

Changes in Plasma Free Amino Acid Profile in Endurance Athletes over a 9-Month Training Cycle

 , , , , ,
, , , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Testing Protocol
2.4. PFAA Analysis
2.5. Statistics
3. Results
3.1. Descriptive Characteristics
3.2. Clustering Multidemensional Data
3.3. Plasma Free Amino Acid Concentrations
3.4. Training Status Biomarkers
4. Discussion
4.1. Normal or Abnormal Response?
4.2. Comparison with Other Long-Term Studies
4.3. Metabolic Context
4.3.1. Proteinogenic PFAAs
4.3.2. Non-Proteinogenic PFAAs
4.4. Practical Perspective
4.5. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
The Protocol of Plasma Free Amino Acids Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Amino Acid | Abbrev. | MRM Transition (Q1 → Q3) | Collision Energy (eV) | Limit of Quantitation (μmol∙L−1) | |
|---|---|---|---|---|---|---|
| Analyte | Internal Std. | |||||
| 1 | 1-methylhistidine | 1MHis | 318.2 → 121.1 | 310.2 → 113.1 | 30 | 0.2 |
| 2 | 3-methylhistidine | 3MHis | 318.2 → 121.1 | 310.2 → 113.1 | 30 | 0.2 |
| 3 | alanine | Ala | 238.2 → 121.1 | 230.2 → 113.1 | 30 | 0.2 |
| 4 | anserine | Ans | 389.2 → 121.1 | 381.2 → 113.1 | 30 | 0.5 |
| 5 | arginine | Arg | 323.2 → 121.1 | 315.2 → 113.1 | 30 | 0.5 |
| 6 | argininosuccinic acid | Asa | 439.2 → 121.1 | 431.2 → 113.1 | 50 | 1.0 |
| 7 | asparagine | Asn | 281.2 → 121.1 | 273.2 → 113.1 | 30 | 0.5 |
| 8 | aspartic acid | Asp | 282.1 → 121.1 | 274.1 → 113.1 | 30 | 0.1 |
| 9 | carnosine | Car | 375.2 → 121.1 | 367.2 → 113.1 | 30 | 0.5 |
| 10 | citrulline | Cit | 324.2 → 121.1 | 316.2 → 113.1 | 30 | 0.5 |
| 11 | cystathionine | Cth | 519.3 → 121.1 | 503.3 → 113.1 | 50 | 0.5 |
| 12 | cystine | Cys | 537.2 → 121.1 | 521.2 → 113.1 | 50 | 1.0 |
| 13 | ethanolamine | EtN | 210.2 → 121.1 | 202.2 → 113.1 | 30 | 0.5 |
| 14 | glutamic acid | Glu | 296.2 → 121.1 | 288.2 → 113.1 | 30 | 0.5 |
| 15 | glutamine | Gln | 295.2 → 121.1 | 287.2 → 113.1 | 30 | 0.5 |
| 16 | glycine | Gly | 224.1 → 121.1 | 216.1 → 113.1 | 30 | 1.0 |
| 17 | histidine | His | 304.2 → 121.1 | 296.2 → 113.1 | 30 | 0.5 |
| 18 | homocitrulline | Hcit | 338.2 → 121.1 | 330.2 → 113.1 | 30 | 0.2 |
| 19 | homocystine | Hcy | 565.3 → 121.1 | 549.3 → 113.1 | 50 | 0.5 |
| 20 | hydroxyproline | Hyp | 280.1 → 121.1 | 272.1 → 113.1 | 30 | 0.2 |
| 21 | isoleucine | Ile | 280.2 → 121.1 | 272.2 → 113.1 | 30 | 0.5 |
| 22 | leucine | Leu | 280.2 → 121.1 | 272.2 → 113.1 | 30 | 0.5 |
| 23 | lysine | Lys | 443.3 → 121.1 | 427.3 → 113.1 | 50 | 0.5 |
| 24 | methionine | Met | 298.2 → 121.1 | 290.2 → 113.1 | 30 | 0.1 |
| 25 | ornithine | Orn | 429.3 → 121.1 | 413.3 → 113.1 | 50 | 0.5 |
| 26 | phenylalanine | Phe | 314.2 → 121.1 | 306.2 → 113.1 | 30 | 0.2 |
| 27 | phosphoethanolamine | PEtN | 290.1 → 121.1 | 282.1 → 113.1 | 30 | 0.5 |
| 28 | phosphoserine | PSer | 334.1 → 121.1 | 326.1 → 113.1 | 30 | 0.5 |
| 29 | proline | Pro | 264.2 → 121.1 | 256.2 → 113.1 | 30 | 0.1 |
| 30 | sarcosine | Sar | 238.2 → 121.1 | 230.2 → 113.1 | 30 | 0.2 |
| 31 | serine | Ser | 254.2 → 121.1 | 246.2 → 113.1 | 30 | 0.5 |
| 32 | taurine | Tau | 274.1 → 121.1 | 266.1 → 113.1 | 30 | 0.5 |
| 33 | threonine | Thr | 268.2 → 121.1 | 260.2 → 113.1 | 30 | 0.2 |
| 34 | tryptophan | Trp | 353.2 → 121.1 | 345.2 → 113.1 | 30 | 0.1 |
| 35 | tyrosine | Tyr | 330.2 → 121.1 | 322.2 → 113.1 | 30 | 0.5 |
| 36 | valine | Val | 266.2 → 121.1 | 258.2 → 113.1 | 30 | 0.2 |
| 37 | α-aminoadipic acid | Aad | 310.2 → 121.1 | 302.2 → 113.1 | 30 | 0.2 |
| 38 | α-amino-n-butyric acid | Abu | 252.2 → 121.1 | 244.2 → 113.1 | 30 | 0.2 |
| 39 | β-alanine | bAla | 238.2 → 121.1 | 230.2 → 113.1 | 30 | 0.5 |
| 40 | β-aminoisobutyric acid | bAib | 252.2 → 121.1 | 244.2 → 113.1 | 30 | 0.2 |
| 41 | γ-amino-n-butyric acid | GABA | 252.2 → 121.1 | 244.2 → 113.1 | 30 | 0.05 |
| 42 | δ-hydroxylysine | Hyl | 459.3 → 121.1 | 443.3 → 113.1 | 50 | 0.5 |
Appendix B


References
- Ishikura, K.; Ra, S.-G.; Ohmori, H. Exercise-induced changes in amino acid levels in skeletal muscle and plasma. J. Phys. Fitness Sports Med. 2013, 2, 301–310. [Google Scholar] [CrossRef]
- Kamei, Y.; Hatazawa, Y.; Uchitomi, R.; Yoshimura, R.; Miura, S. Regulation of skeletal muscle function by amino acids. Nutrients 2020, 12, 261. [Google Scholar] [CrossRef]
- Rennie, M.J.; Tipton, K.D. Protein and amino acid metabolism during and after exercise and the effects of nutrition. Annu. Rev. Nutr. 2000, 20, 457–483. [Google Scholar] [CrossRef]
- Gibala, M.J. Regulation of skeletal muscle amino acid metabolism during exercise. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.F.; Regmi, N.; Wu, G. Homeostatic regulation of plasma amino acid concentrations. Front. Biosci. (Landmark Ed.) 2018, 23, 640–655. [Google Scholar] [CrossRef] [PubMed]
- Stone, M.; Clayton, Z.S.; Buono, M.J.; Kern, M. Exercise intensity influences plasma and sweat amino acid concentrations: A crossover trial. J. Sports Med. Phys. Fit. 2022, 62, 525–530. [Google Scholar] [CrossRef]
- Areces, F.; González-Millán, C.; Salinero, J.J.; Abian-Vicen, J.; Lara, B.; Gallo-Salazar, C.; Ruiz-Vicente, D.; Del Coso, J. Changes in serum free amino acids and muscle fatigue experienced during a half-ironman triathlon. PLoS ONE 2015, 10, e0138376. [Google Scholar] [CrossRef]
- Gawedzka, A.; Grandys, M.; Duda, K.; Zapart-Bukowska, J.; Zoladz, J.A.; Majerczak, J. Plasma BCAA concentrations during exercise of varied intensities in young healthy men − the impact of endurance training. PeerJ 2020, 8, e10491. [Google Scholar] [CrossRef]
- Kargotich, S.; Rowbottom, D.G.; Keast, D.; Goodman, C.; Dawson, B.; Morton, A.R. Plasma glutamine changes after high-intensity exercise in elite male swimmers. Res. Sports Med. 2005, 13, 7–21. [Google Scholar] [CrossRef]
- Lehmann, M.; Huonker, M.; Dimeo, F.; Heinz, N.; Gastmann, U.; Treis, N.; Steinacker, J.M.; Keul, J.; Kajewski, R.; Häussinger, D. Serum amino acid concentrations in nine athletes before and after the 1993 Colmar ultra triathlon. Int. J. Sports Med. 1995, 16, 155–159. [Google Scholar] [CrossRef]
- Ikonen, J.N.; Joro, R.; Uusitalo, A.L.; Kyröläinen, H.; Kovanen, V.; Atalay, M.; Tanskanen-Tervo, M.M. Effects of military training on plasma amino acid concentrations and their associations with overreaching. Exp. Biol. Med. (Maywood) 2020, 245, 1029–1038. [Google Scholar] [CrossRef]
- Strüder, H.K.; Hollmann, W.; Platen, P.; Wöstmann, R.; Weicker, H.; Molderings, G.J. Effect of acute and chronic exercise on plasma amino acids and prolactin concentrations and on [3H]ketanserin binding to serotonin2A receptors on human platelets. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 79, 318–324. [Google Scholar] [CrossRef]
- Tanaka, H.; West, K.A.; Duncan, G.E.; Bassett, D.R., Jr. Changes in plasma tryptophan/branched chain amino acid ratio in responses to training volume variation. Int. J. Sports Med. 1997, 18, 270–275. [Google Scholar] [CrossRef] [PubMed]
- van den Baar, M.T.; Fekkes, D.; van den Hoogenband, C.R.; Duivenvoorden, H.; Pepplinkhuizen, L. Plasma amino acids and sports injuries. Amino Acids 2004, 26, 71–76. [Google Scholar] [CrossRef]
- Gwin, J.A.; Hatch-McChesney, A.; Pitts, K.P.; O’Brien, R.P.; Karis, A.J.; Carrigan, C.T.; McClung, J.P.; Karl, J.P.; Margolis, L.M. Initial military training modulates serum fatty acid and amino acid metabolites. Physiol. Rep. 2022, 10, e15385. [Google Scholar] [CrossRef]
- Krause, S.; Langrock, M.; Weiss, M. Influence of seasonal variations in training loads on selected amino acids and parameters of the psychoimmunological network in a swimming team. Int. J. Sports Med. 2002, 23, 380–387. [Google Scholar] [CrossRef]
- Kingsbury, K.J.; Kay, L.; Hjelm, M. Contrasting plasma free amino acid patterns in elite athletes: Association with fatigue and infection. Br. J. Sports Med. 1998, 32, 25–32. [Google Scholar] [CrossRef]
- Bompa, T.; Buzzichelli, C.A. Periodization. Theory and Methodology of Training, 6th ed.; Human Kinetics Publishers: Champaign, IL, USA, 2018; ISBN 1492544809. [Google Scholar]
- Sugimoto, T.; Kamei, Y. Regulation of skeletal muscle function by amino acids, especially by non-proteinogenic amino acids. J. Nutr. Sci. Vitaminol. 2022, 68, S31–S33. [Google Scholar] [CrossRef] [PubMed]
- Corte, Z.; Venta, R. Biological variation of free plasma amino acids in healthy individuals. Clin. Chem. Lab. Med. 2010, 48, 99–104. [Google Scholar] [CrossRef]
- Trinschek, J.; Zieliński, J.; Zarębska, E.A.; Kusy, K. Male and female athletes matched for maximum oxygen uptake per skeletal muscle mass: Equal but still different. J. Sports Med. Phys. Fit. 2023, 63, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Wang, Z.; Heymsfield, S.B.; Baumgartner, R.N.; Gallagher, D. Total-body skeletal muscle mass: Estimation by a new dual-energy X-ray absorptiometry method. Am. J. Clin. Nutr. 2002, 76, 378–383. [Google Scholar] [CrossRef]
- Matysiak, J.; Dereziński, P.; Klupczyńska, A.; Matysiak, J.; Kaczmarek, E.; Kokot, Z.J. Effects of a honeybee sting on the serum free amino acid profile in humans. PLoS ONE 2014, 9, e103533. [Google Scholar] [CrossRef]
- Matomäki, P.; Kainulainen, H.; Kyröläinen, H. Corrected whole blood biomarkers—The equation of Dill and Costill revisited. Physiol. Rep. 2018, 6, e13749. [Google Scholar] [CrossRef]
- Smith, D.J.; Norris, S.R. Changes in glutamine and glutamate concentrations for tracking training tolerance. Med. Sci. Sports Exerc. 2000, 32, 684–689. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Lehmann, M.; Mann, H.; Gastmann, U.; Keul, J.; Vetter, D.; Steinacker, J.M.; Häussinger, D. Unaccustomed high-mileage vs intensity training-related changes in performance and serum amino acid levels. Int. J. Sports Med. 1996, 17, 187–192. [Google Scholar] [CrossRef]
- Trinschek, J.; Zieliński, J.; Kusy, K. Maximal oxygen uptake adjusted for skeletal muscle mass in competitive speed-power and endurance male athletes: Changes in a one-year training cycle. Int. J. Environ. Res. Public Health 2020, 17, 6226. [Google Scholar] [CrossRef] [PubMed]
- Ciekot-Sołtysiak, M.; Kusy, K.; Podgórski, T.; Zieliński, J. Training-induced annual changes in red blood cell profile in highly-trained endurance and speed-power athletes. J. Sports Med. Phys. Fit. 2018, 58, 1859–1866. [Google Scholar] [CrossRef]
- Mougios, V. Reference intervals for serum creatine kinase in athletes. Br. J. Sports Med. 2007, 41, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Pospieszna, B.; Kusy, K.; Słomińska, E.M.; Dudzinska, W.; Ciekot-Sołtysiak, M.; Zieliński, J. The Effect of Training on Erythrocyte Energy Status and Plasma Purine Metabolites in Athletes. Metabolites 2019, 10, 5. [Google Scholar] [CrossRef]
- Włodarczyk, M.; Kusy, K.; Słomińska, E.M.; Krasiński, Z.; Zieliński, J. Change in lactate, ammonia, and hypoxanthine concentrations in a 1-year training cycle in highly trained athletes: Applying biomarkers as tools to assess training status. J. Strength Cond. Res. 2020, 34, 355–364. [Google Scholar] [CrossRef]
- Zarębska, E.A.; Kusy, K.; Słomińska, E.M.; Kruszyna, Ł.; Zieliński, J. Alterations in Exercise-Induced Plasma Adenosine Triphosphate Concentration in Highly Trained Athletes in a One-Year Training Cycle. Metabolites 2019, 9, 230. [Google Scholar] [CrossRef]
- Zieliński, J.; Kusy, K. Training-induced adaptation in purine metabolism in high-level sprinters vs. triathletes. J. Appl. Physiol. 2012, 112, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.R.; Petersen, S.R. Effects of endurance training on transient oxygen uptake response in cyclists. J. Sports Sci. 1998, 16, 733–788. [Google Scholar] [CrossRef]
- Wagenmakers, A.J. Muscle amino acid metabolism at rest and during exercise: Role in human physiology and metabolism. Exerc. Sport Sci. Rev. 1998, 26, 287–314. [Google Scholar] [CrossRef]
- Graham, T.E.; Turcotte, L.P.; Kiens, B.; Richter, E.A. Training and muscle ammonia and amino acid metabolism in humans during prolonged exercise. J. Appl. Physiol. 1995, 78, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; A, J.; Wang, G.; Lu, H.; Huang, X.; Liu, Y.; Zha, W.; Hao, H.; Zhang, Y.; Liu, L.; et al. Metabolomic investigation into variation of endogenous metabolites in professional athletes subject to strength-endurance training. J. Appl. Physiol. 2009, 106, 531–538. [Google Scholar] [CrossRef]
- Hellsten-Westing, Y.; Norman, B.; Balsom, P.D.; Sjödin, B. Decreased resting levels of adenine nucleotides in human skeletal muscle after high-intensity training. J. Appl. Physiol. 1993, 74, 2523–2528. [Google Scholar] [CrossRef]
- Stathis, C.G.; Febbraio, M.A.; Carey, M.F.; Snow, R.J. Influence of sprint training on human skeletal muscle purine nucleotide metabolism. J. Appl. Physiol. 1994, 76, 1802–1809. [Google Scholar] [CrossRef]
- Abe, H. Role of histidine-related compounds as intracellular proton buffering constituents in vertebrate muscle. Biochemistry 2013, 65, 757–765. [Google Scholar]
- Artioli, G.G.; Sale, C.; Jones, R.L. Carnosine in health and disease. Eur. J. Sport Sci. 2019, 19, 30–39. [Google Scholar] [CrossRef]
- Brisola, G.M.P.; Zagatto, A.M. Ergogenic effects of ß-alanine supplementation on different sports modalities: Strong evidence or only incipient findings? J. Strength. Cond. Res. 2019, 33, 253–282. [Google Scholar] [CrossRef]
- Church, D.D.; Hoffman, J.R.; Varanoske, A.N.; Wang, R.; Baker, K.M.; La Monica, M.B.; Beyer, K.S.; Dodd, S.J.; Oliveira, L.P.; Harris, R.C.; et al. Comparison of two β-alanine dosing protocols on muscle carnosine elevations. J. Am. Coll. Nutr. 2017, 36, 608–616. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D.; Ma, Z.; Wang, C.; Gu, S.; Zhou, Z.; Zuo, H. Plasma β-alanine is positively associated with risk of ischemic stroke: A nested case-control study. J. Nutr. 2023, 153, 1162–1169. [Google Scholar] [CrossRef]
- Hawley, J.A.; Hargreaves, M.; Joyner, M.J.; Zierath, J.R. Integrative biology of exercise. Cell 2014, 159, 738–749. [Google Scholar] [CrossRef]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; O’Sullivan, J.; Cheng, S.; Rhee, E.P.; et al. 2-Aminoadipic acid is a biomarker for diabetes risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef]
- Moolenaar, S.H.; Poggi-Bach, J.; Engelke, U.F.; Corstiaensen, J.M.; Heerschap, A.; de Jong, J.G.; Binzak, B.A.; Vockley, J.; Wevers, R.A. Defect in dimethylglycine dehydrogenase, a new inborn error of metabolism: NMR spectroscopy study. Clin. Chem. 1999, 45, 459–464. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Felder, T.K.; Ring-Dimitriou, S.; Auer, S.; Soyal, S.M.; Kedenko, L.; Rinnerthaler, M.; Cadamuro, J.; Haschke-Becher, E.; Aigner, E.; Paulweber, B.; et al. Specific circulating phospholipids, acylcarnitines, amino acids and biogenic amines are aerobic exercise markers. J. Sci. Med. Sport 2017, 20, 700–705. [Google Scholar] [CrossRef]
- Patel, D.; Witt, S.N. Ethanolamine and phosphatidylethanolamine: Partners in health and disease. Oxid. Med. Cell Longev. 2017, 2017, 4829180. [Google Scholar] [CrossRef]
- Tasseva, G.; Bai, H.D.; Davidescu, M.; Haromy, A.; Michelakis, E.; Vance, J.E. Phosphatidylethanolamine deficiency in Mammalian mitochondria impairs oxidative phosphorylation and alters mitochondrial morphology. J. Biol. Chem. 2013, 288, 4158–4173. [Google Scholar] [CrossRef]
- Kurtz, J.A.; VanDusseldorp, T.A.; Doyle, J.A.; Otis, J.S. Taurine in sports and exercise. J. Int. Soc. Sports Nutr. 2021, 18, 39. [Google Scholar] [CrossRef]
- Hansen, S.; Andersen, M.; Cornett, C.; Gradinaru, R.; Grunnet, N. A role for taurine in mitochondrial function. J. Biomed. Sci. 2010, 17 (Suppl. S1), S23. [Google Scholar] [CrossRef]
- Hall, E.C.R.; Semenova, E.A.; Bondareva, E.A.; Borisov, O.V.; Andryushchenko, O.N.; Andryushchenko, L.B.; Zmijewski, P.; Generozov, E.V.; Ahmetov, I.I. Association of muscle fiber composition with health and exercise-related traits in athletes and untrained subjects. Biol. Sport. 2021, 38, 659–666. [Google Scholar] [CrossRef]
- Thirupathi, A.; Pinho, R.A.; Baker, J.S.; István, B.; Gu, Y. Taurine reverses oxidative damages and restores the muscle function in overuse of exercised muscle. Front. Physiol. 2020, 11, 582449. [Google Scholar] [CrossRef]
- Ward, R.J.; Francaux, M.; Cuisinier, C.; Sturbois, X.; De Witte, P. Changes in plasma taurine levels after different endurance events. Amino Acids 1999, 16, 71–77. [Google Scholar] [CrossRef]
- Nagao, K.; Kimura, T. Use of plasma-free amino acids as biomarkers for detecting and predicting disease risk. Nutr. Rev. 2020, 78 (Suppl. S3), 79–85. [Google Scholar] [CrossRef]
- Ferrando, A.A.; Wolfe, R.R.; Hirsch, K.R.; Church, D.D.; Kviatkovsky, S.A.; Roberts, M.D.; Stout, J.R.; Gonzalez, D.E.; Sowinski, R.J.; Kreider, R.B.; et al. International Society of Sports Nutrition Position Stand: Effects of essential amino acid supplementation on exercise and performance. J. Int. Soc. Sports. Nutr. 2023, 20, 2263409. [Google Scholar] [CrossRef]
- Jäger, R.; Kerksick, C.M.; Campbell, B.I.; Cribb, P.J.; Wells, S.D.; Skwiat, T.M.; Purpura, M.; Ziegenfuss, T.N.; Ferrando, A.A.; Arent, S.M.; et al. International Society of Sports Nutrition Position Stand: Protein and exercise. J. Int. Soc. Sports Nutr. 2017, 14, 20. [Google Scholar] [CrossRef]




| Training Phases | ANOVA | ||||
|---|---|---|---|---|---|
| Transition | General | Specific | Competition | p-Value (η2) | |
| Weight (kg) | 75.2 ± 8.9 | 73.5 ± 8.1 | 73.7 ± 7.9 | 73.7 ± 7.7 | 0.052 (0.28) |
| Body Mass Index (kg·m−2) | 22.7 ± 2.4 | 22.2 ± 2.2 | 22.3 ± 2.0 | 22.3 ± 1.9 | 0.058 (0.27) |
| Fat Mass (kg) | 12.2 ± 3.1 | 10.1 ± 2.4 T | 10.0 ± 2.1 T | 10.1 ± 1.9 T | 0.001 (0.54) |
| Fat Mass (%) | 16.0 ± 2.9 | 13.6 ± 2.4 T | 13.4 ± 2.0 T | 13.7 ± 1.7 T | <0.001 (0.54) |
| Skeletal Muscle Mass (kg) | 32.7 ± 4.0 | 32.4 ± 3.5 | 32.8 ± 3.7 | 32.8 ± 3.5 | 0.570 (0.06) |
| Skeletal Muscle Mass (%) | 43.5 ± 1.7 | 44.2 ± 1.5 | 44.4 ± 1.5 T | 44.5 ± 1.1 T | 0.013 (0.30) |
| White Blood Cells (109·L−1) | 4.9 ± 0.8 | 4.6 ± 1.1 | 5.4 ± 1.0 | 4.9 ± 0.9 | 0.161 (0.16) |
| Red Blood Cells (1012·L−1) | 4.9 ± 0.4 | 5.0 ± 0.4 | 4.9 ± 0.4 | 4.8 ± 0.4 | 0.177 (0.15) |
| Hemoglobin (mmol·L−1) | 9.4 ± 0.4 | 9.3 ± 0.6 | 9.2 ± 0.5 | 8.6 ± 0.6 T,G,S | 0.002 (0.52) |
| Hematocrit (%) | 42.4 ± 2.4 | 42.9 ± 2.4 | 43.2 ± 2.6 | 42.4 ± 2.7 | 0.652 (0.05) |
| Creatine Kinase (U·L−1) | 164 ± 118 | 184 ± 89 | 190 ± 149 | 235 ± 138 | 0.480 (0.08) |
| O2max (L·min−1) | 4.84 ± 0.75 | 4.84 ± 0.55 | 4.98 ± 0.67 | 5.04 ± 0.7 | 0.149 (0.16) |
| O2max (L·min−1·kg weight −1) | 64.1 ± 3.8 | 66.0 ± 2.9 | 67.5 ± 3.9 | 68.2 ± 4.1 T | 0.032 (0.25) |
| O2max (L·min−1·kg SMM−1) | 148 ± 11 | 150 ± 10 | 152 ± 12 | 153 ± 11 | 0.281 (0.12) |
| Maximum Heart Rate (bpm) | 191 ± 7 | 188 ± 7 | 190 ± 6 | 188 ± 7 T | 0.032 (0.30) |
| Training Phase | Rest | Exhaustion | Post Exercise | ANOVA Effects p-Value (η2), Post-Hoc | ||||
|---|---|---|---|---|---|---|---|---|
| 15th min | 30th min | Training Phase | Exercise Stage | Interaction | ||||
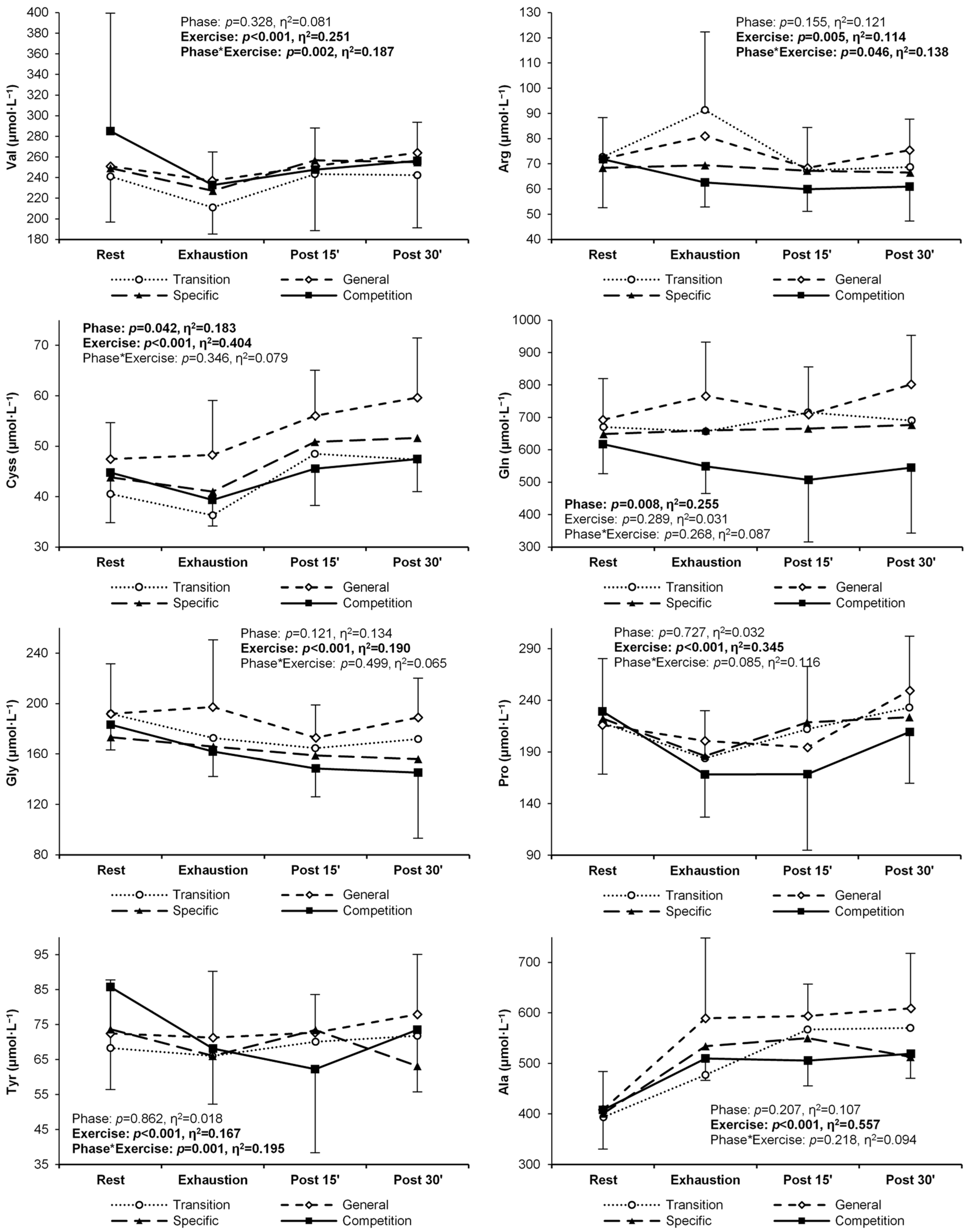
| Histidine | T | 82.6 ± 21.9 | 77.0 ± 14.0 | 90.8 ± 22.0 | 81.5 ± 35.9 | 0.109 (0.14) | <0.001 (0.18) E < P15,P30 | 0.247 (0.09) |
| G | 82.2 ± 10.9 | 84.8 ± 18.9 | 89.3 ± 14.3 | 95.4 ± 22.7 | ||||
| S | 78.1 ± 15.8 | 75.2 ± 13.3 | 90.6 ± 22.3 | 86.0 ± 16.2 | ||||
| C | 79.0 ± 13.8 | 66.7 ± 9.1 | 74.0 ± 12.2 | 76.1 ± 7.3 | ||||
| C | 137 ± 45 | 96 ± 12 R | 102 ± 17 R | 99 ± 14 R | ||||
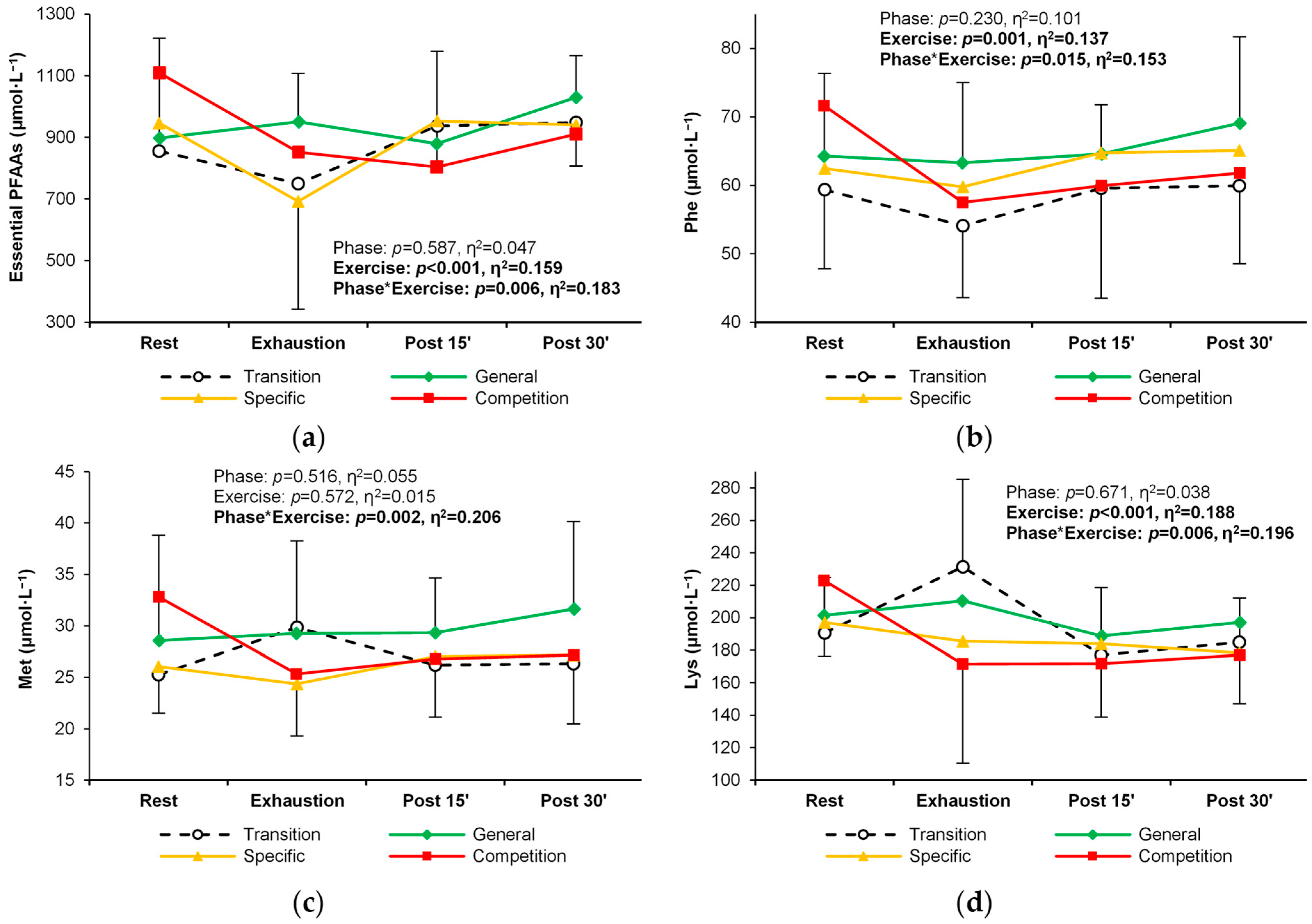
| Lysine | T | 191 ± 34 | 231 ± 54 | 177 ± 41 E | 185 ± 27 E | 0.671 (0.04) | <0.001 (0.19) R > P15,P30 E > P15,P30 | 0.006 (0.20) |
| G | 202 ± 35 | 210 ± 56 | 189 ± 27 | 197 ± 36 | ||||
| S | 197 ± 26 | 185 ± 28 | 184 ± 25 | 178 ± 27 | ||||
| C | 223 ± 47 | 172 ± 61 | 172 ± 33 R | 177 ± 30 R | ||||
| Methionine | T | 25.3 ± 6.3 | 29.8 ± 8.8 | 26.2 ± 9.0 | 26.3 ± 6.2 | 0.516 (0.06) | 0.572 (0.02) | 0.002 (0.21) |
| G | 28.6 ± 10.2 | 29.3 ± 9.0 | 29.3 ± 5.3 | 31.6 ± 8.5 | ||||
| S | 26.1 ± 4.5 | 24.4 ± 5.1 | 27.0 ± 5.9 | 27.2 ± 6.7 | ||||
| C | 32.8 ± 7.3 | 25.3 ± 4.3 R | 26.8 ± 5.1 | 27.2 ± 4.4 | ||||
| Phenylalanine | T | 59.4 ± 11.6 | 54.1 ± 10.5 | 59.6 ± 16.0 | 59.9 ± 11.3 | 0.230 (0.10) | 0.001 (0.14) E < R,P30 | 0.015 (0.15) |
| G | 64.3 ± 12.1 | 63.3 ± 11.7 | 64.6 ± 7.2 | 69.1 ± 12.6 | ||||
| S | 62.5 ± 6.4 | 59.8 ± 8.7 | 64.7 ± 8.8 | 65.1 ± 9.1 | ||||
| C | 71.6 ± 11.3 | 57.5 ± 4.8 R | 59.9 ± 6.1 R | 61.8 ± 4.1 | ||||
| Threonine | T | 131 ± 28 | 108 ± 17 | 113 ± 50 | 129 ± 29 | 0.168 (0.12) | <0.001 (0.43) R > E,P15 E < P15 P30 | 0.188 (0.10) |
| G | 140 ± 31 | 112 ± 44 | 130 ± 22 | 144 ± 31 | ||||
| S | 125 ± 14 | 101 ± 13 | 117 ± 22 | 117 ± 19 | ||||
| C | 136 ± 32 | 88 ± 36 | 109 ± 26 | 117 ± 24 | ||||
| Tryptophan | T | 50.0 ± 10.5 | 34.7 ± 6.6 | 44.2 ± 12.6 | 46.8 ± 10.1 | 0.320 (0.08) | <0.001 (0.61) R > E, P15,P30 E < P15,P30 P15 < P30 | 0.297 (0.08) |
| G | 51.2 ± 7.1 | 40.0 ± 7.0 | 44.4 ± 6.0 | 51.9 ± 9.1 | ||||
| S | 51.3 ± 10.6 | 37.4 ± 8.7 | 44.6 ± 8.8 | 49.9 ± 9.0 | ||||
| C | 50.6 ± 8.0 | 33.8 ± 4.8 | 38.3 ± 6.1 | 43.5 ± 5.8 | ||||
| Isoleucine | T | 61.0 ± 26.8 | 53.4 ± 12.8 | 60.2 ± 16.1 | 58.1 ± 17.1 | 0.527 (0.05) | 0.011 (0.12) R > E,P30 | 0.009 (0.21) |
| G | 60.0 ± 13.8 | 58.3 ± 8.4 | 60.2 ± 7.0 | 61.7 ± 5.2 | ||||
| S | 52.7 ± 9.4 | 50.9 ± 9.9 | 55.8 ± 9.7 | 53.8 ± 12.8 | ||||
| C | 76.4 ± 29.4 S | 52.9 ± 9.3 R | 54.8 ± 10.3 R | 52.7 ± 9.5 R | ||||
| Leucine | T | 105 ± 40 | 84 ± 45 | 113 ± 27 | 111 ± 27 | 0.955 (0.01) | 0.030 (0.09) R > E | 0.002 (0.23) |
| G | 110 ± 21 | 107 ± 17 | 112 ± 14 | 115 ± 14 | ||||
| S | 95 ± 34 | 102 ± 19 | 113 ± 16 | 108 ± 21 | ||||
| C | 137 ± 45 | 96 ± 12 R | 102 ± 17 R | 99 ± 14 R | ||||
| Valine | T | 241 ± 44 | 211 ± 26 | 243 ± 55 | 242 ± 51 | 0.328 (0.08) | <0.001 (0.25) R > E E < P15,P30 | 0.002 (0.19) |
| G | 251 ± 28 | 237 ± 33 | 251 ± 22 | 264 ± 27 | ||||
| S | 249 ± 26 | 227 ± 31 | 257 ± 36 | 255 ± 39 | ||||
| C | 285 ± 114 | 233 ± 32 R | 248 ± 40 R | 256 ± 37 R | ||||
| Total BCAAs | T | 407 ± 107 | 367 ± 53 | 417 ± 96 | 411 ± 93 | 0.681 (0.04) | <0.001 (0.18) R > E E < P15,P30 | 0.002 (0.22) |
| G | 420 ± 58 | 402 ± 56 | 423 ± 39 | 441 ± 39 | ||||
| S | 405 ± 48 | 380 ± 58 | 426 ± 60 | 417 ± 71 | ||||
| C | 516 ± 147 | 381 ± 50 R | 405 ± 66 R | 408 ± 58 R | ||||
| Total essential PFAAs | T | 856 ± 322 | 749 ± 374 R | 937 ± 209 R | 949 ± 170 R | 0.587 (0.05) | <0.001 (0.16) R > E,P15 E < P30 | 0.006 (0.18) |
| G | 898 ± 323 | 951 ± 157 | 879 ± 299 | 1030 ± 136 | ||||
| S | 946 ± 102 | 693 ± 351 | 953 ± 131 | 940 ± 133 | ||||
| C | 1109 ± 225 | 851 ± 84 | 804 ± 296 | 910 ± 108 | ||||
| Training Phase | Rest | Exhaustion | Post Exercise | ANOVA Effects p-Value (η2), Post-Hoc | ||||
|---|---|---|---|---|---|---|---|---|
| 15th min | 30th min | Training Phase | Exercise Stage | Interaction | ||||
| Alanine | T | 393 ± 63 | 477 ± 177 | 567 ± 99 | 570 ± 134 | 0.207 (0.03) | <0.001 (0.35) R < E,P15, P30 | 0.085 (0.12) |
| G | 409 ± 75 | 589 ± 159 | 594 ± 63 | 609 ± 109 | ||||
| S | 401 ± 78 | 534 ± 98 | 550 ± 127 | 513 ± 98 | ||||
| C | 408 ± 78 | 509 ± 43 | 506 ± 50 | 519 ± 49 | ||||
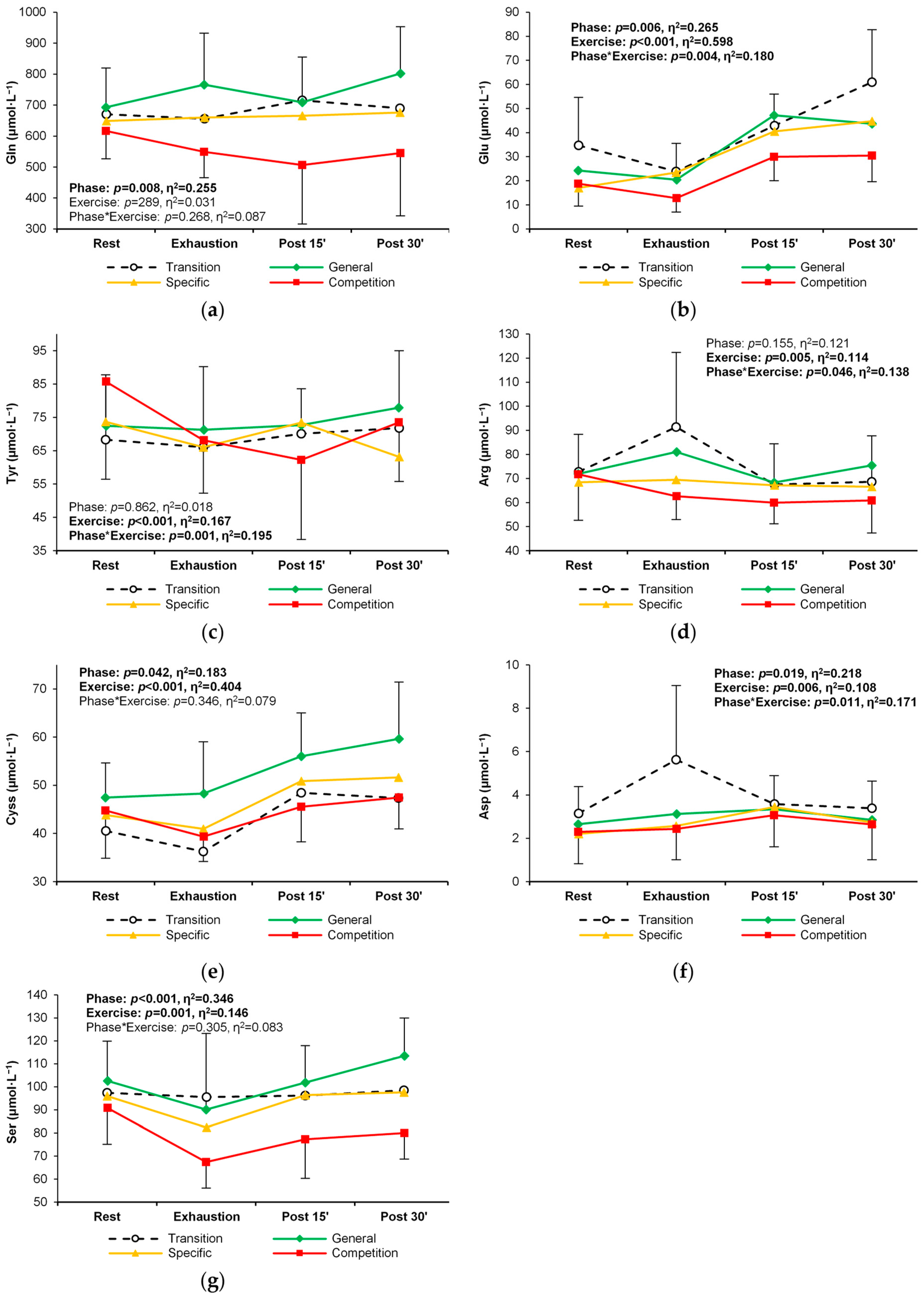
| Arginine | T | 72.7 ± 15.6 | 91.4 ± 30.9 C | 67.5 ± 16.9 E | 68.7 ± 19.0 E | 0.155 (0.12) | 0.005 (0.11) E > P15,P30 | 0.046 (0.14) |
| G | 71.9 ± 17.3 | 81.0 ± 23.8 | 68.3 ± 14.6 | 75.4 ± 16.1 | ||||
| S | 68.4 ± 12.9 | 69.4 ± 13.7 | 67.2 ± 15.3 | 66.5 ± 18.1 | ||||
| C | 71.8 ± 19.1 | 62.6 ± 9.7 | 59.9 ± 8.7 | 60.9 ± 13.6 | ||||
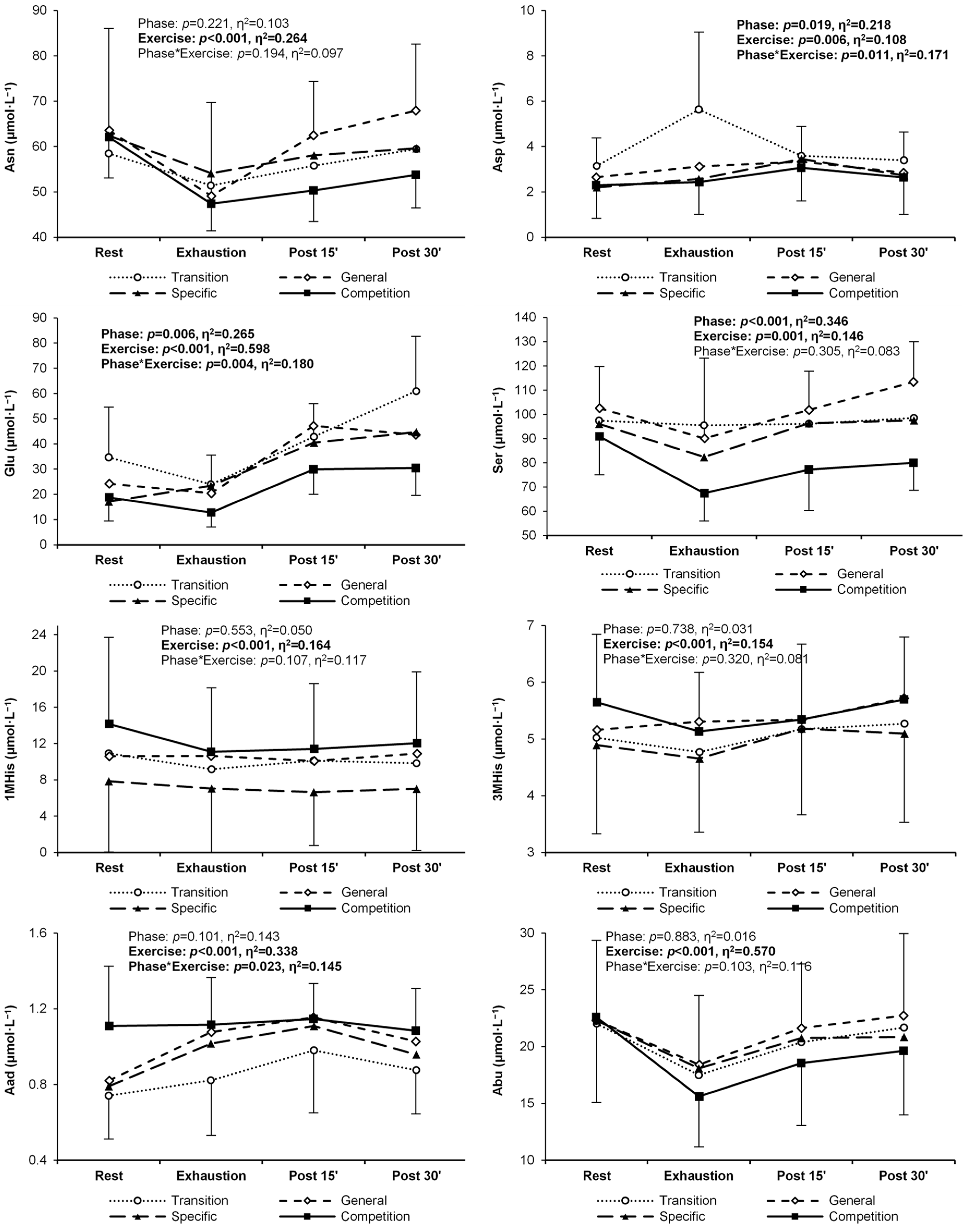
| Asparagine | T | 58.5 ± 9.8 | 51.4 ± 8.9 | 55.8 ± 13.8 | 59.4 ± 14.3 | 0.221 (0.10) | <0.001 (0.26) R > E,P15 | 0.194 (0.10) |
| G | 63.6 ± 22.5 | 49.1 ± 20.6 | 62.5 ± 11.9 | 68.0 ± 14.6 | ||||
| S | 62.4 ± 10.9 | 54.1 ± 9.4 | 58.1 ± 10.3 | 59.6 ± 12.5 | ||||
| C | 62.1 ± 9.0 | 47.4 ± 6.0 | 50.3 ± 6.8 | 53.8 ± 7.3 | ||||
| Aspartate | T | 3.14 ± 1.24 | 5.63 ± 3.42 R | 3.59 ± 1.30 E | 3.39 ± 1.25 E | 0.019 (0.22) T < C | 0.006 (0.11) R < E,P15 | 0.011 (0.17) |
| G | 2.66 ± 0.99 | 3.12 ± 1.26 T | 3.34 ± 1.28 | 2.85 ± 0.76 | ||||
| S | 2.20 ± 1.06 | 2.57 ± 1.06 T | 3.45 ± 1.21 | 2.73 ± 1.10 | ||||
| C | 2.30 ± 1.47 | 2.43 ± 1.42 T | 3.06 ± 1.45 | 2.64 ± 1.63 | ||||
| Cystine | T | 40.5 ± 9.1 | 36.3 ± 8.7 | 48.5 ± 12.9 | 47.4 ± 9.6 | 0.042 (0.18) | <0.001 (0.40) R,E < P15,P30 | 0.346 (0.08) |
| G | 47.5 ± 7.2 | 48.3 ± 10.8 | 56.0 ± 9.0 | 59.6 ± 11.8 | ||||
| S | 43.8 ± 10.2 | 41.0 ± 9.0 | 50.9 ± 13.5 | 51.7 ± 13.3 | ||||
| C | 44.8 ± 9.9 | 39.3 ± 5.2 | 45.5 ± 7.3 | 47.4 ± 6.5 | ||||
| Glutamate | T | 34.7 ± 20.0 | 23.9 ± 11.7 | 42.8 ± 13.2 E | 60.9 ± 21.8 R,E,P15 | 0.006 (0.27) T > C | <0.001 (0.60) R,E < P15,P30 | 0.004 (0.18) |
| G | 24.3 ± 11.3 | 20.4 ± 9.2 | 47.1 ± 22.5 R,E | 43.6 ± 13.5 R,E | ||||
| S | 17.1 ± 5.3 | 23.5 ± 14.1 | 40.4 ± 17.1 R,E | 44.7 ± 17.3 R,E | ||||
| C | 18.8 ± 9.3 | 12.7 ± 5.8 | 29.9 ± 9.8 E | 30.5 ± 10.9 T,E | ||||
| Glutamine | T | 670 ± 115 | 656 ± 127 | 715 ± 198 | 690 ± 176 | 0.008 (0.26) G > C | 0.289 (0.03) | 0.268 (0.09) |
| G | 693 ± 127 | 766 ± 166 | 709 ± 147 | 802 ± 151 | ||||
| S | 649 ± 140 | 660 ± 156 | 666 ± 132 | 676 ± 107 | ||||
| C | 617 ± 90 | 549 ± 83 | 507 ± 191 | 545 ± 202 | ||||
| Glycine | T | 192 ± 31 | 173 ± 28 | 165 ± 43 | 172 ± 43 | 0.121 (0.13) | <0.001 (0.19) R > P15,P30 E > P15 | 0.499 (0.07) |
| G | 192 ± 40 | 197 ± 53 | 173 ± 26 | 189 ± 31 | ||||
| S | 173 ± 34 | 166 ± 23 | 159 ± 29 | 156 ± 25 | ||||
| C | 183 ± 20 | 162 ± 20 | 149 ± 23 | 145 ± 52 | ||||
| Proline | T | 218 ± 62 | 184 ± 46 | 212 ± 61 | 233 ± 69 | 0.727 (0.03) | <0.001 (0.35) R > E; E < P15,P30 P15 < P30 | 0.085 (0.12) |
| G | 216 ± 42 | 201 ± 49 | 195 ± 75 | 249 ± 60 | ||||
| S | 222 ± 47 | 186 ± 37 | 219 ± 47 | 224 ± 43 | ||||
| C | 229 ± 61 | 168 ± 41 | 169 ± 74 | 209 ± 50 | ||||
| Serine | T | 97.4 ± 18.4 | 95.5 ± 26.5 | 96.2 ± 22.6 | 98.5 ± 19.5 | <0.001 (0.35) T, G > C | 0.001 (0.15) R > E E < P30 | 0.305 (0.08) |
| G | 93.7 ± 35.0 | 90.1 ± 33.1 | 101.8 ± 16.1 | 113.5 ± 16.5 | ||||
| S | 96.0 ± 14.3 | 82.4 ± 13.8 | 96.3 ± 13.2 | 97.5 ± 15.0 | ||||
| C | 90.9 ± 15.8 | 67.4 ± 11.4 | 77.2 ± 16.9 | 80.0 ± 11.4 | ||||
| Tyrosine | T | 68.3 ± 16.5 | 66.0 ± 14.1 | 70.1 ± 20.4 | 71.8 ± 20.9 | 0.862 (0.02) | <0.001 (0.17) R > E E < P30 | 0.001 (0.21) |
| G | 72.4 ± 15.3 | 71.3 ± 19.0 | 72.7 ± 10.9 | 77.9 ± 17.1 | ||||
| S | 73.7 ± 16.1 | 66.1 ± 12.9 | 73.5 ± 17.7 | 63.2 ± 25.1 | ||||
| C | 85.8 ± 29.3 | 68.1 ± 15.9 R | 62.2 ± 23.9 R | 73.5 ± 17.7 R | ||||
| Total non-essen- tial PFAAs | T | 1849 ± 296 | 1716 ± 637 | 2043 ± 457 | 2075 ± 438 | 0.014 (0.23) G > C | 0.003 (0.13) R < P30 E < P30 | 0.251 (0.09) |
| G | 1894 ± 285 | 2134 ± 454 | 2100 ± 236 | 2290 ± 337 | ||||
| S | 1809 ± 298 | 1696 ± 632 | 1983 ± 339 | 1820 ± 644 | ||||
| C | 1670 ± 583 | 1688 ± 151 | 1730 ± 194 | 1840 ± 163 | ||||
| Training Phase | Rest | Exhaustion | Post Exercise | ANOVA Effects p-Value (η2), Post-Hoc | ||||
|---|---|---|---|---|---|---|---|---|
| 15th min | 30th min | Training Phase | Exercise Stage | Interaction | ||||
| 1-Methylhistidi-ne | T | 10.9 ± 9.4 | 9.2 ± 8.1 | 10.1 ± 8.6 | 9.8 ± 8.2 | 0.553 (0.05) | <0.001 (0.16) R > E,P15,P30 | 0.107 (0.12) |
| G | 10.6 ± 9.3 | 10.6 ± 9.9 | 10.1 ± 9.0 | 10.9 ± 10.1 | ||||
| S | 7.9 ± 7.8 | 7.0 ± 7.3 | 6.6 ± 5.9 | 7.0 ± 6.8 | ||||
| C | 14.2 ± 9.6 | 11.1 ± 7.1 | 11.4 ± 7.2 | 12.0 ± 7.9 | ||||
| 3-Methylhisti- dine | T | 5.02 ± 1.24 | 4.77 ± 1.05 | 5.18 ± 1.35 | 5.27 ± 1.06 | 0.738 (0.03) | <0.001 (0.15) E < P15,P30 | 0.320 (0.08) |
| G | 5.16 ± 1.17 | 5.31 ± 1.62 | 5.34 ± 1.00 | 5.72 ± 1.33 | ||||
| S | 4.89 ± 1.56 | 4.65 ± 1.30 | 5.18 ± 1.52 | 5.10 ± 1.57 | ||||
| C | 5.65 ± 1.20 | 5.13 ± 1.04 | 5.35 ± 1.32 | 5.70 ± 1.10 | ||||
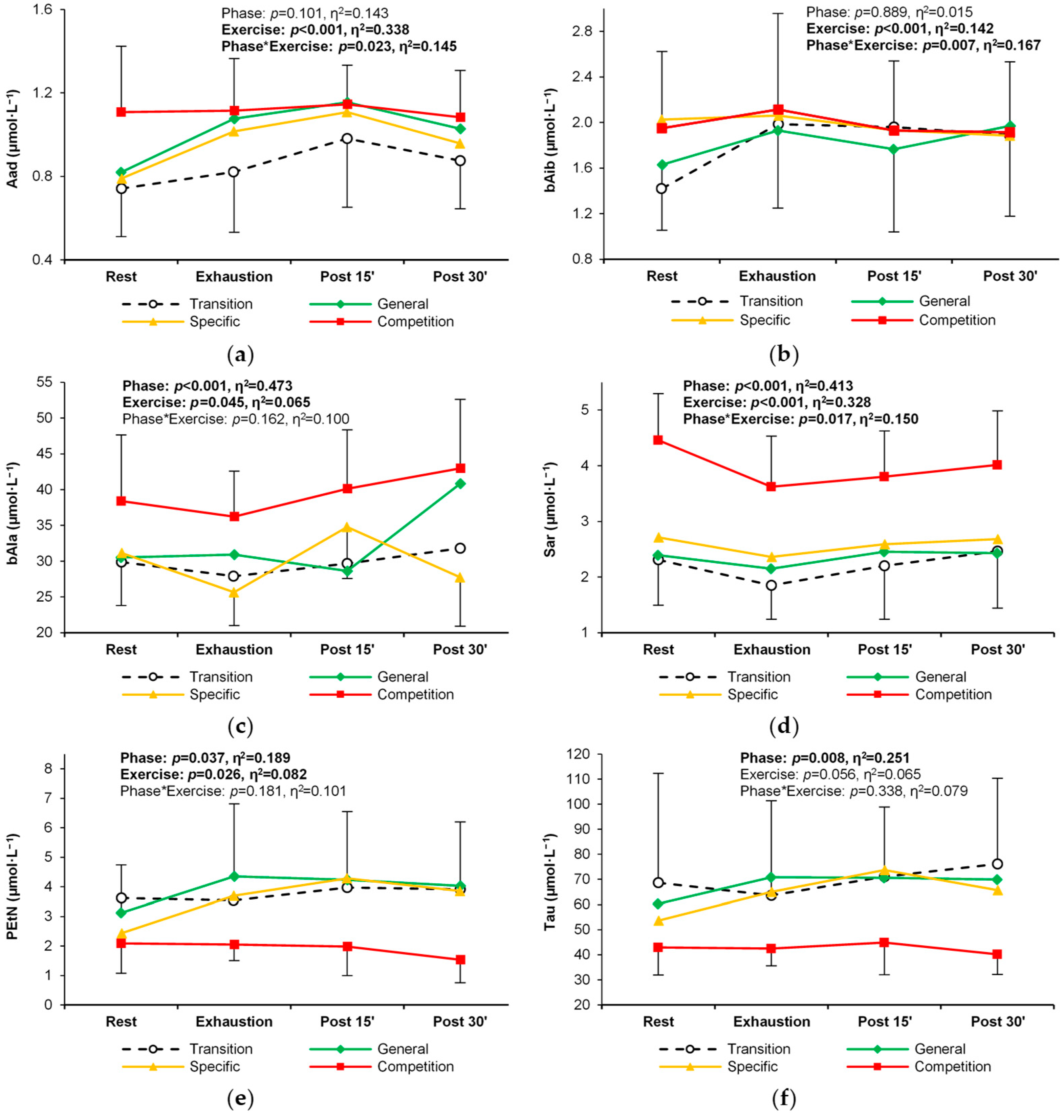
| Aminoadipic acid | T | 0.74 ± 0.23 | 0.82 ± 0.29 | 0.98 ± 0.33 R | 0.88 ± 0.23 | 0.101 (0.14) | <0.001 (0.34) R < E,P15,P30 E < P15 P15 > P30 | 0.023 (0.15) |
| G | 0.82 ± 0.22 | 1.08 ± 0.46 R | 1.15 ± 0.23 R | 1.03 ± 0.25 | ||||
| S | 0.79 ± 0.25 | 1.02 ± 0.34 | 1.11 ± 0.28 R | 0.96 ± 0.21 | ||||
| C | 1.11 ± 0.32 | 1.12 ± 0.25 | 1.15 ± 0.19 | 1.08 ± 0.22 | ||||
| Aminobutyric acid | T | 22.0 ± 7.0 | 17.5 ± 7.5 | 20.4 ± 7.5 | 21.7 ± 8.2 | 0.883 (0.02) | <0.001 (0.57) R > E,P15 E < P15,P30 | 0.103 (0.12) |
| G | 22.3 ± 7.0 | 18.4 ± 6.1 | 21.6 ± 5.6 | 22.7 ± 7.2 | ||||
| S | 22.3 ± 6.8 | 18.1 ± 6.1 | 20.8 ± 6.8 | 20.8 ± 6.1 | ||||
| C | 22.6 ± 7.5 | 15.6 ± 4.4 | 18.5 ± 5.4 | 19.6 ± 5.6 | ||||
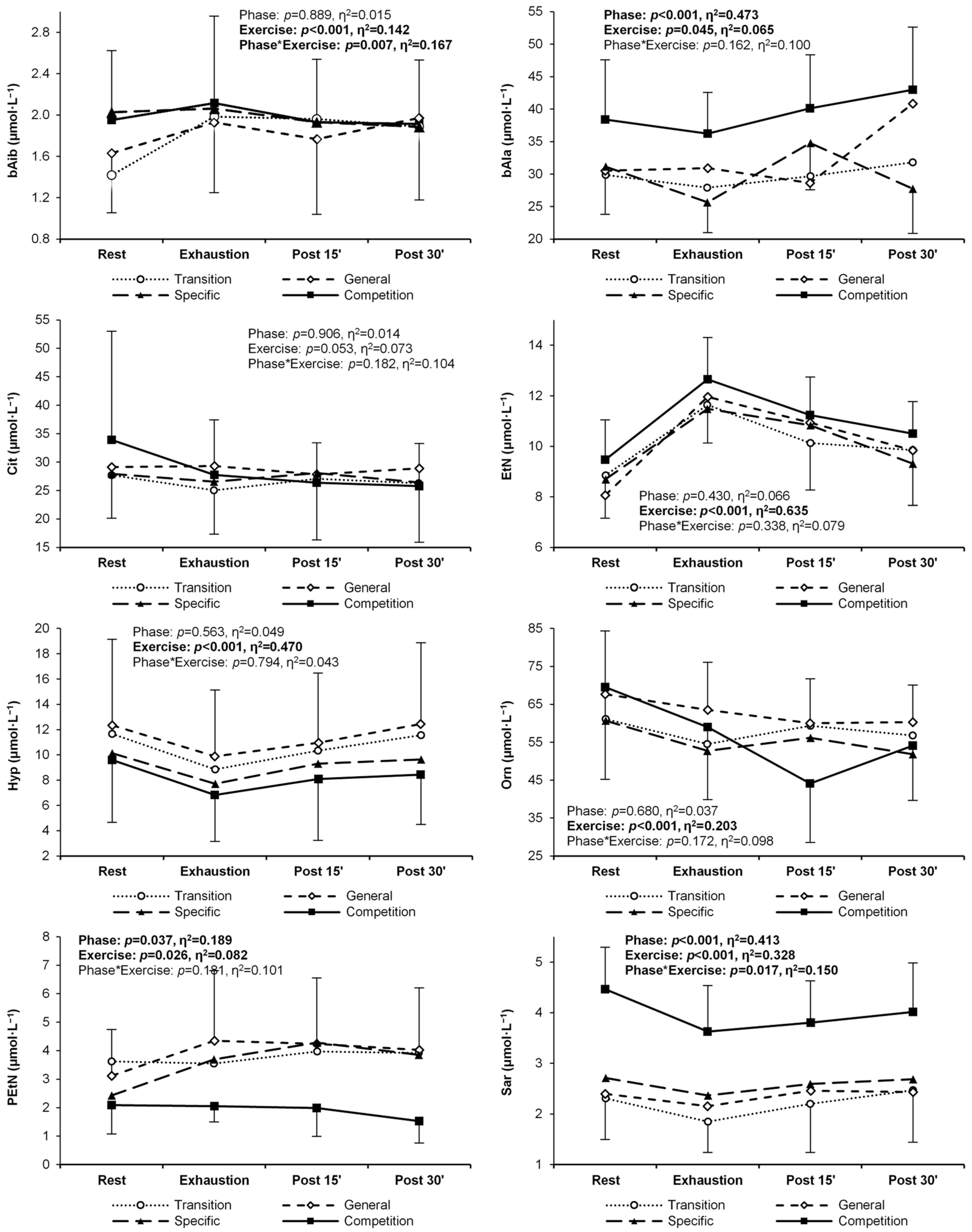
| β-Aminoisobu- tyric acid | T | 1.42 ± 0.56 | 1.98 ± 0.76 R | 1.96 ± 0.98 R | 1.89 ± 0.72 R | 0.889 (0.02) | <0.001 (0.14) R < E | 0.007 (0.17) |
| G | 1.63 ± 0.58 | 1.93 ± 0.68 | 1.77 ± 0.73 | 1.97 ± 0.79 | ||||
| S | 2.03 ± 0.83 | 2.06 ± 0.84 | 1.93 ± 0.77 | 1.88 ± 0.77 | ||||
| C | 1.95 ± 0.67 | 2.11 ± 0.84 | 1.93 ± 0.61 | 1.91 ± 0.62 | ||||
| β-Alanine | T | 29.9 ± 9.1 | 27.9 ± 6.0 | 29.7 ± 6.9 | 31.8 ± 5.9 | <0.001 (0.47) C > T,G,S | 0.045 (0.07) E < P30 | 0.162 (0.10) |
| G | 30.5 ± 9.6 | 30.9 ± 13.0 | 28.6 ± 5.9 | 40.9 ± 19.7 | ||||
| S | 31.2 ± 7.4 | 25.6 ± 4.7 | 34.8 ± 7.2 | 27.7 ± 6.8 | ||||
| C | 38.4 ± 9.2 | 36.2 ± 6.4 | 40.1 ± 8.2 | 43.0 ± 9.6 | ||||
| Citrulline | T | 27.7 ± 7.5 | 25.0 ± 7.7 | 27.0 ± 10.7 | 26.3 ± 10.4 | 0.906 (0.01) | 0.053 (0.07) | 0.182 (0.10) |
| G | 29.1 ± 8.9 | 29.3 ± 9.5 | 27.8 ± 6.2 | 28.9 ± 6.2 | ||||
| S | 27.9 ± 6.6 | 26.5 ± 6.6 | 28.1 ± 6.8 | 26.4 ± 7.0 | ||||
| C | 33.9 ± 19.1 | 27.7 ± 9.7 | 26.4 ± 7.0 | 25.8 ± 7.5 | ||||
| Ethanolamine | T | 8.8 ± 1.7 | 11.6 ± 1.5 | 10.1 ± 1.9 | 9.8 ± 2.2 | 0.430 (0.07) | <0.001 (0.64) R < E,P15,P30 E > P15,P30 P15 < P30 | 0.338 (0.08) |
| G | 8.1 ± 0.9 | 12.0 ± 1.6 | 10.9 ± 1.8 | 9.8 ± 1.7 | ||||
| S | 8.7 ± 1.8 | 11.5 ± 2.4 | 10.8 ± 1.9 | 9.3 ± 1.6 | ||||
| C | 9.5 ± 1.6 | 12.7 ± 1.7 | 11.2 ± 1.5 | 10.5 ± 1.3 | ||||
| Hydroxyproli-ne | T | 11.7 ± 7.0 | 8.8 ± 6.2 | 10.3 ± 6.1 | 11.6 ± 7.6 | 0.563 (0.05) | <0.001 (0.47) R > E,P15 E < P15,P30 P15 > P30 | 0.794 (0.04) |
| G | 12.3 ± 6.8 | 9.9 ± 5.2 | 11.0 ± 5.5 | 12.5 ± 6.4 | ||||
| S | 10.1 ± 6.3 | 7.7 ± 4.9 | 9.3 ± 5.4 | 9.6 ± 6.2 | ||||
| C | 9.6 ± 4.9 | 6.8 ± 3.7 | 8.1 ± 4.9 | 8.4 ± 3.9 | ||||
| Ornithine | T | 61.1 ± 16.9 | 54.5 ± 13.7 | 59.4 ± 25.8 | 56.8 ± 19.0 | 0.680 (0.04) | <0.001 (0.20) R > E,P15,P30 | 0.172 (0.10) |
| G | 67.7 ± 16.7 | 63.6 ± 12.5 | 60.0 ± 11.7 | 60.3 ± 9.8 | ||||
| S | 60.7 ± 12.8 | 52.7 ± 11.1 | 56.2 ± 14.2 | 51.8 ± 12.1 | ||||
| C | 69.6 ± 24.3 | 59.0 ± 19.1 | 44.1 ± 15.6 | 54.1 ± 14.5 | ||||
| Phosphoetha- nolamine | T | 3.62 ± 3.04 | 3.55 ± 2.99 | 3.97 ± 2.14 | 3.92 ± 2.60 | 0.037 (0.19) | 0.026 (0.08) R < P15 | 0.181 (0.10) |
| G | 3.12 ± 1.62 | 4.35 ± 2.46 | 4.24 ± 2.31 | 4.03 ± 2.17 | ||||
| S | 2.43 ± 1.34 | 3.70 ± 1.82 | 4.28 ± 2.34 | 3.85 ± 2.31 | ||||
| C | 2.09 ± 1.01 | 2.05 ± 0.55 | 1.98 ± 0.99 | 1.53 ± 0.78 | ||||
| Sarcosine | T | 2.31 ± 0.81 | 1.85 ± 0.61 | 2.20 ± 0.96 | 2.47 ± 1.03 E | <0.001 (0.41) T,G,S < C | <0.001 (0.33) R > E,P15 E < P15,P30 | 0.017 (0.15) |
| G | 2.39 ± 0.86 | 2.15 ± 0.95 | 2.46 ± 1.03 | 2.43 ± 0.79 | ||||
| S | 2.71 ± 0.94 | 2.36 ± 0.88 | 2.59 ± 0.85 | 2.68 ± 1.30 | ||||
| C | 4.46 ± 0.83 T,G,S | 3.63 ± 0.91 T,R | 3.80 ± 0.82 T,R | 4.01 ± 0.97 T,G | ||||
| Taurine | T | 68.7 ± 43.6 | 63.6 ± 37.8 | 71.1 ± 27.8 | 76.2 ± 34.1 | 0.008 (0.25) T > C G > C | 0.056 (0.07) | 0.338 (0.08) |
| G | 60.4 ± 21.7 | 70.9 ± 25.7 | 70.7 ± 16.4 | 70.0 ± 15.5 | ||||
| S | 53.7 ± 19.6 | 65.0 ± 20.3 | 73.8 ± 22.9 | 65.7 ± 16.5 | ||||
| C | 42.9 ± 10.9 | 42.5 ± 6.9 | 45.0 ± 13.0 | 40.2 ± 8.0 | ||||
| Total non-proteino- genic PFAAs | T | 254 ± 69 | 231 ± 68 | 252 ± 67 | 258 ± 69 | 0.467 (0.06) | 0.109 (0.05) | 0.069 (0.13) |
| G | 254 ± 41 | 260 ± 49 | 256 ± 23 | 271 ± 27 | ||||
| S | 235 ± 48 | 228 ± 45 | 255 ± 48 | 233 ± 44 | ||||
| C | 256 ± 46 | 226 ± 28 | 228 ± 34 | 210 ± 71 | ||||
| Training Phase | Rest | Exhaustion | Post Exercise | ANOVA Effects p-Value (η2), Post-Hoc | ||||
|---|---|---|---|---|---|---|---|---|
| 15th min | 30th min | Training Phase | Exercise Stage | Interaction | ||||
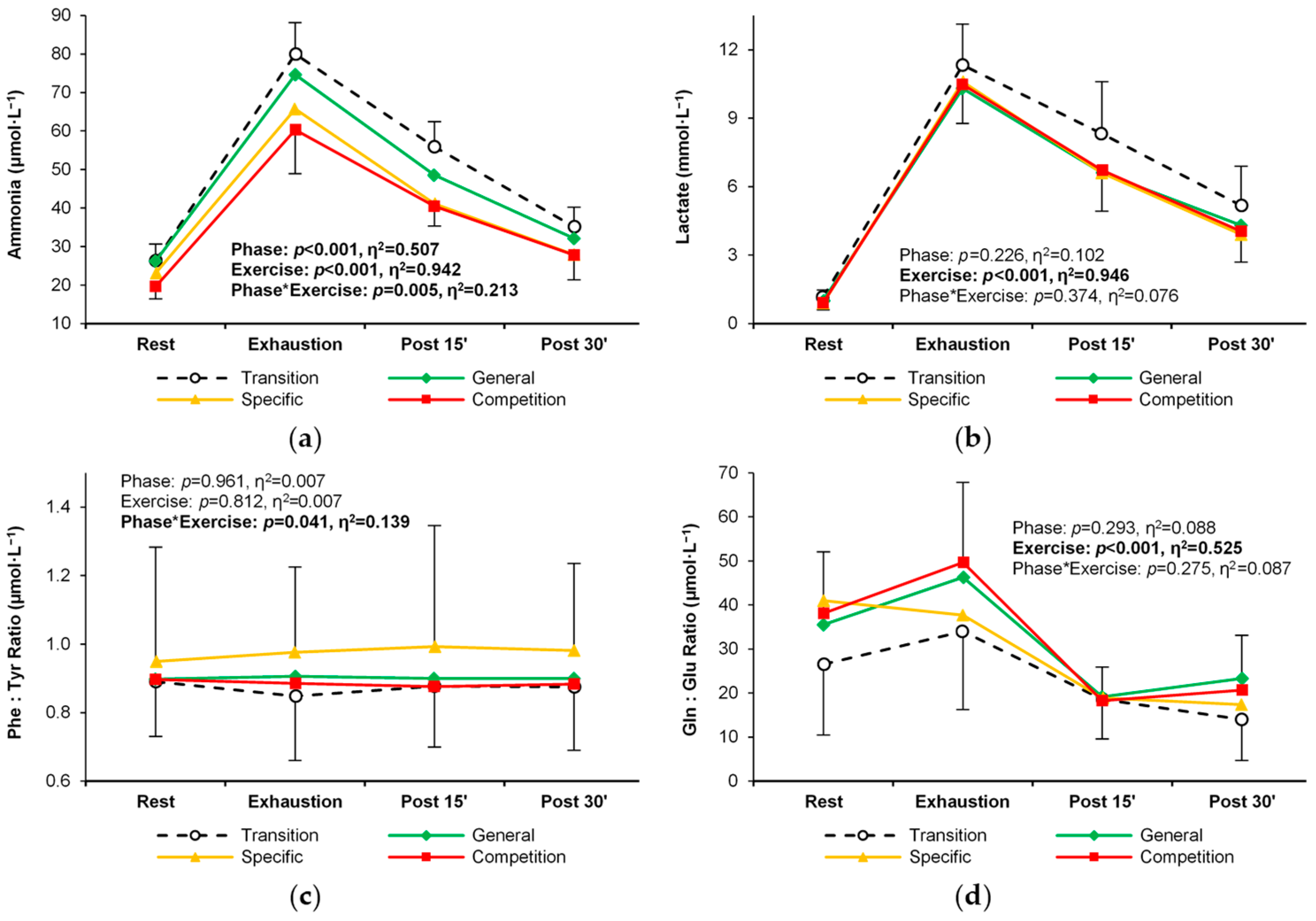
| Ammonia (μmol·L−1) | T | 26.4 ± 4.3 | 80.0 ± 8.2 R | 55.9 ± 6.5 R,E | 35.2 ± 5.0 R,E,P15 | <0.001 (0.51) T > S,C G > C | <0.001 (0.94) R < E,P15,P30 E > P15,P30 P15 > P30 | 0.005 (0.21) |
| G | 26.3 ± 5.1 | 74.6 ± 11.1 R | 48.5 ± 5.8 R,E | 32.1 ± 2.8 E,P15 | ||||
| S | 23.1 ± 3.1 | 65.7 ± 11.6 R | 41.1 ± 6.4 R,E | 27.9 ± 3.2 E,P15 | ||||
| C | 19.6 ± 3.2 | 60.3 ± 11.4 R | 40.5 ± 5.1 R,E | 27.8 ± 6.4 E,P15 | ||||
| Lactate (mmol·L−1) | T | 1.16 ± 0.32 | 11.32 ± 1.81 | 8.32 ± 2.29 | 5.17 ± 1.71 | 0.226 (0.10) | <0.001 (0.95) R < E,P15,P30 E > P15,P30 P15 > P30 | 0.374 (0.08) |
| G | 1.01 ± 0.18 | 10.30 ± 1.52 | 6.61 ± 1.77 | 4.30 ± 1.78 | ||||
| S | 0.90 ± 0.18 | 10.60 ± 2.05 | 6.60 ± 2.16 | 3.90 ± 1.63 | ||||
| C | 0.91 ± 0.31 | 10.47 ± 1.70 | 6.72 ± 1.80 | 4.04 ± 1.34 | ||||
| Phenylalani- ne: Tyrosine ratio | T | 0.89 ± 0.16 | 0.85 ± 0.19 | 0.88 ± 0.18 | 0.87 ± 0.18 | 0.961 (0.17) | 0.812 (0.01) | 0.041 (0.14) |
| G | 0.90 ± 0.12 | 0.91 ± 0.11 | 0.90 ± 0.12 | 0.90 ± 0.10 | ||||
| S | 0.95 ± 0.33 | 0.98 ± 0.25 | 0.99 ± 0.35 | 0.98 ± 0.25 | ||||
| C | 0.90 ± 0.21 | 0.89 ± 0.21 | 0.88 ± 0.18 | 0.88 ± 0.21 | ||||
| Glutamine: Glutamate ratio | T | 26.5 ± 16.0 | 34.0 ± 17.8 | 18.6 ± 9.0 | 14.0 ± 9.4 | 0.293 (0.09) | <0.001 (0.53) R < E,P15,P30 E > P15,P30 | 0.275 (0.09) |
| G | 35.5 ± 19.9 | 46.3 ± 28.9 | 19.1 ± 11.2 | 23.3 ± 20.2 | ||||
| S | 41.0 ± 14.5 | 37.7 ± 19.0 | 18.9 ± 7.8 | 17.4 ± 6.8 | ||||
| C | 38.1 ± 13.9 | 49.7 ± 18.2 | 18.3 ± 7.6 | 20.7 ± 12.4 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kusy, K.; Ciekot-Sołtysiak, M.; Matysiak, J.; Klupczyńska-Gabryszak, A.; Plewa, S.; Zarębska, E.A.; Kokot, Z.J.; Dereziński, P.; Zieliński, J. Changes in Plasma Free Amino Acid Profile in Endurance Athletes over a 9-Month Training Cycle. Metabolites 2024, 14, 353. https://doi.org/10.3390/metabo14070353
Kusy K, Ciekot-Sołtysiak M, Matysiak J, Klupczyńska-Gabryszak A, Plewa S, Zarębska EA, Kokot ZJ, Dereziński P, Zieliński J. Changes in Plasma Free Amino Acid Profile in Endurance Athletes over a 9-Month Training Cycle. Metabolites. 2024; 14(7):353. https://doi.org/10.3390/metabo14070353
Chicago/Turabian StyleKusy, Krzysztof, Monika Ciekot-Sołtysiak, Jan Matysiak, Agnieszka Klupczyńska-Gabryszak, Szymon Plewa, Ewa Anna Zarębska, Zenon J. Kokot, Paweł Dereziński, and Jacek Zieliński. 2024. "Changes in Plasma Free Amino Acid Profile in Endurance Athletes over a 9-Month Training Cycle" Metabolites 14, no. 7: 353. https://doi.org/10.3390/metabo14070353
APA StyleKusy, K., Ciekot-Sołtysiak, M., Matysiak, J., Klupczyńska-Gabryszak, A., Plewa, S., Zarębska, E. A., Kokot, Z. J., Dereziński, P., & Zieliński, J. (2024). Changes in Plasma Free Amino Acid Profile in Endurance Athletes over a 9-Month Training Cycle. Metabolites, 14(7), 353. https://doi.org/10.3390/metabo14070353