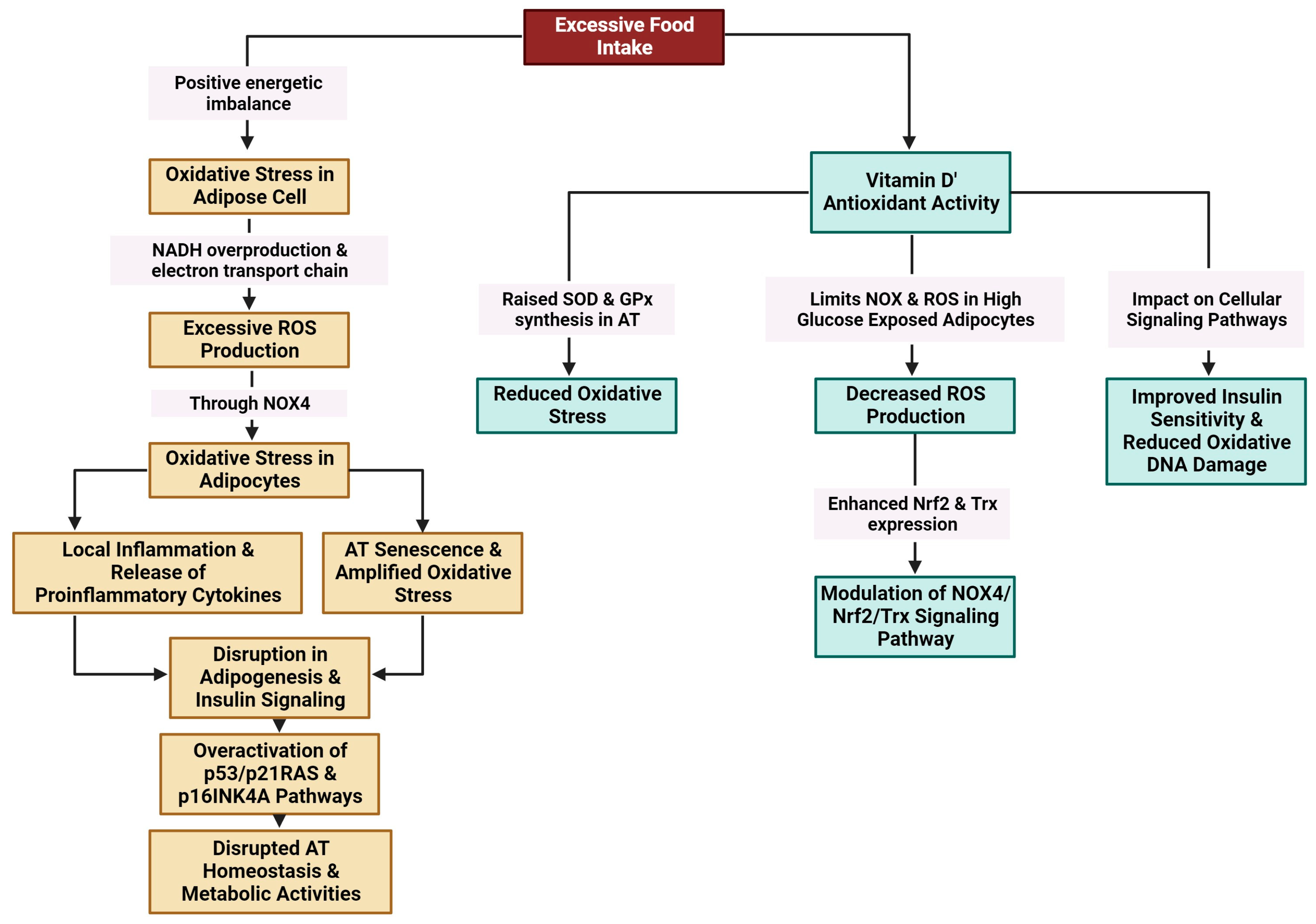
3.3.5. Oxidative Stress
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1,25(OH)2D3 | 1,25-Dihydroxyvitamin D3 |
| 25(OH)D | 25-Dihydroxyvitamin D |
| AMPK | AMP-activated protein kinase |
| AT | Adipose tissue |
| ATEs | Adipose tissue eosinophils |
| ATMs | Adipose tissue macrophages |
| BMI | Body mass index |
| C/EBP | CCAAT/enhancer-binding protein |
| CDKNlA | Cyclin-dependent kinase inhibitor-1 A |
| CPT1B | Carnitine palmitoyltransferase 1B |
| CRP | C-reactive protein |
| Cyp27b1 | Cytochrome P450, family 27, subfamily b, polypeptide 1 |
| DAG/IP3 | Diacylglycerol/inositol |
| ER | Endoplasmic reticulum |
| EVPs | Extracellular vesicles and particles |
| FATP4 | Fatty acid transport protein 4 |
| GATA-3 | Trans-acting T-cell-specific transcription factor |
| GPx | Glutathione peroxidase |
| HIF1α | Hypoxia-inducible factor 1-alpha |
| HOMA-IR | Homeostatic model assessment for insulin resistance |
| IFN | Interferon |
| IL | Interleukins |
| ILC2 | Group-2 innate lymphoid |
| IκB | Inhibitor of kappa B |
| JAK-STAT | Janus kinase-signal transducer and activator of transcription |
| LPSs | Lipopolysaccharides |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein-1 (CCL2, chemokine (C-C motif) ligand 2) |
| miRs | MicroRNAs |
| mRNA | Messenger RNA |
| MVECs | Microvascular endothelial cells |
| NAD | Nicotinamide adenine dinucleotide |
| NADH | Nicotinamide adenine dinucleotide (NAD) hydrogenase |
| NADPH | Nicotinamide adenine dinucleotide phosphate (NADP) hydrogenase |
| NF-κB | Nuclear factor kappa B subunit 1 |
| NLRP3 | NLR family pyrin domain containing 3 |
| NOX4 | NADPH oxidase 4 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| OPN | Osteopontin |
| p16 | p16-INK4a (cyclin-dependent kinase inhibitor 2A) |
| p21 | P21 (cyclin-dependent kinase inhibitor 1) |
| p42/44 (ERK1/2) | Extracellular signal-related kinase |
| p53 | Cellular tumour antigen p53 (tumour suppressor p53) |
| PAI-1 | Plasminogen activator inhibitor-1 |
| Pam3CSK4 | Pam3CysSerLys4 |
| PARP1 | Poly (ADP-ribose) polymerase 1 |
| PDGFRα+ | Platelet-derived growth factor receptor alpha |
| PFM | Percentage of fat mass |
| PI3K | Phosphoinositide 3-kinase |
| PKC | Protein kinase C |
| PLC | Phospholipase C |
| pp38 | Phosphoprotein 38 |
| PPAR γ | peroxisome proliferator-activated receptor γ |
| PPARA | peroxisome proliferator-activated receptor alpha |
| RCTs | Randomized controlled trials |
| ROS | Reactive oxygen species |
| RXR | Retinoid X receptor |
| SA β-gal | Senescence-associated β-galactosidase |
| SASP | Senescence-associated secretory phenotype |
| SAT | Subcutaneous adipose tissue |
| scWAT | Subcutaneous white adipose tissue |
| SIRT1 | Sirtuin 1 gene |
| SOD | Superoxide dismutase |
| SREBP1c | Sterol regulatory element-binding protein 1 |
| STAT 6 | Factor signal transducer and activator of transcription 6 |
| SVCs | Stromal vascular cells |
| SVF | Stromal vascular fraction |
| T2DM | Type 2 diabetes mellitus |
| TG/HDL | Triglyceride (TG) to high-density lipoprotein (HDL) |
| TGFβ | Transforming growth factor beta |
| TLR | Toll-like receptor |
| TNFα | Tumour necrosis factor α |
| Tregs | Regulatory T cells |
| Trf1 | Telomere repeat-binding factor 1 |
| Trx | Thioredoxin |
| VAT | Visceral adipose tissue |
| VDR | Vitamin D receptor |
| VDRE | Vitamin D response element |
| VEGF | Vascular endothelial growth factor |
| vWAT | Visceral white adipose tissue |
| WAT | White adipose tissue |
| WAT-MSCs | White-adipose-tissue-resident multipotent stromal cells |
References
- Frasca, D.; Blomberg, B.B. Adipose Tissue, Immune Aging, and Cellular Senescence. Semin. Immunopathol. 2020, 42, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; He, B. Adipose Tissue Aging: An Update on Mechanisms and Therapeutic Strategies. Metabolism 2023, 138, 155328. [Google Scholar] [CrossRef]
- Qian, S.; Tang, Y.; Tang, Q.-Q. Adipose Tissue Plasticity and the Pleiotropic Roles of BMP Signaling. J. Biol. Chem. 2021, 296, 100678. [Google Scholar] [CrossRef] [PubMed]
- Smith, U.; Li, Q.; Rydén, M.; Spalding, K.L. Cellular Senescence and Its Role in White Adipose Tissue. Int. J. Obes. 2021, 45, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Rana, B.M.J.; Jou, E.; Barlow, J.L.; Rodriguez-Rodriguez, N.; Walker, J.A.; Knox, C.; Jolin, H.E.; Hardman, C.S.; Sivasubramaniam, M.; Szeto, A.; et al. A Stromal Cell Niche Sustains ILC2-Mediated Type-2 Conditioning in Adipose Tissue. J. Exp. Med. 2019, 216, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Molofsky, A.B.; Liang, H.-E.; Ricardo-Gonzalez, R.R.; Jouihan, H.A.; Bando, J.K.; Chawla, A.; Locksley, R.M. Eosinophils Sustain Adipose Alternatively Activated Macrophages Associated with Glucose Homeostasis. Science 2011, 332, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chan, Y.T.; Revelo, X.S.; Winer, D.A. The Immune Landscape of Visceral Adipose Tissue During Obesity and Aging. Front. Endocrinol. 2020, 11, 267. [Google Scholar] [CrossRef] [PubMed]
- Moslemi, E.; Musazadeh, V.; Kavyani, Z.; Naghsh, N.; Shoura, S.M.S.; Dehghan, P. Efficacy of Vitamin D Supplementation as an Adjunct Therapy for Improving Inflammatory and Oxidative Stress Biomarkers: An Umbrella Meta-Analysis. Pharmacol. Res. 2022, 186, 106484. [Google Scholar] [CrossRef]
- Chang, E. Effects of Vitamin D Supplementation on Adipose Tissue Inflammation and NF-κB/AMPK Activation in Obese Mice Fed a High-Fat Diet. Int. J. Mol. Sci. 2022, 23, 10915. [Google Scholar] [CrossRef]
- Martinelli, R.P.; Rayego-Mateos, S.; Alique, M.; Márquez-Expósito, L.; Tejedor-Santamaria, L.; Ortiz, A.; González-Parra, E.; Ruiz-Ortega, M. Vitamin D, Cellular Senescence and Chronic Kidney Diseases: What Is Missing in the Equation? Nutrients 2023, 15, 1349. [Google Scholar] [CrossRef]
- Narasimhan, A.; Flores, R.R.; Camell, C.D.; Bernlohr, D.A.; Robbins, P.D.; Niedernhofer, L.J. Cellular Senescence in Obesity and Associated Complications: A New Therapeutic Target. Curr. Diab. Rep. 2022, 22, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Nerstedt, A.; Smith, U. The Impact of Cellular Senescence in Human Adipose Tissue. J. Cell Commun. Signal. 2023, 17, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Lazar, M.A. Peroxisome Proliferator Activated Receptor Gamma, CCAAT/Enhancer-Binding Protein Alpha, and Cell Cycle Status Regulate the Commitment to Adipocyte Differentiation. J. Biol. Chem. 1997, 272, 21473–21478. [Google Scholar] [CrossRef]
- Madsen, M.S.; Siersbæk, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome Proliferator-Activated Receptor γ and C/EBPα Synergistically Activate Key Metabolic Adipocyte Genes by Assisted Loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.-Y.; Zhang, H.; Tan, P.-C.; Zhou, S.-B.; Li, Q.-F. Adipose Tissue Aging: Mechanisms and Therapeutic Implications. Cell Death Dis. 2022, 13, 300. [Google Scholar] [CrossRef] [PubMed]
- Tabula Muris Consortium. A Single-Cell Transcriptomic Atlas Characterizes Ageing Tissues in the Mouse. Nature 2020, 583, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Varghese, M.; Song, J.; Singer, K. Age and Sex: Impact on Adipose Tissue Metabolism and Inflammation. Mech. Ageing Dev. 2021, 199, 111563. [Google Scholar] [CrossRef]
- Cartwright, M.J.; Schlauch, K.; Lenburg, M.E.; Tchkonia, T.; Pirtskhalava, T.; Cartwright, A.; Thomou, T.; Kirkland, J.L. Aging, Depot Origin, and Preadipocyte Gene Expression. J. Gerontol. A. Biol. Sci. Med. Sci. 2010, 65, 242–251. [Google Scholar] [CrossRef]
- Lee, G. Cellular Senescence: The Villain of Metabolic Disease? Mol. Cells 2022, 45, 531–533. [Google Scholar] [CrossRef]
- Lee, G.; Kim, Y.Y.; Jang, H.; Han, J.S.; Nahmgoong, H.; Park, Y.J.; Han, S.M.; Cho, C.; Lim, S.; Noh, J.-R.; et al. SREBP1c-PARP1 Axis Tunes Anti-Senescence Activity of Adipocytes and Ameliorates Metabolic Imbalance in Obesity. Cell Metab. 2022, 34, 702–718.e5. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Achari, A.E.; Jain, S.K. Vitamin D Supplementation Inhibits Oxidative Stress and Upregulate SIRT1/AMPK/GLUT4 Cascade in High Glucose-Treated 3T3L1 Adipocytes and in Adipose Tissue of High Fat Diet-Fed Diabetic Mice. Arch. Biochem. Biophys. 2017, 615, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.E.; Ricotti, R.; Roccio, M.; Moia, S.; Bellone, S.; Prodam, F.; Bona, G. Pediatric Obesity and Vitamin D Deficiency: A Proteomic Approach Identifies Multimeric Adiponectin as a Key Link between These Conditions. PLoS ONE 2014, 9, e83685. [Google Scholar] [CrossRef] [PubMed]
- Tchkonia, T.; Morbeck, D.E.; Von Zglinicki, T.; Van Deursen, J.; Lustgarten, J.; Scrable, H.; Khosla, S.; Jensen, M.D.; Kirkland, J.L. Fat Tissue, Aging, and Cellular Senescence. Aging Cell 2010, 9, 667–684. [Google Scholar] [CrossRef] [PubMed]
- Matacchione, G.; Perugini, J.; Di Mercurio, E.; Sabbatinelli, J.; Prattichizzo, F.; Senzacqua, M.; Storci, G.; Dani, C.; Lezoche, G.; Guerrieri, M.; et al. Senescent Macrophages in the Human Adipose Tissue as a Source of Inflammaging. GeroScience 2022, 44, 1941–1960. [Google Scholar] [CrossRef] [PubMed]
- Schosserer, M.; Grillari, J.; Wolfrum, C.; Scheideler, M. Age-Induced Changes in White, Brite, and Brown Adipose Depots: A Mini-Review. Gerontology 2018, 64, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Palmer, A.K.; Ding, H.; Weivoda, M.M.; Pirtskhalava, T.; White, T.A.; Sepe, A.; Johnson, K.O.; Stout, M.B.; Giorgadze, N.; et al. Targeting Senescent Cells Enhances Adipogenesis and Metabolic Function in Old Age. eLife 2015, 4, e12997. [Google Scholar] [CrossRef]
- Schafer, M.J.; Zhang, X.; Kumar, A.; Atkinson, E.J.; Zhu, Y.; Jachim, S.; Mazula, D.L.; Brown, A.K.; Berning, M.; Aversa, Z.; et al. The Senescence-Associated Secretome as an Indicator of Age and Medical Risk. JCI Insight 2020, 5, e133668. [Google Scholar] [CrossRef]
- Zaragosi, L.-E.; Wdziekonski, B.; Villageois, P.; Keophiphath, M.; Maumus, M.; Tchkonia, T.; Bourlier, V.; Mohsen-Kanson, T.; Ladoux, A.; Elabd, C.; et al. Activin A Plays a Critical Role in Proliferation and Differentiation of Human Adipose Progenitors. Diabetes 2010, 59, 2513–2521. [Google Scholar] [CrossRef]
- Gao, H.; Arner, P.; Beauchef, G.; Guéré, C.; Vie, K.; Dahlman, I.; Mejhert, N.; Rydén, M. Age-Induced Reduction in Human Lipolysis: A Potential Role for Adipocyte Noradrenaline Degradation. Cell Metab. 2020, 32, 1–3. [Google Scholar] [CrossRef]
- Camell, C.D.; Sander, J.; Spadaro, O.; Lee, A.; Nguyen, K.Y.; Wing, A.; Goldberg, E.L.; Youm, Y.-H.; Brown, C.W.; Elsworth, J.; et al. Inflammasome-Driven Catecholamine Catabolism in Macrophages Blunts Lipolysis during Ageing. Nature 2017, 550, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, M.; Kumar, S.; Sharife, H.; Volinsky, E.; Gileles-Hillel, A.; Licht, T.; Permyakova, A.; Hinden, L.; Azar, S.; Friedmann, Y.; et al. Counteracting Age-Related VEGF Signaling Insufficiency Promotes Healthy Aging and Extends Life Span. Science 2021, 373, eabc8479. [Google Scholar] [CrossRef] [PubMed]
- Sawaki, D.; Czibik, G.; Pini, M.; Ternacle, J.; Suffee, N.; Mercedes, R.; Marcelin, G.; Surenaud, M.; Marcos, E.; Gual, P.; et al. Visceral Adipose Tissue Drives Cardiac Aging Through Modulation of Fibroblast Senescence by Osteopontin Production. Circulation 2018, 138, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Diaz, A.; Romero, M.; Thaller, S.; Blomberg, B.B. Metabolic Requirements of Human Pro-Inflammatory B Cells in Aging and Obesity. PLoS ONE 2019, 14, e0219545. [Google Scholar] [CrossRef] [PubMed]
- Villaret, A.; Galitzky, J.; Decaunes, P.; Estève, D.; Marques, M.-A.; Sengenès, C.; Chiotasso, P.; Tchkonia, T.; Lafontan, M.; Kirkland, J.L.; et al. Adipose Tissue Endothelial Cells from Obese Human Subjects: Differences among Depots in Angiogenic, Metabolic, and Inflammatory Gene Expression and Cellular Senescence. Diabetes 2010, 59, 2755–2763. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Diaz, A.; Romero, M.; Vazquez, T.; Blomberg, B.B. Obesity Induces Pro-Inflammatory B Cells and Impairs B Cell Function in Old Mice. Mech. Ageing Dev. 2017, 162, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Dahlquist, K.J.V.; Camell, C.D. Aging Leukocytes and the Inflammatory Microenvironment of the Adipose Tissue. Diabetes 2022, 71, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Romero, M.; Diaz, A.; Garcia, D.; Thaller, S.; Blomberg, B.B. B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity. Int. J. Mol. Sci. 2021, 22, 1839. [Google Scholar] [CrossRef]
- Tchkonia, T.; Kirkland, J.L. Aging, Cell Senescence, and Chronic Disease: Emerging Therapeutic Strategies. JAMA 2018, 320, 1319–1320. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P.-Y. The NLRP3 Inflammasome: Molecular Activation and Regulation to Therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Blevins, H.M.; Xu, Y.; Biby, S.; Zhang, S. The NLRP3 Inflammasome Pathway: A Review of Mechanisms and Inhibitors for the Treatment of Inflammatory Diseases. Front. Aging Neurosci. 2022, 14, 879021. [Google Scholar] [CrossRef] [PubMed]
- Summers, S.A. Ceramides in Insulin Resistance and Lipotoxicity. Prog. Lipid Res. 2006, 45, 42–72. [Google Scholar] [CrossRef] [PubMed]
- Brigger, D.; Riether, C.; van Brummelen, R.; Mosher, K.I.; Shiu, A.; Ding, Z.; Zbären, N.; Gasser, P.; Guntern, P.; Yousef, H.; et al. Eosinophils Regulate Adipose Tissue Inflammation and Sustain Physical and Immunological Fitness in Old Age. Nat. Metab. 2020, 2, 688–702. [Google Scholar] [CrossRef] [PubMed]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The Ageing Systemic Milieu Negatively Regulates Neurogenesis and Cognitive Function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, E.L.; Shchukina, I.; Youm, Y.-H.; Ryu, S.; Tsusaka, T.; Young, K.C.; Camell, C.D.; Dlugos, T.; Artyomov, M.N.; Dixit, V.D. IL-33 Causes Thermogenic Failure in Aging by Expanding Dysfunctional Adipose ILC2. Cell Metab. 2021, 33, 2277–2287.e5. [Google Scholar] [CrossRef] [PubMed]
- Cipolletta, D.; Cohen, P.; Spiegelman, B.M.; Benoist, C.; Mathis, D. Appearance and Disappearance of the mRNA Signature Characteristic of Treg Cells in Visceral Adipose Tissue: Age, Diet, and PPARγ Effects. Proc. Natl. Acad. Sci. USA 2015, 112, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-Q.; Zhao, W.-H. The IL-33/ST2 Axis Is Specifically Required for Development of Adipose Tissue-Resident Regulatory T Cells. Cell. Mol. Immunol. 2015, 12, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Lynch, L.; Nowak, M.; Varghese, B.; Clark, J.; Hogan, A.E.; Toxavidis, V.; Balk, S.P.; O’Shea, D.; O’Farrelly, C.; Exley, M.A. Adipose Tissue Invariant NKT Cells Protect against Diet-Induced Obesity and Metabolic Disorder through Regulatory Cytokine Production. Immunity 2012, 37, 574–587. [Google Scholar] [CrossRef]
- Adlowitz, D.G.; Barnard, J.; Biear, J.N.; Cistrone, C.; Owen, T.; Wang, W.; Palanichamy, A.; Ezealah, E.; Campbell, D.; Wei, C.; et al. Expansion of Activated Peripheral Blood Memory B Cells in Rheumatoid Arthritis, Impact of B Cell Depletion Therapy, and Biomarkers of Response. PLoS ONE 2015, 10, e0128269. [Google Scholar] [CrossRef]
- Siqueira, I.R.; de Souza Rodrigues, A.; Flores, M.S.; Vieira Cunha, E.L.; Goldberg, M.; Harmon, B.; Batabyal, R.; Freishtat, R.J.; Cechinel, L.R. Circulating Extracellular Vesicles and Particles Derived From Adipocytes: The Potential Role in Spreading MicroRNAs Associated With Cellular Senescence. Front. Aging 2022, 3, 867100. [Google Scholar] [CrossRef]
- Matz, A.J.; Qu, L.; Karlinsey, K.; Vella, A.T.; Zhou, B. Capturing the Multifaceted Function of Adipose Tissue Macrophages. Front. Immunol. 2023, 14, 1148188. [Google Scholar] [CrossRef] [PubMed]
- Rakib, A.; Kiran, S.; Mandal, M.; Singh, U.P. MicroRNAs: A Crossroad That Connects Obesity to Immunity and Aging. Immun. Ageing 2022, 19, 64. [Google Scholar] [CrossRef] [PubMed]
- Hubal, M.J.; Nadler, E.P.; Ferrante, S.C.; Barberio, M.D.; Suh, J.-H.; Wang, J.; Dohm, G.L.; Pories, W.J.; Mietus-Snyder, M.; Freishtat, R.J. Circulating adipocyte-derived exosomal micrornas associated with decreased insulin resistance after gastric bypass. Obesity 2017, 25, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Carmeliet, G.; Lieben, L.; Watanabe, M.; Perino, A.; Auwerx, J.; Schoonjans, K.; Verstuyf, A. Vitamin D and Energy Homeostasis: Of Mice and Men. Nat. Rev. Endocrinol. 2014, 10, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Narvaez, C.J.; Matthews, D.; Broun, E.; Chan, M.; Welsh, J. Lean Phenotype and Resistance to Diet-Induced Obesity in Vitamin D Receptor Knockout Mice Correlates with Induction of Uncoupling Protein-1 in White Adipose Tissue. Endocrinology 2009, 150, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.E.; Kong, J.; Zhang, W.; Szeto, F.L.; Ye, H.; Deb, D.K.; Brady, M.J.; Li, Y.C. Targeted Expression of Human Vitamin D Receptor in Adipocytes Decreases Energy Expenditure and Induces Obesity in Mice. J. Biol. Chem. 2011, 286, 33804–33810. [Google Scholar] [CrossRef] [PubMed]
- Marcotorchino, J.; Tourniaire, F.; Astier, J.; Karkeni, E.; Canault, M.; Amiot, M.-J.; Bendahan, D.; Bernard, M.; Martin, J.-C.; Giannesini, B.; et al. Vitamin D Protects against Diet-Induced Obesity by Enhancing Fatty Acid Oxidation. J. Nutr. Biochem. 2014, 25, 1077–1083. [Google Scholar] [CrossRef]
- Kang, E.-J.; Lee, J.-E.; An, S.-M.; Lee, J.H.; Kwon, H.S.; Kim, B.C.; Kim, S.J.; Kim, J.M.; Hwang, D.Y.; Jung, Y.-J.; et al. The Effects of Vitamin D3 on Lipogenesis in the Liver and Adipose Tissue of Pregnant Rats. Int. J. Mol. Med. 2015, 36, 1151–1158. [Google Scholar] [CrossRef]
- Izzo, M.; Carrizzo, A.; Izzo, C.; Cappello, E.; Cecere, D.; Ciccarelli, M.; Iannece, P.; Damato, A.; Vecchione, C.; Pompeo, F. Vitamin D: Not Just Bone Metabolism but a Key Player in Cardiovascular Diseases. Life 2021, 11, 452. [Google Scholar] [CrossRef]
- Nemeth, Z.; Patonai, A.; Simon-Szabó, L.; Takács, I. Interplay of Vitamin D and SIRT1 in Tissue-Specific Metabolism-Potential Roles in Prevention and Treatment of Non-Communicable Diseases Including Cancer. Int. J. Mol. Sci. 2023, 24, 6154. [Google Scholar] [CrossRef]
- Szymczak-Pajor, I.; Miazek, K.; Selmi, A.; Balcerczyk, A.; Śliwińska, A. The Action of Vitamin D in Adipose Tissue: Is There the Link between Vitamin D Deficiency and Adipose Tissue-Related Metabolic Disorders? Int. J. Mol. Sci. 2022, 23, 956–N.PAG. [Google Scholar] [CrossRef] [PubMed]
- Pludowski, P.; Takacs, I.; Boyanov, M.; Belaya, Z.; Diaconu, C.C.; Mokhort, T.; Zherdova, N.; Rasa, I.; Payer, J.; Pilz, S. Clinical Practice in the Prevention, Diagnosis and Treatment of Vitamin D Deficiency: A Central and Eastern European Expert Consensus Statement. Nutrients 2022, 14, 1483. [Google Scholar] [CrossRef] [PubMed]
- Vieth, R.; Holick, M.F. Chapter 57B—The IOM—Endocrine Society Controversy on Recommended Vitamin D Targets: In Support of the Endocrine Society Position. In Vitamin D, 4th ed.; Feldman, D., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1091–1107. ISBN 978-0-12-809965-0. [Google Scholar]
- Holick, M.F. Vitamin D Deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Evaluation, Treatment, and Prevention of Vitamin D Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Han, S.N. The Role of Vitamin D in Adipose Tissue Biology: Adipocyte Differentiation, Energy Metabolism, and Inflammation. J. Lipid Atheroscler. 2021, 10, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Carrelli, A.; Bucovsky, M.; Horst, R.; Cremers, S.; Zhang, C.; Bessler, M.; Schrope, B.; Evanko, J.; Blanco, J.; Silverberg, S.J.; et al. Vitamin D Storage in Adipose Tissue of Obese and Normal Weight Women. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2017, 32, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Drincic, A.T.; Armas, L.A.G.; Van Diest, E.E.; Heaney, R.P. Volumetric Dilution, Rather than Sequestration Best Explains the Low Vitamin D Status of Obesity. Obesity 2012, 20, 1444–1448. [Google Scholar] [CrossRef]
- Pannu, P.K.; Zhao, Y.; Soares, M.J. Reductions in Body Weight and Percent Fat Mass Increase the Vitamin D Status of Obese Subjects: A Systematic Review and Metaregression Analysis. Nutr. Res. 2016, 36, 201–213. [Google Scholar] [CrossRef]
- Mallard, S.R.; Howe, A.S.; Houghton, L.A. Vitamin D Status and Weight Loss: A Systematic Review and Meta-Analysis of Randomized and Nonrandomized Controlled Weight-Loss Trials. Am. J. Clin. Nutr. 2016, 104, 1151–1159. [Google Scholar] [CrossRef]
- Renzaho, A.M.N.; Halliday, J.A.; Nowson, C. Vitamin D, Obesity, and Obesity-Related Chronic Disease among Ethnic Minorities: A Systematic Review. Nutrition 2011, 27, 868–879. [Google Scholar] [CrossRef]
- Saneei, P.; Salehi-Abargouei, A.; Esmaillzadeh, A. Serum 25-Hydroxy Vitamin D Levels in Relation to Body Mass Index: A Systematic Review and Meta-Analysis. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2013, 14, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Vimaleswaran, K.S.; Berry, D.J.; Lu, C.; Tikkanen, E.; Pilz, S.; Hiraki, L.T.; Cooper, J.D.; Dastani, Z.; Li, R.; Houston, D.K.; et al. Causal Relationship between Obesity and Vitamin D Status: Bi-Directional Mendelian Randomization Analysis of Multiple Cohorts. PLoS Med. 2013, 10, e1001383. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Santos, M.; Costa, P.R.F.; Assis, A.M.O.; Santos, C.a.S.T.; Santos, D.B. Obesity and Vitamin D Deficiency: A Systematic Review and Meta-Analysis. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2015, 16, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Shantavasinkul, P.C.; Phanachet, P.; Puchaiwattananon, O.; Chailurkit, L.; Lepananon, T.; Chanprasertyotin, S.; Ongphiphadhanakul, B.; Warodomwichit, D. Vitamin D Status Is a Determinant of Skeletal Muscle Mass in Obesity According to Body Fat Percentage. Nutrition 2015, 31, 801–806. [Google Scholar] [CrossRef]
- Golzarand, M.; Hollis, B.W.; Mirmiran, P.; Wagner, C.L.; Shab-Bidar, S. Vitamin D Supplementation and Body Fat Mass: A Systematic Review and Meta-Analysis. Eur. J. Clin. Nutr. 2018, 72, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Zittermann, A.; Gummert, J.F.; Börgermann, J. Vitamin D Deficiency and Mortality. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.B.; Valdes, A.M.; Gardner, J.P.; Paximadas, D.; Kimura, M.; Nessa, A.; Lu, X.; Surdulescu, G.L.; Swaminathan, R.; Spector, T.D.; et al. Higher Serum Vitamin D Concentrations Are Associated with Longer Leukocyte Telomere Length in Women. Am. J. Clin. Nutr. 2007, 86, 1420–1425. [Google Scholar] [CrossRef]
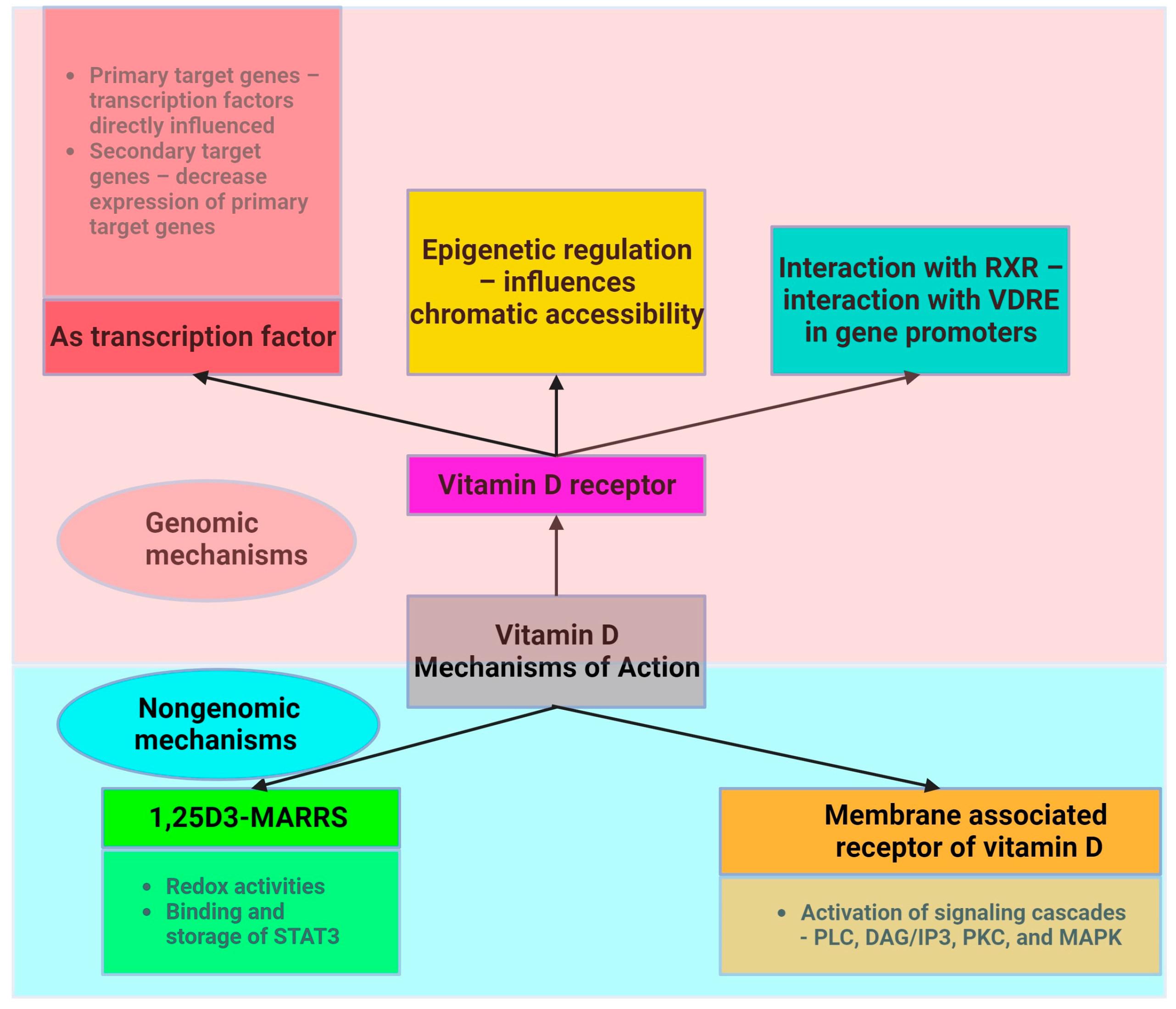
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.-C.; Jurutka, P.W. Molecular Mechanisms of Vitamin D Action. Calcif. Tissue Int. 2013, 92, 77–98. [Google Scholar] [CrossRef]
- Keisala, T.; Minasyan, A.; Lou, Y.-R.; Zou, J.; Kalueff, A.V.; Pyykkö, I.; Tuohimaa, P. Premature Aging in Vitamin D Receptor Mutant Mice. J. Steroid Biochem. Mol. Biol. 2009, 115, 91–97. [Google Scholar] [CrossRef]
- Haussler, M.R.; Haussler, C.A.; Jurutka, P.W. Chapter Ten—Genomically Anchored Vitamin D Receptor Mediates an Abundance of Bioprotective Actions Elicited by Its 1,25-Dihydroxyvitamin D Hormonal Ligand. In Vitamins and Hormones; Litwack, G., Ed.; Hormone Receptors: Structures and Functions; Academic Press: Cambridge, MA, USA, 2023; Volume 123, pp. 313–383. [Google Scholar]
- Chen, L.; Yang, R.; Qiao, W.; Zhang, W.; Chen, J.; Mao, L.; Goltzman, D.; Miao, D. 1,25-Dihydroxyvitamin D Exerts an Antiaging Role by Activation of Nrf2-antioxidant Signaling and Inactivation of P16/P53-senescence Signaling. Aging Cell 2019, 18, 12951. [Google Scholar] [CrossRef]
- Chen, G.Y.; Nuñez, G. Sterile Inflammation: Sensing and Reacting to Damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 Trafficking and Its Influence on LPS-Induced pro-Inflammatory Signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.H.; Slaby, E.M.; Stephanie, G.; Yu, C.; Watts, D.M.; Liu, H.; Szeto, G.L. Lipid-Mediated Insertion of Toll-Like Receptor (TLR) Ligands for Facile Immune Cell Engineering. Front. Immunol. 2020, 11, 560. [Google Scholar] [CrossRef] [PubMed]
- Rogero, M.M.; Calder, P.C. Obesity, Inflammation, Toll-Like Receptor 4 and Fatty Acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Jialal, I.; Kaur, H.; Devaraj, S. Toll-like Receptor Status in Obesity and Metabolic Syndrome: A Translational Perspective. J. Clin. Endocrinol. Metab. 2014, 99, 39–48. [Google Scholar] [CrossRef]
- Youssef-Elabd, E.M.; McGee, K.C.; Tripathi, G.; Aldaghri, N.; Abdalla, M.S.; Sharada, H.M.; Ashour, E.; Amin, A.I.; Ceriello, A.; O’Hare, J.P.; et al. Acute and Chronic Saturated Fatty Acid Treatment as a Key Instigator of the TLR-Mediated Inflammatory Response in Human Adipose Tissue, in Vitro. J. Nutr. Biochem. 2012, 23, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Vitseva, O.I.; Tanriverdi, K.; Tchkonia, T.T.; Kirkland, J.L.; McDonnell, M.E.; Apovian, C.M.; Freedman, J.; Gokce, N. Inducible Toll-like Receptor and NF-κB Regulatory Pathway Expression in Human Adipose Tissue. Obesity 2008, 16, 932–937. [Google Scholar] [CrossRef]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Salvador, J.; Frühbeck, G. Increased Tenascin C and Toll-like Receptor 4 Levels in Visceral Adipose Tissue as a Link between Inflammation and Extracellular Matrix Remodeling in Obesity. J. Clin. Endocrinol. Metab. 2012, 97, E1880–E1889. [Google Scholar] [CrossRef]
- Hari, P.; Millar, F.R.; Tarrats, N.; Birch, J.; Quintanilla, A.; Rink, C.J.; Fernández-Duran, I.; Muir, M.; Finch, A.J.; Brunton, V.G.; et al. The Innate Immune Sensor Toll-like Receptor 2 Controls the Senescence-Associated Secretory Phenotype. Sci. Adv. 2019, 5, eaaw0254. [Google Scholar] [CrossRef]
- Olivieri, F.; Rippo, M.R.; Prattichizzo, F.; Babini, L.; Graciotti, L.; Recchioni, R.; Procopio, A.D. Toll like Receptor Signaling in “Inflammaging”: microRNA as New Players. Immun. Ageing 2013, 10, 11. [Google Scholar] [CrossRef]
- Khalil, R.; Diab-Assaf, M.; Lemaitre, J.-M. Emerging Therapeutic Approaches to Target the Dark Side of Senescent Cells: New Hopes to Treat Aging as a Disease and to Delay Age-Related Pathologies. Cells 2023, 12, 915. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, S.; Thiemermann, C. Role of Metabolic Endotoxemia in Systemic Inflammation and Potential Interventions. Front. Immunol. 2021, 11, 594150. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, K.; Wessner, B.; Laggner, U.; Ploder, M.; Tamandl, D.; Friedl, J.; Zügel, U.; Steinmeyer, A.; Pollak, A.; Roth, E.; et al. Vitamin D3 Down-Regulates Monocyte TLR Expression and Triggers Hyporesponsiveness to Pathogen-Associated Molecular Patterns. Eur. J. Immunol. 2006, 36, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, I.; Pawliczak, R. The Active Metabolite of Vitamin D3 as a Potential Immunomodulator. Scand. J. Immunol. 2016, 83, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Ojaimi, S.; Skinner, N.A.; Strauss, B.J.; Sundararajan, V.; Woolley, I.; Visvanathan, K. Vitamin D Deficiency Impacts on Expression of Toll-like Receptor-2 and Cytokine Profile: A Pilot Study. J. Transl. Med. 2013, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M.; Malaguarnera, G.; De Gregorio, C.; Palumbo, M.; Nunnari, G.; Malaguarnera, L. Immuno-Modulatory Effects of Vitamin D3 in Human Monocyte and Macrophages. Cell. Immunol. 2012, 280, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Dickie, L.J.; Church, L.D.; Coulthard, L.R.; Mathews, R.J.; Emery, P.; McDermott, M.F. Vitamin D3 Down-Regulates Intracellular Toll-like Receptor 9 Expression and Toll-like Receptor 9-Induced IL-6 Production in Human Monocytes. Rheumatology 2010, 49, 1466–1471. [Google Scholar] [CrossRef]
- Kolieb, E.; Maher, S.A.; Shalaby, M.N.; Alsuhaibani, A.M.; Alharthi, A.; Hassan, W.A.; El-Sayed, K. Vitamin D and Swimming Exercise Prevent Obesity in Rats under a High-Fat Diet via Targeting FATP4 and TLR4 in the Liver and Adipose Tissue. Int. J. Environ. Res. Public. Health 2022, 19, 13740. [Google Scholar] [CrossRef]
- de Gregorio, E.; Colell, A.; Morales, A.; Marí, M. Relevance of SIRT1-NF-κB Axis as Therapeutic Target to Ameliorate Inflammation in Liver Disease. Int. J. Mol. Sci. 2020, 21, 3858. [Google Scholar] [CrossRef]
- Wang, R.; Yu, Z.; Sunchu, B.; Shoaf, J.; Dang, I.; Zhao, S.; Caples, K.; Bradley, L.; Beaver, L.M.; Ho, E.; et al. Rapamycin Inhibits the Secretory Phenotype of Senescent Cells by a Nrf2-Independent Mechanism. Aging Cell 2017, 16, 564–574. [Google Scholar] [CrossRef]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic Crosstalk between NF-κB and SIRT1 in the Regulation of Inflammation and Metabolic Disorders. Cell. Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Della Nera, G.; Sabatino, L.; Gaggini, M.; Gorini, F.; Vassalle, C. Vitamin D Determinants, Status, and Antioxidant/Anti-Inflammatory-Related Effects in Cardiovascular Risk and Disease: Not the Last Word in the Controversy. Antioxidants 2023, 12, 948. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, L.; Xu, H.-J.; Li, Y.; Hu, C.-M.; Yang, J.-Y.; Sun, M.-Y. The Anti-Inflammatory Effects of Vitamin D in Tumorigenesis. Int. J. Mol. Sci. 2018, 19, 2736. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Kim, T.Y.; Yoo, J.S.; Seo, Y.; Pae, M.; Han, S.N. Effects of 1,25-Dihydroxyvitamin D3 on the Inflammatory Responses of Stromal Vascular Cells and Adipocytes from Lean and Obese Mice. Nutrients 2020, 12, 364. [Google Scholar] [CrossRef] [PubMed]
- Marcotorchino, J.; Gouranton, E.; Romier, B.; Tourniaire, F.; Astier, J.; Malezet, C.; Amiot, M.-J.; Landrier, J.-F. Vitamin D Reduces the Inflammatory Response and Restores Glucose Uptake in Adipocytes. Mol. Nutr. Food Res. 2012, 56, 1771–1782. [Google Scholar] [CrossRef] [PubMed]
- Roos, J.; Dahlhaus, M.; Funcke, J.-B.; Kustermann, M.; Strauss, G.; Halbgebauer, D.; Boldrin, E.; Holzmann, K.; Möller, P.; Trojanowski, B.M.; et al. miR-146a Regulates Insulin Sensitivity via NPR3. Cell. Mol. Life Sci. 2021, 78, 2987–3003. [Google Scholar] [CrossRef] [PubMed]
- Karkeni, E.; Bonnet, L.; Marcotorchino, J.; Tourniaire, F.; Astier, J.; Ye, J.; Landrier, J.-F. Vitamin D Limits Inflammation-Linked microRNA Expression in Adipocytes in Vitro and in Vivo: A New Mechanism for the Regulation of Inflammation by Vitamin D. Epigenetics 2018, 13, 156–162. [Google Scholar] [CrossRef]
- Haroun, N.; Bennour, I.; Seipelt, E.; Astier, J.; Couturier, C.; Mounien, L.; Landrier, J.-F. Maternal Vitamin D Deficiency in Mice Increases White Adipose Tissue Inflammation in Offspring. Cells 2022, 11, 2024. [Google Scholar] [CrossRef]
- Karkeni, E.; Payet, T.; Astier, J.; Sicard, F.; Mounien, L.; Landrier, J.-F. Proposal of a Bioinformatics Approach to Predict Molecular Mechanisms Involved in Inflammatory Response: Case of ATRA and 1,25(OH)2D in Adipocytes. Epigenetics 2023, 18, 2201516. [Google Scholar] [CrossRef]
- Zhu, J.; Bing, C.; Wilding, J.P.H. Vitamin D Receptor Ligands Attenuate the Inflammatory Profile of IL-1β-Stimulated Human White Preadipocytes via Modulating the NF-κB and Unfolded Protein Response Pathways. Biochem. Biophys. Res. Commun. 2018, 503, 1049–1056. [Google Scholar] [CrossRef]
- Mutt, S.J.; Karhu, T.; Lehtonen, S.; Lehenkari, P.; Carlberg, C.; Saarnio, J.; Sebert, S.; Hyppönen, E.; Järvelin, M.-R.; Herzig, K.-H. Inhibition of Cytokine Secretion from Adipocytes by 1,25-Dihydroxyvitamin via the NF-κB Pathway. FASEB J. 2012, 26, 4400–4407. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Trayhurn, P.; Bing, C. 1,25-Dihydroxyvitamin D3 Inhibits the Cytokine-Induced Secretion of MCP-1 and Reduces Monocyte Recruitment by Human Preadipocytes. Int. J. Obes. 2013, 37, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Wilding, J.P.H.; Bing, C. 1,25-Dihydroxyvitamin D3 Protects against Macrophage-Induced Activation of NFκB and MAPK Signalling and Chemokine Release in Human Adipocytes. PLoS ONE 2013, 8, e61707. [Google Scholar] [CrossRef] [PubMed]
- Castellano-Castillo, D.; Morcillo, S.; Clemente-Postigo, M.; Crujeiras, A.B.; Fernandez-García, J.C.; Torres, E.; Tinahones, F.J.; Macias-Gonzalez, M. Adipose Tissue Inflammation and VDR Expression and Methylation in Colorectal Cancer. Clin. Epigenet. 2018, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Youm, Y.-H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The Ketone Metabolite β-Hydroxybutyrate Blocks NLRP3 Inflammasome–Mediated Inflammatory Disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Y.; Chang, W.; Zhang, L.; Syrigos, K.N.; Li, P. Noncoding RNA-Mediated Regulation of Pyroptotic Cell Death in Cancer. Front. Oncol. 2022, 12, 1015587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-H.; Chen, Y.-P.; Yang, X.; Li, C.-Q. Vitamin D3 Levels and NLRP3 Expression in Murine Models of Obese Asthma: Association with Asthma Outcomes. Braz. J. Med. Biol. Res. 2017, 51, e6841. [Google Scholar] [CrossRef] [PubMed]
- Tulk, S.E.; Liao, K.-C.; Muruve, D.A.; Li, Y.; Beck, P.L.; MacDonald, J.A. Vitamin D3 Metabolites Enhance the NLRP3-Dependent Secretion of IL-1β From Human THP-1 Monocytic Cells. J. Cell. Biochem. 2015, 116, 711–720. [Google Scholar] [CrossRef]
- Sun, X.; Zemel, M.B. Calcium and 1,25-Dihydroxyvitamin D3 Regulation of Adipokine Expression. Obesity 2007, 15, 340–348. [Google Scholar] [CrossRef]
- Sun, X.; Zemel, M.B. Calcitriol and Calcium Regulate Cytokine Production and Adipocyte–Macrophage Cross-Talk. J. Nutr. Biochem. 2008, 19, 392–399. [Google Scholar] [CrossRef]
- Mutt, S.J.; Hyppönen, E.; Saarnio, J.; Järvelin, M.-R.; Herzig, K.-H. Vitamin D and Adipose Tissue-More than Storage. Front. Physiol. 2014, 5, 228. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.I.; Kuryłowicz, A.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Kozniewski, K.; Puzianowska-Kuznicka, M. Vitamin D Receptor Gene Expression in Adipose Tissue of Obese Individuals Is Regulated by miRNA and Correlates with the Pro-Inflammatory Cytokine Level. Int. J. Mol. Sci. 2019, 20, 5272. [Google Scholar] [CrossRef] [PubMed]
- Calton, E.K.; Keane, K.N.; Newsholme, P.; Soares, M.J. The Impact of Vitamin D Levels on Inflammatory Status: A Systematic Review of Immune Cell Studies. PLoS ONE 2015, 10, e0141770. [Google Scholar] [CrossRef] [PubMed]
- Mirza, I.; Mohamed, A.; Deen, H.; Balaji, S.; Elsabbahi, D.; Munasser, A.; Naquiallah, D.; Abdulbaseer, U.; Hassan, C.; Masrur, M.; et al. Obesity-Associated Vitamin D Deficiency Correlates with Adipose Tissue DNA Hypomethylation, Inflammation, and Vascular Dysfunction. Int. J. Mol. Sci. 2022, 23, 14377. [Google Scholar] [CrossRef] [PubMed]
- Lontchi-Yimagou, E.; Kang, S.; Goyal, A.; Zhang, K.; You, J.Y.; Carey, M.; Jain, S.; Bhansali, S.; Kehlenbrink, S.; Guo, P.; et al. Insulin-Sensitizing Effects of Vitamin D Repletion Mediated by Adipocyte Vitamin D Receptor: Studies in Humans and Mice. Mol. Metab. 2020, 42, 101095. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Nadeau, M.; Valle, M.; Bellmann, K.; Marette, A.; Tchernof, A.; Gagnon, C. Vitamin D Reduces LPS-Induced Cytokine Release in Omental Adipose Tissue of Women but Not Men. Steroids 2015, 104, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wamberg, L.; Kampmann, U.; Stødkilde-Jørgensen, H.; Rejnmark, L.; Pedersen, S.B.; Richelsen, B. Effects of Vitamin D Supplementation on Body Fat Accumulation, Inflammation, and Metabolic Risk Factors in Obese Adults with Low Vitamin D Levels—Results from a Randomized Trial. Eur. J. Intern. Med. 2013, 24, 644–649. [Google Scholar] [CrossRef]
- Lorente-Cebrián, S.; Eriksson, A.; Dunlop, T.; Mejhert, N.; Dahlman, I.; Åström, G.; Sjölin, E.; Wåhlén, K.; Carlberg, C.; Laurencikiene, J.; et al. Differential Effects of 1α,25-Dihydroxycholecalciferol on MCP-1 and Adiponectin Production in Human White Adipocytes. Eur. J. Nutr. 2012, 51, 335–342. [Google Scholar] [CrossRef]
- Hii, C.S.; Ferrante, A. The Non-Genomic Actions of Vitamin D. Nutrients 2016, 8, 135. [Google Scholar] [CrossRef]
- Pathania, M.; Dhar, M.; Kumar, A.; Saha, S.; Malhotra, R.; Pathania, M.; Dhar, M.; Kumar, A.; Saha, S.; Malhotra, R. Association of Vitamin D Status With Metabolic Syndrome and Its Individual Risk Factors: A Cross-Sectional Study. Cureus 2023, 15, 38344. [Google Scholar] [CrossRef]
- Melguizo-Rodríguez, L.; Costela-Ruiz, V.J.; García-Recio, E.; De Luna-Bertos, E.; Ruiz, C.; Illescas-Montes, R. Role of Vitamin D in the Metabolic Syndrome. Nutrients 2021, 13, 830. [Google Scholar] [CrossRef] [PubMed]
- Barker, T.; Rogers, V.E.; Levy, M.; Templeton, J.; Goldfine, H.; Schneider, E.D.; Dixon, B.M.; Henriksen, V.T.; Weaver, L.K. Supplemental Vitamin D Increases Serum Cytokines in Those with Initially Low 25-Hydroxyvitamin D: A Randomized, Double Blind, Placebo-Controlled Study. Cytokine 2015, 71, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Alsufiani, H.M.; AlGhamdi, S.A.; AlShaibi, H.F.; Khoja, S.O.; Saif, S.F.; Carlberg, C. A Single Vitamin D3 Bolus Supplementation Improves Vitamin D Status and Reduces Proinflammatory Cytokines in Healthy Females. Nutrients 2022, 14, 3963. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Ozorowski, M.; Wódkiewicz, E.; Otto, S.W.; Kubiak, K.; Malinowski, B. Impact of Vitamin D Supplementation on Inflammatory Markers’ Levels in Obese Patients. Curr. Issues Mol. Biol. 2021, 43, 1606–1622. [Google Scholar] [CrossRef] [PubMed]
- Beilfuss, J.; Berg, V.; Sneve, M.; Jorde, R.; Kamycheva, E. Effects of a 1-Year Supplementation with Cholecalciferol on Interleukin-6, Tumor Necrosis Factor-Alpha and Insulin Resistance in Overweight and Obese Subjects. Cytokine 2012, 60, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, A.E.; Flynn, M.G.; Pinkston, C.; Markofski, M.M.; Jiang, Y.; Donkin, S.S.; Teegarden, D. Vitamin D Supplementation during Exercise Training Does Not Alter Inflammatory Biomarkers in Overweight and Obese Subjects. Eur. J. Appl. Physiol. 2012, 112, 3045–3052. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, M.; Witkowska-Sędek, E.; Rumińska, M.; Stelmaszczyk-Emmel, A.; Sobol, M.; Majcher, A.; Pyrżak, B. Vitamin D Effects on Selected Anti-Inflammatory and Pro-Inflammatory Markers of Obesity-Related Chronic Inflammation. Front. Endocrinol. 2022, 13, 920340. [Google Scholar] [CrossRef]
- Shah, S.; Wilson, D.M.; Bachrach, L.K. Large Doses of Vitamin D Fail to Increase 25-Hydroxyvitamin D Levels or to Alter Cardiovascular Risk Factors in Obese Adolescents: A Pilot Study. J. Adolesc. Health Off. Publ. Soc. Adolesc. Med. 2015, 57, 19–23. [Google Scholar] [CrossRef]
- Jorde, R.; Sneve, M.; Torjesen, P.A.; Figenschau, Y.; Gøransson, L.G.; Omdal, R. No Effect of Supplementation with Cholecalciferol on Cytokines and Markers of Inflammation in Overweight and Obese Subjects. Cytokine 2010, 50, 175–180. [Google Scholar] [CrossRef]
- Belenchia, A.M.; Jones, K.L.; Will, M.; Beversdorf, D.Q.; Vieira-Potter, V.; Rosenfeld, C.S.; Peterson, C.A. Maternal Vitamin D Deficiency during Pregnancy Affects Expression of Adipogenic-Regulating Genes Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) and Vitamin D Receptor (VDR) in Lean Male Mice Offspring. Eur. J. Nutr. 2018, 57, 723–730. [Google Scholar] [CrossRef]
- Cheung, M.M.; Dall, R.D.; Shewokis, P.A.; Altasan, A.; Volpe, S.L.; Amori, R.; Singh, H.; Sukumar, D. The Effect of Combined Magnesium and Vitamin D Supplementation on Vitamin D Status, Systemic Inflammation, and Blood Pressure: A Randomized Double-Blinded Controlled Trial. Nutr. Burbank Los Angel. Cty. Calif 2022, 99–100, 111674. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhu, H.; Chen, L.; Huang, Y.; Christen, W.; Cook, N.R.; Copeland, T.; Mora, S.; Buring, J.E.; Lee, I.-M.; et al. Effects of Vitamin D3 and Marine Omega-3 Fatty Acids Supplementation on Biomarkers of Systemic Inflammation: 4-Year Findings from the VITAL Randomized Trial. Nutrients 2022, 14, 5307. [Google Scholar] [CrossRef] [PubMed]
- Pittas, A.G.; Harris, S.S.; Stark, P.C.; Dawson-Hughes, B. The Effects of Calcium and Vitamin D Supplementation on Blood Glucose and Markers of Inflammation in Nondiabetic Adults. Diabetes Care 2007, 30, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Nazarabadi, P.N.; Etemad, Z.; Hoseini, R.; Moradi, F. Anti-Inflammatory Effects of a Period of Aerobic Training and Vitamin D Supplementation in Postmenopausal Women with Metabolic Syndrome. Int. J. Prev. Med. 2022, 13, 60. [Google Scholar] [CrossRef]
- Pasupuleti, P.; Suchitra, M.M.; Bitla, A.R.; Sachan, A. Attenuation of Oxidative Stress, Interleukin-6, High-Sensitivity C-Reactive Protein, Plasminogen Activator Inhibitor-1, and Fibrinogen with Oral Vitamin D Supplementation in Patients with T2DM Having Vitamin D Deficiency. J. Lab. Physicians 2022, 14, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Cheshmazar, E.; Hosseini, A.F.; Yazdani, B.; Razmpoosh, E.; Zarrati, M. Effects of Vitamin D Supplementation on Omentin-1 and Spexin Levels, Inflammatory Parameters, Lipid Profile, and Anthropometric Indices in Obese and Overweight Adults with Vitamin D Deficiency under Low-Calorie Diet: A Randomized Placebo Controlled Trial. Evid.-Based Complement. Altern. Med. ECAM 2020, 2020, 3826237. [Google Scholar] [CrossRef] [PubMed]
- Limonte, C.P.; Zelnick, L.R.; Ruzinski, J.; Hoofnagle, A.N.; Thadhani, R.; Melamed, M.L.; Lee, I.-M.; Buring, J.E.; Sesso, H.D.; Manson, J.E.; et al. Effects of Long-Term Vitamin D and n-3 Fatty Acid Supplementation on Inflammatory and Cardiac Biomarkers in Patients with Type 2 Diabetes: Secondary Analyses from a Randomised Controlled Trial. Diabetologia 2021, 64, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Sinha-Hikim, I.; Duran, P.; Shen, R.; Lee, M.; Friedman, T.C.; Davidson, M.B. Effect of Long Term Vitamin D Supplementation on Biomarkers of Inflammation in Latino and African-American Subjects with Pre-Diabetes and Hypovitaminosis D. Horm. Metab. Res. Horm. Stoffwechselforschung Horm. Metab. 2015, 47, 280–283. [Google Scholar] [CrossRef]
- Neyestani, T.R.; Nikooyeh, B.; Alavi-Majd, H.; Shariatzadeh, N.; Kalayi, A.; Tayebinejad, N.; Heravifard, S.; Salekzamani, S.; Zahedirad, M. Improvement of Vitamin D Status via Daily Intake of Fortified Yogurt Drink Either with or without Extra Calcium Ameliorates Systemic Inflammatory Biomarkers, Including Adipokines, in the Subjects with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2012, 97, 2005–2011. [Google Scholar] [CrossRef]
- Shab-Bidar, S.; Neyestani, T.R.; Djazayery, A.; Eshraghian, M.-R.; Houshiarrad, A.; Kalayi, A.; Shariatzadeh, N.; Khalaji, N.; Gharavi, A. Improvement of Vitamin D Status Resulted in Amelioration of Biomarkers of Systemic Inflammation in the Subjects with Type 2 Diabetes. Diabetes Metab. Res. Rev. 2012, 28, 424–430. [Google Scholar] [CrossRef]
- Kampmann, U.; Mosekilde, L.; Juhl, C.; Moller, N.; Christensen, B.; Rejnmark, L.; Wamberg, L.; Orskov, L. Effects of 12 Weeks High Dose Vitamin D3 Treatment on Insulin Sensitivity, Beta Cell Function, and Metabolic Markers in Patients with Type 2 Diabetes and Vitamin D Insufficiency—A Double-Blind, Randomized, Placebo-Controlled Trial. Metabolism 2014, 63, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Kopp, L.; Schweinlin, A.; Tingö, L.; Hutchinson, A.N.; Feit, V.; Jähnichen, T.; Lehnert, K.; Vetter, W.; Rings, A.; Jensen, M.G.; et al. Potential Modulation of Inflammation and Physical Function by Combined Probiotics, Omega-3 Supplementation and Vitamin D Supplementation in Overweight/Obese Patients with Chronic Low-Grade Inflammation: A Randomized, Placebo-Controlled Trial. Int. J. Mol. Sci. 2023, 24, 8567. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Tian, L.; Xiao, Y.; Huang, G.; Zhang, M. Effect of Vitamin D Supplementation on Some Inflammatory Biomarkers in Type 2 Diabetes Mellitus Subjects: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Ann. Nutr. Metab. 2018, 73, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Jamka, M.; Woźniewicz, M.; Walkowiak, J.; Bogdański, P.; Jeszka, J.; Stelmach-Mardas, M. The Effect of Vitamin D Supplementation on Selected Inflammatory Biomarkers in Obese and Overweight Subjects: A Systematic Review with Meta-Analysis. Eur. J. Nutr. 2016, 55, 2163–2176. [Google Scholar] [CrossRef]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2019, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Hauck, A.K.; Huang, Y.; Hertzel, A.V.; Bernlohr, D.A. Adipose Oxidative Stress and Protein Carbonylation. J. Biol. Chem. 2019, 294, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Nono Nankam, P.A.; Nguelefack, T.B.; Goedecke, J.H.; Blüher, M. Contribution of Adipose Tissue Oxidative Stress to Obesity-Associated Diabetes Risk and Ethnic Differences: Focus on Women of African Ancestry. Antioxidant 2021, 10, 622. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Ima-Nirwana, S. Vitamin C: A Review on Its Role in the Management of Metabolic Syndrome. Int. J. Med. Sci. 2020, 17, 1625–1638. [Google Scholar] [CrossRef]
- Lefranc, C.; Friederich-Persson, M.; Palacios-Ramirez, R.; Nguyen Dinh Cat, A. Mitochondrial Oxidative Stress in Obesity: Role of the Mineralocorticoid Receptor. J. Endocrinol. 2018, 238, R143–R159. [Google Scholar] [CrossRef]
- Farhangi, M.; Mesgari-Abbasi, M.; Hajiluian, G.; Nameni, G.; Shahabi, P. Adipose Tissue Inflammation and Oxidative Stress: The Ameliorative Effects of Vitamin D. Inflammation 2017, 40, 1688–1697. [Google Scholar] [CrossRef]
- Teixeira, T.M.; da Costa, D.C.; Resende, A.C.; Soulage, C.O.; Bezerra, F.F.; Daleprane, J.B. Activation of Nrf2-Antioxidant Signaling by 1,25-Dihydroxycholecalciferol Prevents Leptin-Induced Oxidative Stress and Inflammation in Human Endothelial Cells. J. Nutr. 2017, 147, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Orimo, M.; Shimizu, I.; Kunieda, T.; Yokoyama, M.; Ito, T.; Nojima, A.; Nabetani, A.; Oike, Y.; Matsubara, H.; et al. A Crucial Role for Adipose Tissue P53 in the Regulation of Insulin Resistance. Nat. Med. 2009, 15, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Ionica, M.; Aburel, O.M.; Vaduva, A.; Petrus, A.; Rațiu, S.; Olariu, S.; Sturza, A.; Muntean, D.M. Vitamin D Alleviates Oxidative Stress in Adipose Tissue and Mesenteric Vessels from Obese Patients with Subclinical Inflammation1. Can. J. Physiol. Pharmacol. 2020, 98, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Samouda, H.; De Beaufort, C.; Gilson, G.; Schritz, A.; Vaillant, M.; Ghaddhab, C.; Ruiz-Castell, M.; Huiart, L.; Dohet, F.; Weber, B.; et al. Relationship of Oxidative Stress to Visceral Adiposity in Youth and Role Played by Vitamin D. Pediatr. Diabetes 2020, 21, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Cătoi, A.F.; Iancu, M.; Pârvu, A.E.; Cecan, A.D.; Bidian, C.; Chera, E.I.; Pop, I.D.; Macri, A.M. Relationship between 25 Hydroxyvitamin D, Overweight/Obesity Status, Pro-Inflammatory and Oxidative Stress Markers in Patients with Type 2 Diabetes: A Simplified Empirical Path Model. Nutrients 2021, 13, 2889. [Google Scholar] [CrossRef] [PubMed]
- Wenclewska, S.; Szymczak-Pajor, I.; Drzewoski, J.; Bunk, M.; Śliwińska, A. Vitamin D Supplementation Reduces Both Oxidative DNA Damage and Insulin Resistance in the Elderly with Metabolic Disorders. Int. J. Mol. Sci. 2019, 20, 2891. [Google Scholar] [CrossRef] [PubMed]
- Rakotoarivelo, V.; Lacraz, G.; Mayhue, M.; Brown, C.; Rottembourg, D.; Fradette, J.; Ilangumaran, S.; Menendez, A.; Langlois, M.-F.; Ramanathan, S. Inflammatory Cytokine Profiles in Visceral and Subcutaneous Adipose Tissues of Obese Patients Undergoing Bariatric Surgery Reveal Lack of Correlation With Obesity or Diabetes. eBioMedicine 2018, 30, 237–247. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Concept | Search Terms |
|---|---|---|
| PUBMED | ||
| 1 | Vitamin D | (‘vitamin D’ [MeSH Terms] OR ‘ergocalciferols’ [MeSH Terms] OR ‘vitamin D’ [All Fields] OR ‘ergocalciferols’ [MeSH Terms] OR ‘ergocalciferols’ [All Fields]) OR ‘treatmen’ [Title/Abstract]) OR ‘supplementation’ [Title/Abstract] OR ‘vitamin D3’ [Title/Abstract] OR ‘vitamin D2’ [Title/Abstract]) |
| 2 | Weight | (‘obesity’ OR ‘overweight’ OR ‘fat’ OR ‘obese’ OR ‘unhealthy weight’ OR ‘high BMI’) |
| 3 | Senescence | (‘senescence’ OR ‘senescent cells’ OR “aging” [MeSH Terms] OR ‘aging’ [All Fields] OR ‘senescence’ [All Fields] OR ‘senesce’ [All Fields] OR ‘senesced’ [All Fields] OR ‘senescences’ [All Fields] OR ‘senescent’ [All Fields] OR ‘senescents’ [All Fields] OR ‘senesces’ [All Fields] OR ‘senescing’ [All Fields]) |
| 3 | TLR | (‘Toll-Like Receptors’ OR ‘TLR’) |
| 4 | NLRP3 inflammasome | (‘NLRP3 Inflammasome’) OR ‘inflammasome’ |
| 5 | Oxidative stress | (‘oxidative stress’ OR ‘reactive oxygen species’ OR ‘ROS’ OR ‘oxidative stress’ [MeSH Terms] OR (‘oxidative’ [All Fields] AND ‘stress’ [All Fields]) OR ‘oxidative stress’ [All Fields]) |
| 6 | Inflammatory markers | (‘inflammatory markers’ OR ‘cytokines’ OR ‘interleukins’ OR ’CRP’ OR ‘C-reactive protein’ OR ‘tumor necrosis factor’ OR ‘TNFα’ OR ‘adipokines’ [MeSH Terms] OR ‘adipokines’ [All Fields] OR ‘adipokine’ [All Fields]) |
| 7 | NF-κB pathway | (‘NF-kappaB signaling pathway’ OR ‘NF-κB pathway’) |
| 8 | CRP | (‘high sensitivity C-reactive protein’ [MeSH Terms]) OR ‘high-sensitivity C-reactive protein’ [MeSH Terms]) OR ‘C-reactive protein’ [MeSH Terms]) OR ‘high-sensitive C-reactive protein’ [MeSH Terms]) OR ‘high sensitive C-reactive protein’ [MeSH Terms]) OR ‘CRP’ [Title/Abstract]) OR ‘hsCRP’ [Title/Abstract]) |
| 3 | Combinations | 1 AND 2 AND 3; 1 AND 2 AND 4; 1 AND 2 AND 5; 1 AND 2 AND 5; 1 AND 2 AND 6; 1 AND 2 AND 7 |
| EMBASE/EBSCO | ||
| 1 | Adipose tissue | (‘adipocyte inflammation’ OR ‘adipose tissue inflammation’ OR ‘interleukin’ OR ‘inflammasomes’ OR ‘adipos * inflammageing’) |
| 2 | Vitamin D | (‘vitamin D’ OR ‘free vitamin D’ OR ‘vitamin D action’ OR ‘vitamin D metabolism’ OR ‘classical action of vitamin D’ OR ‘non-classical action of vitamin D’ OR ‘genomic action of vitamin D’ OR ‘non-genomic action of vitamin D’ OR ‘vitamin D receptor’ OR ‘vitamin D deficiency’ OR ‘vitamin D’ OR ‘vitamin D supplementation’) |
| 3 | Obesity | (‘obesity’ OR ‘overweight’ OR ‘body mass index’ OR ‘fat mass’ OR ‘abdominal obesity’ OR ‘visceral adipose tissue’ OR ‘subcutaneous adipose tissue’ OR ‘human adipose tissue’ OR ‘human preadipocyte’ OR ‘human adipocyte’ OR ‘adipocyte differentiation’ OR ‘adipogenesis’ OR ‘adipose tissue function’ OR ‘weight loss’ OR ‘bariatric surgery’ OR ‘gastric bypass surgery’) |
| 4 | Inflammation | (‘adipocyte inflammation’ OR ‘adipose tissue inflammation’ OR ‘interleukin’ OR ‘inflammasomes’ OR ‘inflammation’ OR ‘inflammatory cytokines’ OR ‘adipokines’ OR ‘cytokine’ OR ‘immune response’ OR ‘leptin’ OR ‘adiponectin’ OR ‘TNF-α’ OR ‘C-reactive protein’ OR ‘interleukin’ OR ‘insulin secretion’ OR ‘insulin resistance’ OR ‘glucose homeostasis’ OR ‘HOMA-B’ OR ‘HOMA-IR’) |
| 5 | Senescence | (‘adipocyte senescence’ OR ‘adipose tissue senescence’ OR ‘senescence’ OR ‘senescent’ OR ‘adipose inflammaging’) |
| Combinations | (1 OR 2 OR 3) AND 4; (1 OR 2 OR 3) AND 5 | |
| Changes Associated with Senescence | Changes Associated with Obesity |
|---|---|
| Accumulation of senescent cells in AT [19]. | Increases in fat mass and the number of large, hypertrophic adipocytes [7]. |
| Elevated SA β-Gal activity indicative of cellular ageing [24]. | Redistribution of fat from subcutaneous to visceral depots, contributing to metabolic dysfunction [2,26]. |
| Presence of senescent cells exhibiting a SASP [24]. | Increased inflammation and infiltration of immune cells [7]. |
| Increased secretion of inflammatory and profibrotic factors [33]. | Dysfunction of endothelial cells leading to vascular challenges and inflammation [35]. |
| Impaired preadipocyte differentiation and functionality [24]. | Hypoxia due to insufficient vascularization [36]. |
| Increased immune cell infiltration driven by factors secreted by senescent cells [1]. | Insulin resistance and risk of type 2 diabetes mellitus (T2DM) [25]. |
| Shift in AT distribution and function [26,38]. | Obesity-related lipotoxicity and metabolic imbalances [39]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popa, A.D.; Niță, O.; Caba, L.; Gherasim, A.; Graur, M.; Mihalache, L.; Arhire, L.I. From the Sun to the Cell: Examining Obesity through the Lens of Vitamin D and Inflammation. Metabolites 2024, 14, 4. https://doi.org/10.3390/metabo14010004
Popa AD, Niță O, Caba L, Gherasim A, Graur M, Mihalache L, Arhire LI. From the Sun to the Cell: Examining Obesity through the Lens of Vitamin D and Inflammation. Metabolites. 2024; 14(1):4. https://doi.org/10.3390/metabo14010004
Chicago/Turabian StylePopa, Alina Delia, Otilia Niță, Lavinia Caba, Andreea Gherasim, Mariana Graur, Laura Mihalache, and Lidia Iuliana Arhire. 2024. "From the Sun to the Cell: Examining Obesity through the Lens of Vitamin D and Inflammation" Metabolites 14, no. 1: 4. https://doi.org/10.3390/metabo14010004