
Analysis of Relationships between Metabolic Changes and Selected Nutrient Intake in Women Environmentally Exposed to Arsenic


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Urine Collection and As Analysis
2.3. Diet Assessment
2.4. Urine Sample Preparations and Untargeted Metabolomics Analysis
2.5. Data Processing and Statistical Analysis
3. Results
3.1. General Characteristic
3.2. Metabolic Differences between WL and WH Groups and Their Relationship with As
3.3. Association between Dietary Nutrient Intake Involved in iAs Metabolism and Signal Intensity of Putatively Annotated Metabolites
3.3.1. Negative Association
3.3.2. Positive Association
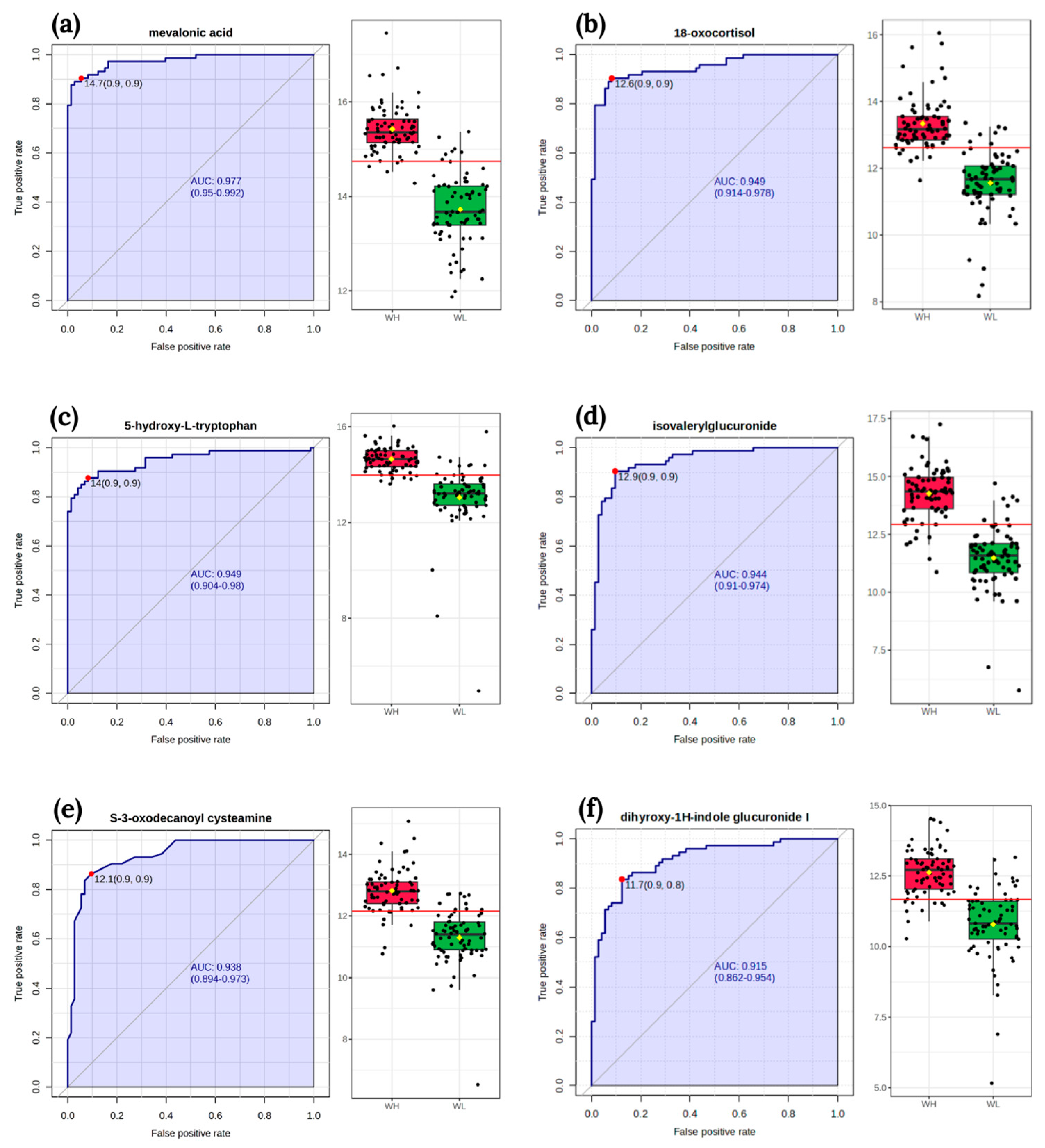
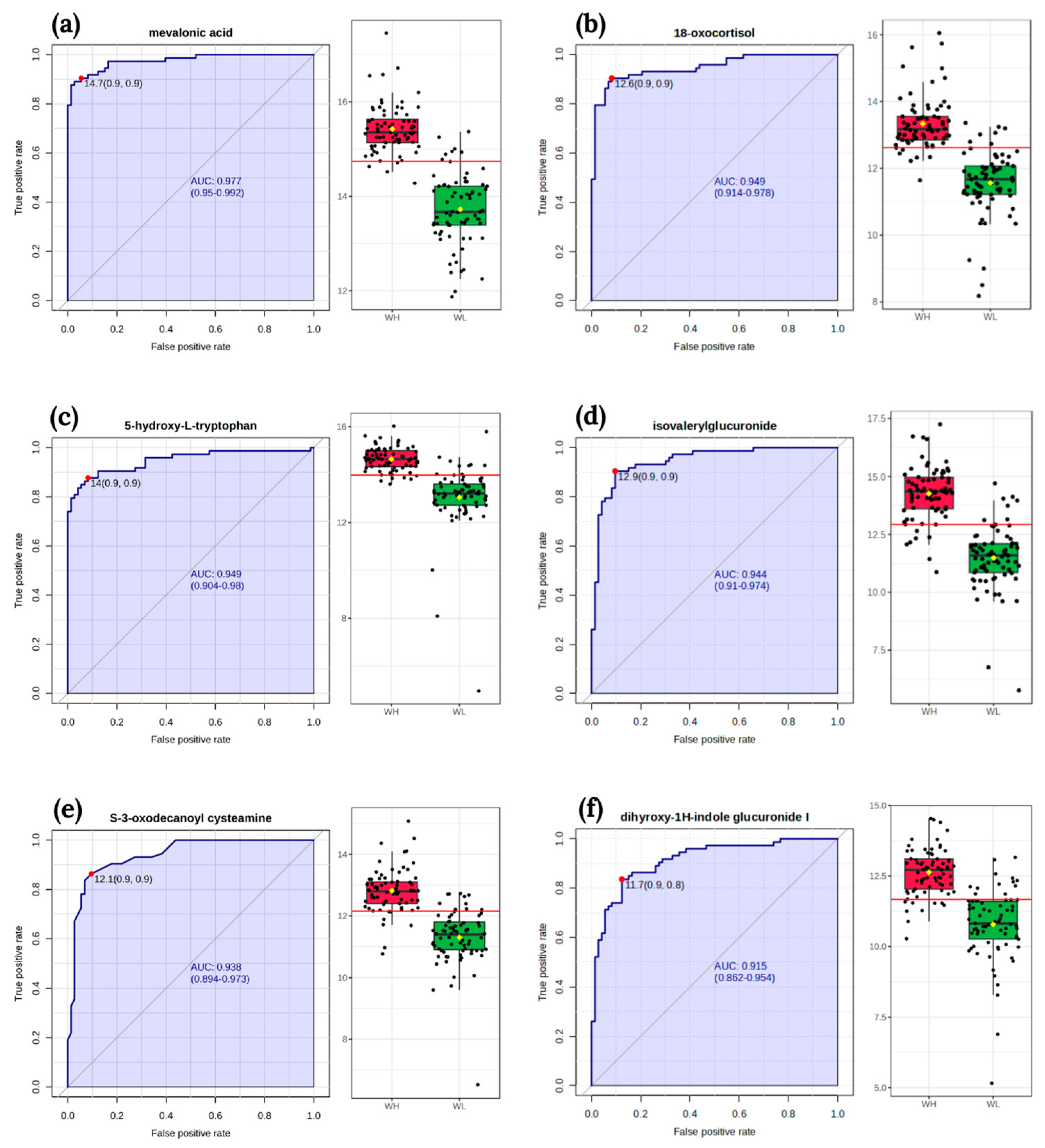
3.4. Biomarker Analysis
4. Discussion
4.1. Urinary As Concentrations
4.2. Metabolites That Were Negatively Associated with Nutrient Intake
4.2.1. Carbohydrate Metabolism
4.2.2. Lipid Metabolism
4.2.3. Amino Acid Metabolism
4.3. Metabolites That Were Positively Associated with Nutrient Intake
4.4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baker, B.A.; Cassano, V.A.; Murray, C. Arsenic Exposure, Assessment, Toxicity, Diagnosis, and Management: Guidance for Occupational and Environmental Physicians. J. Occup. Environ. Med. 2018, 60, 634–639. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); Arcella, D.; Cascio, C.; Gómez Ruiz, J.Á. Chronic Dietary Exposure to Inorganic Arsenic. EFSA J. 2021, 19, e06380. [Google Scholar] [CrossRef]
- International Agency for Research Cancer (IARC). Arsenic, Metals, Fibres, and Dusts. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2012; Volume 100 Pt C, pp. 41–93. ISBN 978-92-832-1320-8. [Google Scholar]
- Chowdhury, R.; Ramond, A.; O’Keeffe, L.M.; Shahzad, S.; Kunutsor, S.K.; Muka, T.; Gregson, J.; Willeit, P.; Warnakula, S.; Khan, H.; et al. Environmental Toxic Metal Contaminants and Risk of Cardiovascular Disease: Systematic Review and Meta-Analysis. BMJ 2018, 362, 3310. [Google Scholar] [CrossRef]
- Rahimi Kakavandi, N.; Mousavi, T.; Asadi, T.; Moradi, A.; Esmaeili, M.; Habibian Sezavar, A.; Nikfar, S.; Abdollahi, M. An Updated Systematic Review and Dose-Response Meta-Analysis on the Relation between Exposure to Arsenic and Risk of Type 2 Diabetes. Toxicol. Lett. 2023, 384, 115–127. [Google Scholar] [CrossRef]
- Jalili, C.; Kazemi, M.; Cheng, H.; Mohammadi, H.; Babaei, A.; Taheri, E.; Moradi, S. Associations between Exposure to Heavy Metals and the Risk of Chronic Kidney Disease: A Systematic Review and Meta-Analysis. Crit. Rev. Toxicol. 2021, 51, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Escudero-Lourdes, C. Toxicity Mechanisms of Arsenic That Are Shared with Neurodegenerative Diseases and Cognitive Impairment: Role of Oxidative Stress and Inflammatory Responses. NeuroToxicology 2016, 53, 223–235. [Google Scholar] [CrossRef]
- Kim, K.; Argos, M.; Persky, V.W.; Freels, S.; Sargis, R.M.; Turyk, M.E. Associations of Exposure to Metal and Metal Mixtures with Thyroid Hormones: Results from the NHANES 2007–2012. Environ. Res. 2022, 212, 113413. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, Z.; Cao, Y.; Wang, J.; Shen, L.; Jiang, T.; Li, D.; Zou, W.; Zong, K.; Liang, D.; et al. Exposure to Multiple Toxic Metals and Polycystic Ovary Syndrome Risk: Endocrine Disrupting Effect from As, Pb and Ba. Sci. Total Environ. 2022, 849, 157780. [Google Scholar] [CrossRef]
- Liu, H.; Lu, S.; Zhang, B.; Xia, W.; Liu, W.; Peng, Y.; Zhang, H.; Wu, K.; Xu, S.; Li, Y. Maternal Arsenic Exposure and Birth Outcomes: A Birth Cohort Study in Wuhan, China. Environ. Pollut. 2018, 236, 817–823. [Google Scholar] [CrossRef]
- Vahter, M. Mechanisms of Arsenic Biotransformation. Toxicology 2002, 181–182, 211–217. [Google Scholar] [CrossRef]
- Cullen, W.R. Chemical Mechanism of Arsenic Biomethylation. Chem. Res. Toxicol. 2014, 27, 457–461. [Google Scholar] [CrossRef] [PubMed]
- El-Ghiaty, M.A.; El-Kadi, A.O.S. The Duality of Arsenic Metabolism: Impact on Human Health. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Sijko, M.; Kozłowska, L. Influence of Dietary Compounds on Arsenic Metabolism and Toxicity. Part II—Human Studies. Toxics 2021, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, L.; Janasik, B.; Nowicka, K.; Wąsowicz, W. A Urinary Metabolomics Study of a Polish Subpopulation Environmentally Exposed to Arsenic. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. GMS 2019, 54, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Janasik, B.; Reszka, E.; Stanislawska, M.; Wieczorek, E.; Fendler, W.; Wasowicz, W. Biological Monitoring and the Influence of Genetic Polymorphism of As3MT and GSTs on Distribution of Urinary Arsenic Species in Occupational Exposure Workers. Int. Arch. Occup. Environ. Health 2015, 88, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Szponar, L.; Wolnicka, K.; Rychlik, E. Album Fotografii Produktów i Potraw; Album of Photographs of Food Products and Dishes; National Food and Nutrition Institute: Warsaw, Poland, 2000; ISBN 83-86060-51-4. [Google Scholar]
- Poslusna, K.; Ruprich, J.; De Vries, J.H.M.; Jakubikova, M.; Van’T Veer, P. Misreporting of Energy and Micronutrient Intake Estimated by Food Records and 24 Hour Recalls, Control and Adjustment Methods in Practice. Br. J. Nutr. 2009, 101, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, M.; Rychlik, E.; Stoś, K.; Charzewska, J. Normy Żywienia Dla Populacji Polski i Ich Zastosowanie; Nutrition Standards for the Polish Population; National Food and Nutrition Institute: Warsaw, Poland, 2020. [Google Scholar]
- Southam, A.D.; Haglington, L.D.; Najdekr, L.; Jankevics, A.; Weber, R.J.M.; Dunn, W.B. Assessment of Human Plasma and Urine Sample Preparation for Reproducible and High-Throughput UHPLC-MS Clinical Metabolic Phenotyping. Anal. 2020, 145, 6511–6523. [Google Scholar] [CrossRef]
- Kozłowska, L.; Santonen, T.; Duca, R.C.; Godderis, L.; Jagiello, K.; Janasik, B.; Van Nieuwenhuyse, A.; Poels, K.; Puzyn, T.; Scheepers, P.T.J.; et al. HBM4EU Chromates Study: Urinary Metabolomics Study of Workers Exposed to Hexavalent Chromium. Metabolites 2022, 12, 362. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, 622–631. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Du, D.; Zheng, W.; Hu, L.; Yang, H.; Cheng, J.; Gong, M. MetaboGroup S: A Group Entropy-Based Web Platform for Evaluating Normalization Methods in Blood Metabolomics Data from Maintenance Hemodialysis Patients. Anal. Chem. 2018, 90, 11124–11130. [Google Scholar] [CrossRef]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC–HRMS Spectra Processing, Multi-Omics Integration and Covariate Adjustment of Global Metabolomics Data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef] [PubMed]
- De Loma, J.; Tirado, N.; Ascui, F.; Levi, M.; Vahter, M.; Broberg, K.; Gardon, J. Elevated Arsenic Exposure and Efficient Arsenic Metabolism in Indigenous Women around Lake Poopó, Bolivia. Sci. Total Environ. 2019, 657, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Roswall, N.; Hvidtfeldt, U.A.; Harrington, J.; Levine, K.E.; Sørensen, M.; Tjønneland, A.; Meliker, J.R.; Raaschou-Nielsen, O. Predictors of Urinary Arsenic Levels among Postmenopausal Danish Women. Int. J. Environ. Res. Public. Health 2018, 15, 1340. [Google Scholar] [CrossRef]
- Wang, X.; Gao, D.; Zhang, G.; Zhang, X.; Li, Q.; Gao, Q.; Chen, R.; Xu, S.; Huang, L.; Zhang, Y.; et al. Exposure to Multiple Metals in Early Pregnancy and Gestational Diabetes Mellitus: A Prospective Cohort Study. Environ. Int. 2020, 135, 105370. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H. A Review on Environmental Factors Regulating Arsenic Methylation in Humans. Toxicol. Appl. Pharmacol. 2009, 235, 338–350. [Google Scholar] [CrossRef]
- Schulz, C.; Wilhelm, M.; Heudorf, U.; Kolossa-Gehring, M. Update of the Reference and HBM Values Derived by the German Human Biomonitoring Commission. Int. J. Hyg. Environ. Health 2011, 215, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Jing, N.; Wang, X.; Yang, X.; Liu, Q.; Wang, H.; Dong, F.; He, K.; Wang, N. Ultra-Performance Liquid Chromatography-Tandem Mass Spectrometry for the Analysis of Complex Compounds in Serum and Its Application in Accurate Detection of Early Arsenic Exposure. ACS Omega 2021, 6, 28326–28333. [Google Scholar] [CrossRef]
- Li, H.; Qin, Q.; Shi, X.; He, J.; Xu, G. Modified Metabolites Mapping by Liquid Chromatography-High Resolution Mass Spectrometry Using Full Scan/All Ion Fragmentation/Neutral Loss Acquisition. J. Chromatogr. A 2019, 1583, 80–87. [Google Scholar] [CrossRef]
- Pullella, K.; Kotsopoulos, J. Arsenic Exposure and Breast Cancer Risk: A Re-Evaluation of the Literature. Nutrients 2020, 12, 3305. [Google Scholar] [CrossRef]
- Moslehi, R.; Stagnar, C.; Srinivasan, S.; Radziszowski, P.; Carpenter, D.O. The Possible Role of Arsenic and Gene-Arsenic Interactions in Susceptibility to Breast Cancer: A Systematic Review. Rev. Environ. Health 2021, 36, 523–534. [Google Scholar] [CrossRef]
- Wu, F.; Chi, L.; Ru, H.; Parvez, F.; Slavkovich, V.; Eunus, M.; Ahmed, A.; Islam, T.; Rakibuz-Zaman, M.; Hasan, R.; et al. Arsenic Exposure from Drinking Water and Urinary Metabolomics: Associations and Long-Term Reproducibility in Bangladesh Adults. Environ. Health Perspect. 2018, 126, 017005. [Google Scholar] [CrossRef]
- Hine, D.G.; Tanaka, K. The Identification and the Excretion Pattern of Isovaleryl Glucuronide in the Urine of Patients with Isovaleric Acidemia. Pediatr. Res. 1984, 18, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; de Santiago, I.; Gopalacharyulu, P.; Schmitt, W.D.; Budczies, J.; Kuhberg, M.; Dietel, M.; Aittokallio, T.; Markowetz, F.; Denkert, C.; et al. Accumulated Metabolites of Hydroxybutyric Acid Serve as Diagnostic and Prognostic Biomarkers of Ovarian High-Grade Serous Carcinomas. Cancer Res. 2016, 76, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Kuhara, T.; Inoue, Y.; Matsumoto, I.; Iwasaki, R.; Morimoto, S. Increased Urinary Excretion of β-Hydroxyisovaleric Acid in Ketotic and Nonketotic Type II Diabetes Mellitus. Clin. Chim. Acta 1990, 188, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, M.; Liang, Q.; Jin, S.; Sun, X.; Jiang, Y.; Pan, X.; Zhou, Y.; Peng, Y.; Zhang, B.; et al. Urinary Metabolomics Revealed Arsenic Exposure Related to Metabolic Alterations in General Chinese Pregnant Women. J. Chromatogr. A 2017, 1479, 145–152. [Google Scholar] [CrossRef]
- López-Vicario, C.; Checa, A.; Urdangarin, A.; Aguilar, F.; Alcaraz-Quiles, J.; Caraceni, P.; Amorós, A.; Pavesi, M.; Gómez-Cabrero, D.; Trebicka, J.; et al. Targeted Lipidomics Reveals Extensive Changes in Circulating Lipid Mediators in Patients with Acutely Decompensated Cirrhosis. J. Hepatol. 2020, 73, 817–828. [Google Scholar] [CrossRef]
- Moore, K.P.; Taylor, G.W.; Maltby, N.H.; Siegers, D.; Fuller, R.W.; Dollery, C.T.; Williams, R. Increased Production of Cysteinyl Leukotrienes in Hepatorenal Syndrome. J. Hepatol. 1990, 11, 263–271. [Google Scholar] [CrossRef]
- Lueke, A.J.; Meeusen, J.W.; Donato, L.J.; Gray, A.V.; Butterfield, J.H.; Saenger, A.K. Analytical and Clinical Validation of an LC–MS/MS Method for Urine Leukotriene E4: A Marker of Systemic Mastocytosis. Clin. Biochem. 2016, 49, 979–982. [Google Scholar] [CrossRef]
- Lenders, J.W.M.; Williams, T.A.; Reincke, M.; Gomez-Sanchez, C.E. Diagnosis of Endocrine Disease: 18-Oxocortisol and 18-Hydroxycortisol: Is There Clinical Utility of These Steroids? Eur. J. Endocrinol. 2018, 178, R1–R9. [Google Scholar] [CrossRef]
- Martin, E.; González-Horta, C.; Rager, J.; Bailey, K.A.; Sánchez-Ramírez, B.; Ballinas-Casarrubias, L.; Ishida, M.C.; Gutiérrez-Torres, D.S.; Hernández Cerón, R.; Viniegra Morales, D.; et al. Metabolomic Characteristics of Arsenic-Associated Diabetes in a Prospective Cohort in Chihuahua, Mexico. Toxicol. Sci. 2015, 144, 338–346. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, W.; Zhou, J.; Chen, M.; Huang, X.; Zhu, Y.; Xie, X.; Li, W.; Zhang, Y.; Kan, H.; et al. Metabolomics Analysis of a Mouse Model for Chronic Exposure to Ambient PM2.5. Environ. Pollut. 2019, 247, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Kui, H.; Su, H.; Wang, Q.; Liu, C.; Li, Y.; Tian, Y.; Kong, J.; Sun, G.; Huang, J. Serum Metabolomics Study of Anxiety Disorder Patients Based on LC-MS. Clin. Chim. Acta 2022, 533, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.H.; Li, S.W.; Chang, C.W.; Chen, Y.C.; Liu, Y.H.; Ma, J.T.; Chang, C.P.; Liao, P.C. Rat Hair Metabolomics Analysis Reveals Perturbations of Unsaturated Fatty Acid Biosynthesis, Phenylalanine, and Arachidonic Acid Metabolism Pathways Are Associated with Amyloid-β-Induced Cognitive Deficits. Mol. Neurobiol. 2023, 60, 4373–4395. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, L.; Cao, M.; Guo, L.; Wang, L. Association between Arsenic and Generalized Anxiety Disorder: A Cross-Sectional Study. Women Health Care Issues 2021, 4, 1–5. [Google Scholar] [CrossRef]
- van der Kloet, F.M.; Tempels, F.W.A.; Ismail, N.; van der Heijden, R.; Kasper, P.T.; Rojas-Cherto, M.; van Doorn, R.; Spijksma, G.; Koek, M.; van der Greef, J.; et al. Discovery of Early-Stage Biomarkers for Diabetic Kidney Disease Using Ms-Based Metabolomics (FinnDiane Study). Metabolomics Off. J. Metabolomic Soc. 2012, 8, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Vincent, D.; Hemsworth, J.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.C.; Rochfort, S.J. Effects of Ergotamine on the Central Nervous System Using Untargeted Metabolomics Analysis in a Mouse Model. Sci. Rep. 2021, 11, 19542. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, Q.; Liu, Z.; Xu, P.; Tian, L.; Yan, J.; Li, K.; Lin, B.; Bian, L.; Xi, Z.; et al. The Impact of Subchronic Ozone Exposure on Serum Metabolome and the Mechanisms of Abnormal Bile Acid and Arachidonic Acid Metabolisms in the Liver. Ecotoxicol. Environ. Saf. 2023, 252, 114573. [Google Scholar] [CrossRef]
- Fels, L. Adverse Effects of Chronic Low Level Lead Exposure on Kidney Function-a Risk Group Study in Children. Nephrol. Dial. Transplant. 1998, 13, 2248–2256. [Google Scholar] [CrossRef]
- Metryka, E.; Kupnicka, P.; Kapczuk, P.; Simińska, D.; Tarnowski, M.; Goschorska, M.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages. Int. J. Mol. Sci. 2020, 21, 2254. [Google Scholar] [CrossRef]
- Yao, X.; Sa, R.; Ye, C.; Zhang, D.; Zhang, S.; Xia, H.; Wang, Y.; Jiang, J.; Yin, H.; Ying, H. Effects of Thyroid Hormone Status on Metabolic Pathways of Arachidonic Acid in Mice and Humans: A Targeted Metabolomic Approach. Prostaglandins Other Lipid Mediat. 2015, 118–119, 11–18. [Google Scholar] [CrossRef]
- Xu, H.D.; Luo, W.; Lin, Y.; Zhang, J.; Zhang, L.; Zhang, W.; Huang, S.M. Discovery of Potential Therapeutic Targets for Non-Small Cell Lung Cancer Using High-Throughput Metabolomics Analysis Based on Liquid Chromatography Coupled with Tandem Mass Spectrometry. RSC Adv. 2019, 9, 10905–10913. [Google Scholar] [CrossRef]
- Diez-Fernandez, C.; Hertig, D.; Loup, M.; Diserens, G.; Henry, H.; Vermathen, P.; Nuoffer, J.; Häberle, J.; Braissant, O. Argininosuccinate Neurotoxicity and Prevention by Creatine in Argininosuccinate Lyase Deficiency: An in Vitro Study in Rat Three-dimensional Organotypic Brain Cell Cultures. J. Inherit. Metab. Dis. 2019, 42, 1077–1087. [Google Scholar] [CrossRef]
- Seminotti, B.; Da Silva, J.C.; Ribeiro, R.T.; Leipnitz, G.; Wajner, M. Free Radical Scavengers Prevent Argininosuccinic Acid-Induced Oxidative Stress in the Brain of Developing Rats: A New Adjuvant Therapy for Argininosuccinate Lyase Deficiency? Mol. Neurobiol. 2020, 57, 1233–1244. [Google Scholar] [CrossRef]
- Wei, Y.; Jia, C.; Lan, Y.; Hou, X.; Zuo, J.; Li, J.; Wang, T.; Mao, G. The Association of Tryptophan and Phenylalanine Are Associated with Arsenic-Induced Skin Lesions in a Chinese Population Chronically Exposed to Arsenic via Drinking Water: A Case-Control Study. BMJ Open 2019, 9, 025336. [Google Scholar] [CrossRef]
- Carmean, C.M.; Yokoi, N.; Takahashi, H.; Oduori, O.S.; Kang, C.; Kanagawa, A.; Kirkley, A.G.; Han, G.; Landeche, M.; Hidaka, S.; et al. Arsenic Modifies Serotonin Metabolism through Glucuronidation in Pancreatic β-Cells. Am. J. Physiol. Endocrinol. Metab. 2019, 316, 464–474. [Google Scholar] [CrossRef]
- Huang, M.C.; Douillet, C.; Su, M.; Zhou, K.; Wu, T.; Chen, W.; Galanko, J.A.; Drobná, Z.; Saunders, R.J.; Martin, E.; et al. Metabolomic Profiles of Arsenic (+3 Oxidation State) Methyltransferase Knockout Mice: Effect of Sex and Arsenic Exposure. Arch. Toxicol. 2017, 91, 189–202. [Google Scholar] [CrossRef]
- Yuan, W.; Zhang, J.; Li, S.; Edwards, J.L. Amine Metabolomics of Hyperglycemic Endothelial Cells Using Capillary LC–MS with Isobaric Tagging. J. Proteome Res. 2011, 10, 5242–5250. [Google Scholar] [CrossRef]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; O’Sullivan, J.; Cheng, S.; Rhee, E.P.; et al. 2-Aminoadipic Acid Is a Biomarker for Diabetes Risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef]
- Sell, D.R.; Strauch, C.M.; Shen, W.; Monnier, V.M. 2-Aminoadipic Acid Is a Marker of Protein Carbonyl Oxidation in the Aging Human Skin: Effects of Diabetes, Renal Failure and Sepsis. Biochem. J. 2007, 404, 269–277. [Google Scholar] [CrossRef]
- Osorio-Yáñez, C.; Chin-Chan, M.; Sánchez-Peña, L.C.; Atzatzi-Aguilar, O.G.; Olivares-Reyes, J.A.; Segovia, J.; Del Razo, L.M. The ADMA/DDAH/NO Pathway in Human Vein Endothelial Cells Exposed to Arsenite. Toxicol. Vitr. 2017, 42, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Guo, L.X.; Chen, X.; Sun, J.; Ye, Y.; Sheng, L.; Zhang, Y.; Sun, X.; Ji, J. Long-Term Chronic Arsenic Exposure on Urinary Metabolism in Mice. SSRN Electron. J. 2023, 898, 165499. [Google Scholar] [CrossRef]
- Liabeuf, S.; Glorieux, G.; Lenglet, A.; Diouf, M.; Schepers, E.; Desjardins, L.; Choukroun, G.; Vanholder, R.; Massy, Z.A.; European Uremic Toxin (EUTox) Work Group. Does P-Cresylglucuronide Have the Same Impact on Mortality as Other Protein-Bound Uremic Toxins? PLoS ONE 2013, 8, 67168. [Google Scholar] [CrossRef]
- Zheng, L.; Kuo, C.C.; Fadrowski, J.; Agnew, J.; Weaver, V.M.; Navas-Acien, A. Arsenic and Chronic Kidney Disease: A Systematic Review. Curr. Environ. Health Rep. 2014, 1, 192–207. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. Regulation of the Mevalonate Pathway. Nature 1990, 343, 425–430. [Google Scholar] [CrossRef]
- Lindenthal, B.; Simatupang, A.; Dotti, M.T.; Federico, A.; Lütjohann, D.; von Bergmann, K. Urinary Excretion of Mevalonic Acid as an Indicator of Cholesterol Synthesis. J. Lipid Res. 1996, 37, 2193–2201. [Google Scholar] [CrossRef]
- Qu, C.; Huang, R. Linking the Low-Density Lipoprotein-Cholesterol (LDL) Level to Arsenic Acid, Dimethylarsinic, and Monomethylarsonic: Results from a National Population-Based Study from the NHANES, 2003–2020. Nutrients 2022, 14, 3993. [Google Scholar] [CrossRef]
- Reddy, D.S. Pharmacology of Endogenous Neuroactive Steroids. Crit. Rev. Neurobiol. 2004, 15, 197–234. [Google Scholar] [CrossRef]
- Reddy, D.S. Is There a Physiological Role for the Neurosteroid THDOC in Stress-Sensitive Conditions? Trends Pharmacol. Sci. 2003, 24, 103–106. [Google Scholar] [CrossRef]
- Wetzel, C.H.R.; Vedder, H.; Holsboer, F.; Zieglgänsberger, W.; Deisz, R.A. Bidirectional Effects of the Neuroactive Steroid Tetrahydrodeoxycorticosterone on GABA-Activated Cl- Currents in Cultured Rat Hypothalamic Neurons: THDOC Effects on GABA Currents of Hypothalamic Neurons. Br. J. Pharmacol. 1999, 127, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Jo, E.K. Inosine: A Bioactive Metabolite with Multimodal Actions in Human Diseases. Front. Pharmacol. 2022, 13, 1043970. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zeng, M.; Wang, S.; Cao, B.; Guo, P.; Zhang, Y.; Jia, J.; Zhang, Q.; Zhang, B.; Wang, R.; et al. Thymidine and 2′-Deoxyuridine Reduce Microglial Activation and Improve Oxidative Stress Damage by Modulating Glycolytic Metabolism on the Aβ25-35-Induced Brain Injury. Arch. Biochem. Biophys. 2022, 729, 109377. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Bi, Z.; Li, L.; Wadgaonkar, P.; Qiu, Y.; Almutairy, B.; Zhang, W.; Seno, A.; Thakur, C.; Chen, F. Metabolomic Dynamics of the Arsenic-Transformed Bronchial Epithelial Cells and the Derived Cancer Stem-like Cells. Int. J. Biol. Sci. 2022, 18, 301–314. [Google Scholar] [CrossRef]
- Hasani, M.; Mansour, A.; Asayesh, H.; Djalalinia, S.; Mahdavi Gorabi, A.; Ochi, F.; Qorbani, M. Effect of Glutamine Supplementation on Cardiometabolic Risk Factors and Inflammatory Markers: A Systematic Review and Meta-Analysis. BMC Cardiovasc. Disord. 2021, 21, 190. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Hu, Y.; Chen, X.; Chang, C.; Li, K. Comparative Effects of Different Nutritional Supplements on Inflammation, Nutritional Status, and Clinical Outcomes in Colorectal Cancer Patients: A Systematic Review and Network Meta-Analysis. Nutrients 2023, 15, 2772. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.H.; Chien, Y.W.; Tsai, Y.H.; Hu, Y.M.; Yeh, S.L. Glutamine Reduces the Expression of Leukocyte Integrins Leukocyte Function-Associated Antigen-1 and Macrophage Antigen-1 in Mice Exposed to Arsenic. Nutr. Res. 2008, 28, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.N.; Saleem, A.; Lee, J.Y.; Tokumoto, M.; Hwang, G.W.; Man Chan, H.; Satoh, M. Effects of Long-Term Cadmium Exposure on Urinary Metabolite Profiles in Mice. J. Toxicol. Sci. 2018, 43, 89–100. [Google Scholar] [CrossRef]
- Wang, H.; Tan, L.; Wang, H.F.; Liu, Y.; Yin, R.H.; Wang, W.Y.; Chang, X.L.; Jiang, T.; Yu, J.T. Magnetic Resonance Spectroscopy in Alzheimer’s Disease: Systematic Review and Meta-Analysis. J. Alzheimers Dis. 2015, 46, 1049–1070. [Google Scholar] [CrossRef]
- Whitehurst, T.S.; Osugo, M.; Townsend, L.; Shatalina, E.; Vava, R.; Onwordi, E.C.; Howes, O. Proton Magnetic Resonance Spectroscopy of N-Acetyl Aspartate in Chronic Schizophrenia, First Episode of Psychosis and High-Risk of Psychosis: A Systematic Review and Meta-Analysis. Neurosci. Biobehav. Rev. 2020, 119, 255–267. [Google Scholar] [CrossRef]
- Nabi, M.; Tabassum, N. Role of Environmental Toxicants on Neurodegenerative Disorders. Front. Toxicol. 2022, 4, 837579. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Both Groups | WL | WH | p-Value ** | |||
|---|---|---|---|---|---|---|---|
| n = 146 | n = 73 | n = 73 | |||||
| Mean ± SD | Median (Minimum–Maximum) | Mean ± SD | Median (Minimum–Maximum) | Mean ± SD | Median (Minimum–Maximum) | ||
| Age (years) | 59.2 ± 14.4 | 62.5 (18.0–85.0) | 60.3 ± 15.8 | 65.0 (18.0–84.0) | 58.1 ± 12.9 * | 59.0 (23.0–85.0) | 0.1021 |
| Height (cm) | 163.5 ± 6.5 * | 163.0 (147.0–184.0) | 163.0 ± 6.2 * | 163.0 (152.0–176.0) | 164.0 ± 6.7 * | 164.0 (147.0–184.0) | 0.3659 |
| Body mass (kg) | 69.5 ± 12.6 | 67.5 (47.0–108.0) | 68.3 ± 12.0 | 67.0 (48.0–108.0) | 70.7 ± 13.0 * | 70.0 (47.0–100.0) | 0.2669 |
| As (µg/L) | 14.0 ± 34.8 | 5.0 (0.1–339.5) | 2.5 ± 1.2 * | 2.3 (0.1–5.0) | 25.6 ± 46.5 | 10.8 (5.0–339.5) | 0.0000 |
| Dietary Intake | Both Groups | WL | WH | p-Value ** | |||
|---|---|---|---|---|---|---|---|
| n = 146 | n = 73 | n = 73 | |||||
| Mean ± SD | Median (Minimum–Maximum) | Mean ± SD | Median (Minimum–Maximum) | Mean ± SD | Median (Minimum–Maximum) | ||
| Methionine (mg/kg bm) | 22.14 ± 7.84 | 21.25 (7.37–60.83) | 21.68 ± 6.74 * | 21.64 (7.37–39.57) | 22.61 ± 8.82 | 20.34 (9.22–60.83) | 0.9003 |
| Vitamin B2 (mg/kg bm) | 0.02 ± 0.01 | 0.02 (0.01–0.04) | 0.02 ± 0.01 | 0.02 (0.01–0.04) | 0.02 ± 0.01 * | 0.02 (0.01–0.04) | 0.3298 |
| Vitamin B6 (mg/kg bm) | 0.02 ± 0.01 | 0.02 (0.01–0.06) | 0.02 ± 0.01 * | 0.02 (0.01–0.05) | 0.03 ± 0.01 | 0.02 (0.01–0.06) | 0.0239 |
| Vitamin B12 (µg/kg bm) | 0.03 ± 0.02 | 0.03 (0.01–0.15) | 0.03 ± 0.01 | 0.03 (0.01–0.08) | 0.04 ± 0.02 | 0.03 (0.01–0.15) | 0.6302 |
| Folate (µg/kg bm) | 3.87 ± 1.55 | 3.63 (0.78–11.24) | 3.75 ± 1.46 | 3.52 (1.62–10.03) | 3.99 ± 1.63 | 3.93 (0.78–11.24) | 0.2906 |
| Zinc (mg/kg bm) | 0.12 ± 0.04 | 0.12 (0.05–0.20) | 0.12 ± 0.04 * | 0.12 (0.05–0.20) | 0.12 ± 0.04 * | 0.12 (0.06–0.20) | 0.8613 |
| Metabolites | HMDB ID | R * | p-Value |
|---|---|---|---|
| isovalerylglucuronide | HMDB0002091 | 0.6334 | 0.0000 |
| argininosuccinic acid | HMDB0000052 | 0.5867 | 0.0000 |
| 5-hydroxy-L-tryptophan | HMDB0000472 | 0.6017 | 0.0000 |
| mevalonic acid | HMDB0000227 | 0.7124 | 0.0000 |
| inosine | HMDB0000195 | −0.2082 | 0.0117 |
| 18-oxocortisol | HMDB0000332 | 0.6790 | 0.0000 |
| N-acetyl-L-aspartic acid | HMDB0000812 | 0.5710 | 0.0000 |
| tetrahydrodeoxycorticosterone | HMDB0000879 | 0.4252 | 0.0000 |
| thromboxane B2 | HMDB0003252 | 0.6593 | 0.0000 |
| aminoadipic acid | HMDB0000510 | 0.5776 | 0.0000 |
| deoxyuridine | HMDB0000012 | 0.4520 | 0.0000 |
| S-3-oxodecanoyl cysteamine | HMDB0059773 | 0.6553 | 0.0000 |
| glutamine | HMDB0000641 | 0.4289 | 0.0000 |
| 3-hydroxydecanedioic acid | HMDB0340579 | 0.4524 | 0.0000 |
| L-arginine | HMDB0000517 | 0.2097 | 0.0111 |
| dihyroxy-1H-indole glucuronide I | HMDB0059997 | 0.5758 | 0.0000 |
| 11-trans-LTE4 | HMDB0062286 | 0.4089 | 0.0000 |
| p-cresol glucuronide | HMDB0011686 | 0.3585 | 0.0000 |
| Correlation between Nutrient Intake *** and Metabolites | Both Groups | WL | WH | |||
|---|---|---|---|---|---|---|
| R | p | R | p | R | p | |
| methionine and argininosuccinic acid | NS | −0.3215 ** | 0.0055 | NS | ||
| vitamin B2 and argininosuccinic acid | NS | −0.3099 ** | 0.0076 | NS | ||
| vitamin B12 and argininosuccinic acid | NS | −0.3039 ** | 0.0090 | NS | ||
| folate and 5-hydroxy-L-tryptophan | NS | NS | −0.2328 ** | 0.0475 | ||
| zinc and 5-hydroxy-L-tryptophan | NS | −0.2434 ** | 0.0380 | NS | ||
| vitamin B2 and dihyroxy-1H-indole glucuronide I | NS | NS | −0.2568 * | 0.0280 | ||
| vitamin B6 and dihyroxy-1H-indole glucuronide I | NS | NS | −0.2489 ** | 0.0337 | ||
| vitamin B2 and 11-trans-LTE4 | NS | −0.2457 ** | 0.0361 | NS | ||
| folate and 11-trans-LTE4 | NS | −0.3060 ** | 0.0085 | NS | ||
| zinc and 11-trans-LTE4 | −0.1885 ** | 0.0227 | −0.2530 ** | 0.0308 | −0.2486 * | 0.0340 |
| methionine and isovalerylglucuronide | NS | NS | −0.2428 ** | 0.0385 | ||
| vitamin B2 and isovalerylglucuronide | NS | NS | −0.2983 * | 0.0100 | ||
| vitamin B6 and isovalerylglucuronide | NS | NS | −0.2812 ** | 0.0160 | ||
| folate and isovalerylglucuronide | NS | NS | −0.3754 ** | 0.0011 | ||
| zinc and isovalerylglucuronide | NS | NS | −0.2947 * | 0.0110 | ||
| vitamin B2 and mevalonic acid | NS | −0.2395 ** | 0.0413 | NS | ||
| vitamin B6 and mevalonic acid | NS | −0.2340 * | 0.0460 | NS | ||
| folate and mevalonic acid | NS | −0.2694 ** | 0.0212 | NS | ||
| vitamin B6 and 18-oxocortisol | NS | NS | −0.3558 ** | 0.0020 | ||
| methionine and aminoadipic acid | NS | −0.2366 * | 0.0440 | NS | ||
| vitamin B2 and aminoadipic acid | NS | −0.2943 ** | 0.0115 | NS | ||
| vitamin B12 and aminoadipic acid | NS | −0.3151 ** | 0.0066 | NS | ||
| folate and aminoadipic acid | NS | −0.2398 ** | 0.0410 | NS | ||
| vitamin B2 and 3-hydroxydecanedioic acid | NS | NS | −0.2772 * | 0.0180 | ||
| vitamin B6 and S-3-oxodecanoyl cysteamine | NS | NS | −0.3378 ** | 0.0035 | ||
| methionine and L-arginine | −0.1834 ** | 0.0267 | NS | −0.3256 ** | 0.0049 | |
| vitamin B6 and L-arginine | NS | NS | −0.2451 ** | 0.0366 | ||
| zinc and L-arginine | NS | NS | −0.2623 ** | 0.0250 | ||
| methionine and p-cresol glucuronide | NS | NS | −0.3876 ** | 0.0007 | ||
| vitamin B12 and p-cresol glucuronide | NS | NS | −0.3306 ** | 0.0043 | ||
| zinc and p-cresol glucuronide | NS | NS | −0.3382 ** | 0.0034 | ||
| methionine and thromboxane B2 | NS | NS | −0.2606 ** | 0.0260 | ||
| Correlation between Nutrient Intake *** and Metabolites | Both Groups | WL | WH | |||
|---|---|---|---|---|---|---|
| R | p | R | p | R | p | |
| methionine and inosine | 0.2227 ** | 0.0069 | NS | 0.3426 ** | 0.0030 | |
| vitamin B2 and inosine | 0.2281 ** | 0.0056 | 0.2731 ** | 0.0194 | 0.2551 * | 0.0290 |
| vitamin B6 and inosine | NS | 0.2330 ** | 0.0472 | NS | ||
| vitamin B12 and inosine | 0.1785 ** | 0.0311 | NS | NS | ||
| folate and inosine | 0.1770 ** | 0.0326 | NS | NS | ||
| zinc and inosine | 0.1637 ** | 0.0483 | NS | NS | ||
| vitamin B2 and N-acetyl-L-aspartic acid | NS | NS | 0.3387 ** | 0.0034 | ||
| vitamin B6 and N-acetyl-L-aspartic acid | 0.1693 ** | 0.0411 | NS | 0.3303 ** | 0.0043 | |
| folate and N-acetyl-L-aspartic acid | NS | NS | 0.2839 ** | 0.0149 | ||
| zinc and N-acetyl-L-aspartic acid | NS | NS | 0.2362 ** | 0.0442 | ||
| methionine and tetrahydrodeoxycorticosterone | NS | NS | 0.3372 ** | 0.0035 | ||
| vitamin B2 and tetrahydrodeoxycorticosterone | NS | NS | 0.2508 * | 0.0320 | ||
| vitamin B12 and tetrahydrodeoxycorticosterone | NS | NS | 0.2617 ** | 0.0253 | ||
| methionine and deoxyuridine | 0.1728 ** | 0.0222 | 0.2919 ** | 0.0122 | NS | |
| vitamin B6 and deoxyuridine | 0.2563 ** | 0.0018 | 0.2549 ** | 0.0295 | NS | |
| methionine and glutamine | NS | 0.2995 ** | 0.0100 | NS | ||
| vitamin B2 and glutamine | NS | 0.2816 ** | 0.0158 | NS | ||
| vitamin B6 and glutamine | 0.1994 ** | 0.0158 | 0.2563 ** | 0.0286 | NS | |
| vitamin B12 and glutamine | 0.2010 ** | 0.0150 | 0.3361 ** | 0.0036 | NS | |
| folate and glutamine | 0.2042 ** | 0.0134 | 0.4151 ** | 0.0003 | NS | |
| zinc and glutamine | NS | 0.3369 ** | 0.0036 | NS | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sijko-Szpańska, M.; Kozłowska, L. Analysis of Relationships between Metabolic Changes and Selected Nutrient Intake in Women Environmentally Exposed to Arsenic. Metabolites 2024, 14, 75. https://doi.org/10.3390/metabo14010075
Sijko-Szpańska M, Kozłowska L. Analysis of Relationships between Metabolic Changes and Selected Nutrient Intake in Women Environmentally Exposed to Arsenic. Metabolites. 2024; 14(1):75. https://doi.org/10.3390/metabo14010075
Chicago/Turabian StyleSijko-Szpańska, Monika, and Lucyna Kozłowska. 2024. "Analysis of Relationships between Metabolic Changes and Selected Nutrient Intake in Women Environmentally Exposed to Arsenic" Metabolites 14, no. 1: 75. https://doi.org/10.3390/metabo14010075
APA StyleSijko-Szpańska, M., & Kozłowska, L. (2024). Analysis of Relationships between Metabolic Changes and Selected Nutrient Intake in Women Environmentally Exposed to Arsenic. Metabolites, 14(1), 75. https://doi.org/10.3390/metabo14010075