

Systematic Insights into the Relationship between the Microbiota–Gut–Brain Axis and Stroke with the Focus on Tryptophan Metabolism

Abstract
:1. Introduction
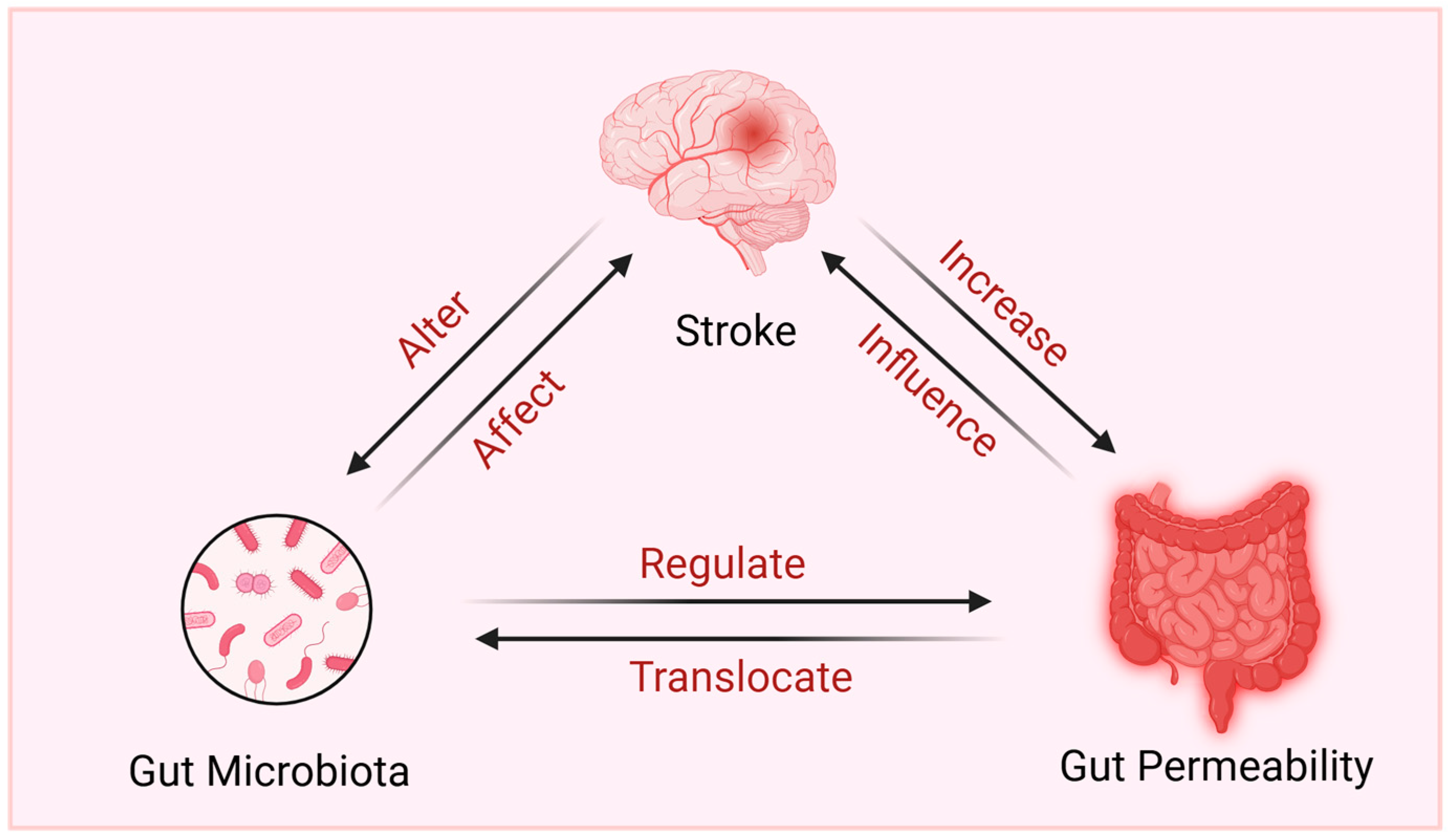
2. Microbiota–Gut–Brain Axis and Stroke
2.1. Gut Microbiota and Stroke: Epidemiological Relationships
2.2. Mechanisms of the Gut–Brain Axis in Stroke
2.2.1. The Barrier Structure of the Microbiota–Gut–Brain Axis
- Gut Barrier and Its Association with Stroke
- Blood–Brain Barrier Can Be Influenced by the Gut Microbiota
2.2.2. The Defense System of the Microbiota–Gut–Brain Axis
- The Immune System in the Gut
- The Immune System in the Brain
- Gut Microbiota Influences the Immune System
2.2.3. The Neural Network of the Microbiota–Gut–Brain Axis
- The Enteric Nervous System
- The Connections between the ENS and CNS
- Gut Microbiota Influences the Nervous System
2.2.4. The Humoral Communication of the Microbiota–Gut–Brain Axis
- Hypothalamic–Pituitary–Adrenal Axis and Its Association with Stroke
- Gut Microbiota Influences the HPA Axis
- Gut Hormones
3. Tryptophan Metabolism along the Microbiota–Gut–Brain Axis and Stroke
3.1. Tryptophan Metabolism Pathways along the Microbiota–Gut–Brain Axis
3.1.1. Kynurenine Pathway in the Gut and Brain
3.1.2. Serotonin Pathway in the Gut and Brain
3.1.3. Microbial Pathway
- The Gut Microbiota’s Role in Direct Trp Metabolism
- Gut Microbiota Influences the Kynurenine and Serotonin Pathways
3.2. Tryptophan Metabolism and Stroke
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Type of Stroke | Cohort Size | α-Diversity in Stroke | Main Changes in Gut Microbiota in Stroke | Year | Refs. |
|---|---|---|---|---|---|
| IS | 30 stroke patients and 30 controls | No difference in Ace, Chao1, Simpson, and Shannon | ↑Odoribacter, Akkermansia, Ruminococcaceae e_UCG_005, norank_p_Flavobacteriaceae, norank_p_Parcubacteria, Victivallis | 2019 | [22] |
| ↓Anaerostipes, Ruminiclostridium_5 | |||||
| IS | 31 stroke patients and 9 controls | No difference in Ace, Chao1, Simpson, Shannon, and observed OTUs | ↑Streptococcus * infantis, Prevotella * copri | 2019 | [207] |
| ↓Blautia obeum | |||||
| IS | 79 stroke patients and 98 controls | No difference in Ace, Chao1, Simpson, Shannon, and observed OTUs | ↑Proteobacteria *, Actinobacteria *, Lactobacillus *, Lactococcus *, Escherichia/Shigella *, Streptococcus *, Collinsella *, Dorea * | 2020 | [208] |
| ↓Firmicutes *, Bacteroidetes *, Faecalibacterium *, Subdoligranulum *, Eubacterium *, Roseburia *, Lachnoclostridium *, Butyricicoccus * | |||||
| IS, TIA, and HS | 349 stroke patients and 51 controls | ↓ Shannon, Simpson, and observed OTUs | ↑Proteobacteria, Escherichia/Shigella, Peptoniphilus, Ezakiella, Enterococcus | 2020 | [209] |
| ↓Firmicutes, Bacteroidetes | |||||
| IS | 20 stroke patients, 10 post-stroke, and 16 controls | ↑Ace, Chao1, and observed OTUs | ↑Actinobacteria *, Bacteroidaceae *, Bifidobacteriaceae *, Coriobacteriaceae *, Enterobacteriaceae, Weeksellaceae, Bacillaceae, Paenibaciiaceae, Brucellaceae, Xanthomnadaceae | 2020 | [210] |
| ↓Firmicutes *, Lachnospiraceae *, Ruminococcaceae*, Prevotellaceae* | |||||
| IS | 28 stroke patients and 28 controls | ↓Shannon and phylogenetic diversity | ↑Enterobacteriaceae, Ruminococcaceae, Veillonellaceae, Lachnospiraceae | 2021 | [12] |
| ↓Bacteroidaceae, Prevotellaceae | |||||
| AIS | 140 stroke patients and 92 controls | N/A | ↑Lactobacillaceae *, Enterobacteriaceae *, Porphyromonadaceae *, Akkermansia * | 2021 | [211] |
| ↓Lachnospiraceae *, Roseburia *, Bacteroides *, Faecalibacterium *, Blautia *, Anaerostipes * | |||||
| AIS and AIS with T2D | 150 stroke patients and 55 controls | N/A | ↑Enterobacteriaceae *, Proteobacteria, Gammaproteobactera, Deltaproteobacteri, Desulfovibrionaceae, Dorea *, Lactobacillus, Megasphaera | 2021 | [212] |
| ↓Lachnospira *, Coprococcus *, Prevotella, Faecalibacterium, Roseburia | |||||
| IS | 82 stroke patients and 82 controls | ↑Shannon and observed OTUs | ↑Lactobacillaceae *, Enterococcaceae *, Streptococcaceae *, Enterobacteriaceae * | 2022 | [25] |
| ↓Ruminococcaceae, Lachnospiraceae | |||||
| ICH | 31 stroke patients and 31 controls | No difference in Chao1, Shannon, Simpson, and observed OTUs | ↑Bacteroidetes, Streptococcus, Bifidobacterium, Akkermansia, Lactobacillus, Enterococcus | 2022 | [213] |
| ↓Firmicutes, Faecalibacterium, Clostridium, Prevotella, Gemmiger, Blautia | |||||
| AIS and AIS with PHS | 17 stroke patients and 10 controls | ↑Chao1 No difference in Ace, Shannon, and Simpson | ↑Verrucomicrobia *, Synergistetes *, Actinobacteria, Proteobacteria, Akkermansia *, Olsenella *, Escherichia_Shigella, Bifidobacterium, Ruminococcaceae_UCG_014 | 2022 | [214] |
| ↓Firmicutes, Bacteroidetes, Megamonas *, Prevotella_9, Agathobacter, Faecalibacterium | |||||
| CS | 30 stroke patients and 33 controls | ↑Chao1, Shannon, Ace, and observed OTUs | ↑Enterobacteriaceae *, Streptococcaceae *, Lactobacillaceae *, Escherichia–Shigella *, Streptococcus *, Lactobacillus *, Klebsilla | 2022 | [24] |
| ↓Veillonellaceae *, Faecalibacterium *, Dialister, Roseburia | |||||
| HS and IS | 20 stroke patients and 20 controls | ↓Chao, Ace, Shannon, Simpson, and observed OTUs | ↑Fusobacteriota *, Desulfobacterota *, Akkermansiaceae, Fusobacteriota, Ruminococcaceae, Oscillospirales | 2023 | [215] |
| ↓Acidobacteriota *, Bacteroides | |||||
| AIS | 90 stroke patients and 60 controls | N/A | ↑Actinobacteriota, Bacteroides, Parabacteroides Faecalibacterium | 2023 | [23] |
| ↓Spirochaetes, Faecallbacterium, Prevotella, Roseburia, Lachnospira | |||||
| IS | 10 stroke patients and 21 controls | ↓Chao1 No difference in Shannon and Simpson | ↑Proteobacteria *, Verrucomicrobiaceae *, Bacteroidaceae *, Streptococcaceae *, Rikenellaceae *, Enterobacteriaceae *, Christensenellacea *, Phascolarctobacterium, Alistipes, Sutterella, Akkermansia, Bacteroides | 2023 | [196] |
| ↓Coriobacteriaceae *, Actinobacteria, Clostridiaceae, Ruminococcaceae, Anaerostipes *, Clostridiales * | |||||
| AIS and AIS with HHTN | 200 stroke patients and 90 controls | No difference in Ace and Shannon | ↑Streptococcaceae *, Lactobacillaceae *, Marinifilaceae *, Lactobacillus *, Veillonella *, Bacteroides, Bifidobacterium, Klebsiella | 2023 | [216] |
| ↓Escherichia-Shigella *, Streptococcus *, Anaerostipes *, Butyricicoccus *, Blautia, Faecalibacterium |
References
- Martin, S.S.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Barone Gibbs, B.; Beaton, A.Z.; Boehme, A.K.; et al. 2024 Heart Disease and Stroke Statistics: A Report of US and Global Data From the American Heart Association. Circulation 2024, 149, e347–e913. [Google Scholar] [CrossRef]
- Xie, X.; Wang, L.; Dong, S.; Ge, S.; Zhu, T. Immune Regulation of the Gut-Brain Axis and Lung-Brain Axis Involved in Ischemic Stroke. Neural Regen. Res. 2024, 19, 519–528. [Google Scholar] [CrossRef]
- Tsivgoulis, G.; Katsanos, A.H.; Sandset, E.C.; Turc, G.; Nguyen, T.N.; Bivard, A.; Fischer, U.; Khatri, P. Thrombolysis for Acute Ischaemic Stroke: Current Status and Future Perspectives. Lancet Neurol. 2023, 22, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Rost, N.S.; Brodtmann, A.; Pase, M.P.; Van Veluw, S.J.; Biffi, A.; Duering, M.; Hinman, J.D.; Dichgans, M. Post-Stroke Cognitive Impairment and Dementia. Circ. Res. 2022, 130, 1252–1271. [Google Scholar] [CrossRef] [PubMed]
- Schöttke, H.; Giabbiconi, C.-M. Post-Stroke Depression and Post-Stroke Anxiety: Prevalence and Predictors. Int. Psychogeriatr. 2015, 27, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, S. Communication Impairment in Patients Following Stroke. Nurs. Stand. 2012, 26, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xiong, Q.; Stary, C.M.; Mahgoub, O.K.; Ye, Y.; Gu, L.; Xiong, X.; Zhu, S. Bidirectional Gut-Brain-Microbiota Axis as a Potential Link between Inflammatory Bowel Disease and Ischemic Stroke. J. Neuroinflamm. 2018, 15, 339. [Google Scholar] [CrossRef]
- Sinagra, E.; Pellegatta, G.; Guarnotta, V.; Maida, M.; Rossi, F.; Conoscenti, G.; Pallio, S.; Alloro, R.; Raimondo, D.; Pace, F.; et al. Microbiota Gut–Brain Axis in Ischemic Stroke: A Narrative Review with a Focus about the Relationship with Inflammatory Bowel Disease. Life 2021, 11, 715. [Google Scholar] [CrossRef]
- Tuz, A.A.; Hasenberg, A.; Hermann, D.M.; Gunzer, M.; Singh, V. Ischemic Stroke and Concomitant Gastrointestinal Complications- a Fatal Combination for Patient Recovery. Front. Immunol. 2022, 13, 1037330. [Google Scholar] [CrossRef]
- Song, J.; Chen, W.; Ye, W. Stroke and the Risk of Gastrointestinal Disorders: A Mendelian Randomization Study. Front. Neurol. 2023, 14, 1131250. [Google Scholar] [CrossRef]
- Roth, W.H.; Cai, A.; Zhang, C.; Chen, M.L.; Merkler, A.E.; Kamel, H. Gastrointestinal Disorders and Risk of First-Ever Ischemic Stroke. Upd. Int. Car. 2020, 51, 3577–3583. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Gao, X.; Xia, G.; Chen, M.; Zeng, N.; Wang, S.; You, C.; Tian, X.; Di, H.; Tang, W.; et al. Rapid Gut Dysbiosis Induced by Stroke Exacerbates Brain Infarction in Turn. Gut 2021, 70, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. 2015, 28, 203. [Google Scholar]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Mu, C.; Farzi, A.; Zhu, W. Tryptophan Metabolism: A Link between the Gut Microbiota and Brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Li, G.; Zheng, Q.; Gu, X.; Shi, Q.; Su, Y.; Chu, Q.; Yuan, X.; Bao, Z.; Lu, J.; et al. Tryptophan Metabolism in Health and Disease. Cell Metab. 2023, 35, 1304–1326. [Google Scholar] [CrossRef]
- Wigner, P.; Saluk-Bijak, J.; Synowiec, E.; Miller, E.; Sliwinski, T.; Cichon, N.; Bijak, M. Variation of Genes Encoding Tryptophan Catabolites Pathway Enzymes in Stroke. J. Clin. Med. 2019, 8, 2133. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.; Hess, M.; Weimer, B.C. Microbial-Derived Tryptophan Metabolites and Their Role in Neurological Disease: Anthranilic Acid and Anthranilic Acid Derivatives. Microorganisms 2023, 11, 1825. [Google Scholar] [CrossRef]
- Boros, F.A.; Maszlag-Török, R.; Szűcs, M.; Annus, Á.; Klivényi, P.; Vécsei, L. Relationships of Ischemic Stroke Occurrence and Outcome with Gene Variants Encoding Enzymes of Tryptophan Metabolism. Biomedicines 2021, 9, 1441. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Gut Bacteria in Health and Disease. Gastroenterol. Hepatol. 2013, 9, 560. [Google Scholar]
- MetaHIT Consortium; Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, X.; Sun, C.; Wu, X.; Lu, M.; Si, Y.; Ye, X.; Wang, T.; Yu, X.; Zhao, X.; et al. Change of Intestinal Microbiota in Cerebral Ischemic Stroke Patients. BMC Microbiol. 2019, 19, 191. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Li, L.; Liu, L.; Ding, B.; Yang, Y.; He, F.; Zhang, L.; Wu, Z. Ischemic Stroke and Dysbiosis of Gut Microbiota: Changes to LPS Levels and Effects on Functional Outcomes. Altern. Ther. Health Med. 2023, 29, 284. [Google Scholar] [PubMed]
- Zheng, Q.; Chen, Y.; Zhai, Y.; Meng, L.; Liu, H.; Tian, H.; Feng, R.; Wang, J.; Zhang, R.; Sun, K.; et al. Gut Dysbiosis Is Associated with the Severity of Cryptogenic Stroke and Enhanced Systemic Inflammatory Response. Front. Immunol. 2022, 13, 836820. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, H.; Wang, X.; Tian, M.; Tian, Y.; Li, Q.; Liang, C.; Peng, X.; Ding, J.; Wu, X.; et al. Combination Effect between Gut Microbiota and Traditional Potentially Modifiable Risk Factors for First-Ever Ischemic Stroke in Tujia, Miao and Han Populations in China. Front. Mol. Neurosci. 2022, 15, 922399. [Google Scholar] [CrossRef] [PubMed]
- Aburto, M.R.; Cryan, J.F. Gastrointestinal and Brain Barriers: Unlocking Gates of Communication across the Microbiota–Gut–Brain Axis. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 222–247. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Stanley, D.; Mason, L.J.; Mackin, K.E.; Srikhanta, Y.N.; Lyras, D.; Prakash, M.D.; Nurgali, K.; Venegas, A.; Hill, M.D.; Moore, R.J.; et al. Translocation and Dissemination of Commensal Bacteria in Post-Stroke Infection. Nat. Med. 2016, 22, 1277–1284. [Google Scholar] [CrossRef]
- Houlden, A.; Goldrick, M.; Brough, D.; Vizi, E.S.; Lénárt, N.; Martinecz, B.; Roberts, I.S.; Denes, A. Brain Injury Induces Specific Changes in the Caecal Microbiota of Mice via Altered Autonomic Activity and Mucoprotein Production. Brain Behav. Immun. 2016, 57, 10–20. [Google Scholar] [CrossRef]
- Amasheh, S.; Fromm, M.; Günzel, D. Claudins of Intestine and Nephron—A Correlation of Molecular Tight Junction Structure and Barrier Function. Acta Physiol. 2011, 201, 133–140. [Google Scholar] [CrossRef]
- Prame Kumar, K.; McKay, L.D.; Nguyen, H.; Kaur, J.; Wilson, J.L.; Suthya, A.R.; McKeown, S.J.; Abud, H.E.; Wong, C.H.Y. Sympathetic-Mediated Intestinal Cell Death Contributes to Gut Barrier Impairment after Stroke. Transl. Stroke Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Hu, Y.; Zhu, N.; Gu, W.; Long, G.; Tao, E.; Fang, M.; Jiang, M. Exploratory Investigation of Intestinal Structure and Function after Stroke in Mice. Mediat. Inflamm. 2021, 2021, 1315797. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Larsson, J.M.H.; Hansson, G.C. The Two Mucus Layers of Colon Are Organized by the MUC2 Mucin, Whereas the Outer Layer Is a Legislator of Host–Microbial Interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 4659–4665. [Google Scholar] [CrossRef] [PubMed]
- Arike, L.; Seiman, A.; Van Der Post, S.; Rodriguez Piñeiro, A.M.; Ermund, A.; Schütte, A.; Bäckhed, F.; Johansson, M.E.V.; Hansson, G.C. Protein Turnover in Epithelial Cells and Mucus along the Gastrointestinal Tract Is Coordinated by the Spatial Location and Microbiota. Cell Rep. 2020, 30, 1077–1087.e3. [Google Scholar] [CrossRef]
- Bevins, C.L.; Salzman, N.H. Paneth Cells, Antimicrobial Peptides and Maintenance of Intestinal Homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef]
- Yu, L.C.-H. Host-Microbial Interactions and Regulation of Intestinal Epithelial Barrier Function: From Physiology to Pathology. World J. Gastrointest. Pathophysiol. 2012, 3, 27. [Google Scholar] [CrossRef]
- Lawther, B.K.; Kumar, S.; Krovvidi, H. Blood–Brain Barrier. Contin. Educ. Anaesth. Crit. Care Pain 2011, 11, 128–132. [Google Scholar] [CrossRef]
- Kim, K.-A.; Kim, D.; Kim, J.-H.; Shin, Y.-J.; Kim, E.-S.; Akram, M.; Kim, E.-H.; Majid, A.; Baek, S.-H.; Bae, O.-N. Autophagy-Mediated Occludin Degradation Contributes to Blood–Brain Barrier Disruption during Ischemia in bEnd.3 Brain Endothelial Cells and Rat Ischemic Stroke Models. Fluids Barriers CNS 2020, 17, 21. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The Gut Microbiota Influences Blood-Brain Barrier Permeability in Mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef]
- Fröhlich, E.E.; Farzi, A.; Mayerhofer, R.; Reichmann, F.; Jačan, A.; Wagner, B.; Zinser, E.; Bordag, N.; Magnes, C.; Fröhlich, E.; et al. Cognitive Impairment by Antibiotic-Induced Gut Dysbiosis: Analysis of Gut Microbiota-Brain Communication. Brain Behav. Immun. 2016, 56, 140–155. [Google Scholar] [CrossRef]
- Rustia, A.J.; Paterson, J.S.; Best, G.; Sokoya, E.M. Microbial Disruption in the Gut Promotes Cerebral Endothelial Dysfunction. Physiol. Rep. 2021, 9, e15100. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.W.; Phillips, S.C.; Ganesh, B.P.; Petrosino, J.F.; Durgan, D.J.; Bryan, R.M. The Gut Microbiome Contributes to Blood-brain Barrier Disruption in Spontaneously Hypertensive Stroke Prone Rats. FASEB J. 2021, 35, e21201. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, W.; Wu, Q.; Gao, X.; Li, J.; Tan, C.; Zhou, H.; Zhu, J.; He, Y.; Yin, J. Fecal Transplantation from Db/Db Mice Treated with Sodium Butyrate Attenuates Ischemic Stroke Injury. Microbiol. Spectr. 2021, 9, e00042-21. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; d’Aigle, J.; Atadja, L.; Quaicoe, V.; Honarpisheh, P.; Ganesh, B.P.; Hassan, A.; Graf, J.; Petrosino, J.; Putluri, N.; et al. Gut Microbiota–Derived Short-Chain Fatty Acids Promote Poststroke Recovery in Aged Mice. Circ. Res. 2020, 127, 453–465. [Google Scholar] [CrossRef]
- Peh, A.; Dinakis, E.; Jama, H.; Anderson, D.; Creek, D.J.; Zheng, G.; De Veer, M.; Mackay, C.R.; Zheng, T.; Kemp-Harper, B.K.; et al. Dietary Fibre Reverses Adverse Post-Stroke Outcomes in Mice via Short-Chain Fatty Acids and Its Sensing Receptors GPR41, GPR43 and GPR109A. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wang, H.-B.; Wang, P.-Y.; Wang, X.; Wan, Y.-L.; Liu, Y.-C. Butyrate Enhances Intestinal Epithelial Barrier Function via Up-Regulation of Tight Junction Protein Claudin-1 Transcription. Digest. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, W.; Gao, X.; Zhu, J.; Li, J.; Wu, Q.; Tan, C.; Zhou, H.; He, Y.; Yin, J. Modulation of the Gut Microbiota of Type 2 Diabetic Mice by Sodium Butyrate Attenuates Ischemic Stroke Injury. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Zheng, L.; Kelly, C.J.; Battista, K.D.; Schaefer, R.; Lanis, J.M.; Alexeev, E.E.; Wang, R.X.; Onyiah, J.C.; Kominsky, D.J.; Colgan, S.P. Microbial-Derived Butyrate Promotes Epithelial Barrier Function through IL-10 Receptor–Dependent Repression of Claudin-2. J. Immunol. 2017, 199, 2976–2984. [Google Scholar] [CrossRef] [PubMed]
- Powell, N.; Walker, M.M.; Talley, N.J. The Mucosal Immune System: Master Regulator of Bidirectional Gut–Brain Communications. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 143–159. [Google Scholar] [CrossRef]
- Mowat, A. McI. Anatomical Basis of Tolerance and Immunity to Intestinal Antigens. Nat. Rev. Immunol. 2003, 3, 331–341. [Google Scholar] [CrossRef]
- Song, C.; Chai, Z.; Chen, S.; Zhang, H.; Zhang, X.; Zhou, Y. Intestinal Mucus Components and Secretion Mechanisms: What We Do and Do Not Know. Exp. Mol. Med. 2023, 55, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Honarpisheh, P.; Bryan, R.M.; McCullough, L.D. Aging Microbiota-Gut-Brain Axis in Stroke Risk and Outcome. Circ. Res. 2022, 130, 1112–1144. [Google Scholar] [CrossRef] [PubMed]
- Kasarello, K.; Cudnoch-Jedrzejewska, A.; Czarzasta, K. Communication of Gut Microbiota and Brain via Immune and Neuroendocrine Signaling. Front. Microbiol. 2023, 14, 1118529. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhu, L.; Song, W.; Zhang, M.; Teng, L.; Wu, M. Crosstalk between the Gut and Brain in Ischemic Stroke: Mechanistic Insights and Therapeutic Options. Mediat. Inflamm. 2022, 2022, 6508046. [Google Scholar] [CrossRef]
- Eletto, D.; Mentucci, F.; Voli, A.; Petrella, A.; Porta, A.; Tosco, A. Helicobacter Pylori Pathogen-Associated Molecular Patterns: Friends or Foes? Int. J. Mol. Sci. 2022, 23, 3531. [Google Scholar] [CrossRef]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw. 2018, 18, e27. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, S.; Hafez, A.; Rezai Jahromi, B.; Kinfe, T.; Lamprecht, A.; Niemelä, M.; Muhammad, S. Role of Damage Associated Molecular Pattern Molecules (DAMPs) in Aneurysmal Subarachnoid Hemorrhage (aSAH). Int. J. Mol. Sci. 2018, 19, 2035. [Google Scholar] [CrossRef]
- Fadakar, K.; Dadkhahfar, S.; Esmaeili, A.; Rezaei, N. The Role of Toll-like Receptors (TLRs) in Stroke. Rev. Neurosci. 2014, 25, 699–712. [Google Scholar] [CrossRef]
- Logsdon, A.F.; Erickson, M.A.; Rhea, E.M.; Salameh, T.S.; Banks, W.A. Gut Reactions: How the Blood–Brain Barrier Connects the Microbiome and the Brain. Exp. Biol. Med. 2018, 243, 159–165. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Peng, J.; Gerner, S.T.; Yin, S.; Jiang, Y. Gut Microbiota-Brain Interaction: An Emerging Immunotherapy for Traumatic Brain Injury. Exp. Neurol. 2021, 337, 113585. [Google Scholar] [CrossRef]
- Brea, D.; Poon, C.; Benakis, C.; Lubitz, G.; Murphy, M.; Iadecola, C.; Anrather, J. Stroke Affects Intestinal Immune Cell Trafficking to the Central Nervous System. Brain Behav. Immun. 2021, 96, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Waisman, A.; Liblau, R.S.; Becher, B. Innate and Adaptive Immune Responses in the CNS. Lancet Neurol. 2015, 14, 945–955. [Google Scholar] [CrossRef]
- Reemst, K.; Noctor, S.C.; Lucassen, P.J.; Hol, E.M. The Indispensable Roles of Microglia and Astrocytes during Brain Development. Front. Hum. Neurosci. 2016, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, G. CNS-Peripheral Immune Interactions in Hemorrhagic Stroke. J. Cereb. Blood Flow Metab. 2023, 43, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kawabori, M.; Yenari, M.A. Innate Inflammatory Responses in Stroke: Mechanisms and Potential Therapeutic Targets. Curr. Med. Chem. 2014, 21, 2076–2097. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, S.; Cunningham, M.W.; Pabbidi, M.R.; Wang, S.; Booz, G.W.; Fan, F. Targeting Vascular Inflammation in Ischemic Stroke: Recent Developments on Novel Immunomodulatory Approaches. Eur. J. Pharmacol. 2018, 833, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Roth, S.; Llovera, G.; Sadler, R.; Garzetti, D.; Stecher, B.; Dichgans, M.; Liesz, A. Microbiota Dysbiosis Controls the Neuroinflammatory Response after Stroke. J. Neurosci. 2016, 36, 7428–7440. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C.; Buckwalter, M.S.; Anrather, J. Immune Responses to Stroke: Mechanisms, Modulation, and Therapeutic Potential. J. Clin. Investig. 2020, 130, 2777–2788. [Google Scholar] [CrossRef]
- Wu, S.; Tabassum, S.; Payne, C.T.; Hu, H.; Gusdon, A.M.; Choi, H.A.; Ren, X.S. Updates of the Role of B-Cells in Ischemic Stroke. Front. Cell. Neurosci. 2024, 18, 1340756. [Google Scholar] [CrossRef]
- Prass, K.; Meisel, C.; Höflich, C.; Braun, J.; Halle, E.; Wolf, T.; Ruscher, K.; Victorov, I.V.; Priller, J.; Dirnagl, U.; et al. Stroke-Induced Immunodeficiency Promotes Spontaneous Bacterial Infections and Is Mediated by Sympathetic Activation Reversal by Poststroke T Helper Cell Type 1–like Immunostimulation. J. Exp. Med. 2003, 198, 725–736. [Google Scholar] [CrossRef]
- Pu, B.; Zhu, H.; Wei, L.; Gu, L.; Zhang, S.; Jian, Z.; Xiong, X. The Involvement of Immune Cells Between Ischemic Stroke and Gut Microbiota. Transl. Stroke Res. 2024, 15, 498–517. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Sadler, R.; Heindl, S.; Llovera, G.; Roth, S.; Benakis, C.; Liesz, A. The Gut Microbiome Primes a Cerebroprotective Immune Response after Stroke. J. Cereb. Blood Flow Metab. 2018, 38, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.T.; Boudaud, M.; Muller, A.; Macpherson, A.J.; Desai, M.S. Maternal Diet and Gut Microbiome Composition Modulate Early-life Immune Development. EMBO Mol. Med. 2023, 15, e17241. [Google Scholar] [CrossRef]
- Zeng, J.; Yang, K.; Nie, H.; Yuan, L.; Wang, S.; Zeng, L.; Ge, A.; Ge, J. The Mechanism of Intestinal Microbiota Regulating Immunity and Inflammation in Ischemic Stroke and the Role of Natural Botanical Active Ingredients in Regulating Intestinal Microbiota: A Review. Biomed. Pharmacother. 2023, 157, 114026. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System and Neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, K.A.; Mawe, G.M. The Enteric Nervous System. Physiol. Rev. 2023, 103, 1487–1564. [Google Scholar] [CrossRef] [PubMed]
- Spencer, N.J.; Hu, H. Enteric Nervous System: Sensory Transduction, Neural Circuits and Gastrointestinal Motility. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Jarret, A.; Jackson, R.; Duizer, C.; Healy, M.E.; Zhao, J.; Rone, J.M.; Bielecki, P.; Sefik, E.; Roulis, M.; Rice, T.; et al. Enteric Nervous System-Derived IL-18 Orchestrates Mucosal Barrier Immunity. Cell 2020, 180, 50–63.e12. [Google Scholar] [CrossRef]
- Muller, P.A.; Koscsó, B.; Rajani, G.M.; Stevanovic, K.; Berres, M.-L.; Hashimoto, D.; Mortha, A.; Leboeuf, M.; Li, X.-M.; Mucida, D.; et al. Crosstalk between Muscularis Macrophages and Enteric Neurons Regulates Gastrointestinal Motility. Cell 2014, 158, 300–313. [Google Scholar] [CrossRef]
- Viola, M.F.; Chavero-Pieres, M.; Modave, E.; Delfini, M.; Stakenborg, N.; Estévez, M.C.; Fabre, N.; Appeltans, I.; Martens, T.; Vandereyken, K.; et al. Dedicated Macrophages Organize and Maintain the Enteric Nervous System. Nature 2023, 618, 818–826. [Google Scholar] [CrossRef]
- Cheng, X.; Boza-Serrano, A.; Turesson, M.F.; Deierborg, T.; Ekblad, E.; Voss, U. Galectin-3 Causes Enteric Neuronal Loss in Mice after Left Sided Permanent Middle Cerebral Artery Occlusion, a Model of Stroke. Sci. Rep. 2016, 6, 32893. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Svensson, M.; Yang, Y.; Deierborg, T.; Ekblad, E.; Voss, U. Focal, but Not Global, Cerebral Ischaemia Causes Loss of Myenteric Neurons and Upregulation of Vasoactive Intestinal Peptide in Mouse Ileum. Int. J. Exp. Path. 2018, 99, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.P.; Wilson, J.L.; Nguyen, H.; McKay, L.D.; Wen, S.W.; Sepehrizadeh, T.; De Veer, M.; Rajasekhar, P.; Carbone, S.E.; Hickey, M.J.; et al. Stroke Alters the Function of Enteric Neurons to Impair Smooth Muscle Relaxation and Dysregulates Gut Transit. JAHA 2024, 13, e033279. [Google Scholar] [CrossRef]
- Goyal, R.K. The Enteric Nervous System. N. Engl. J. Med. 1996, 334, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Li, J.; Zhou, Q.; Li, S.; Xie, C.; Niu, L.; Ma, J.; Li, C. Vagus Nerve Stimulation Alleviated Cerebral Ischemia and Reperfusion Injury in Rats by Inhibiting Pyroptosis via A7 Nicotinic Acetylcholine Receptor. Cell Death Discov. 2022, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Wang, S.-R.; Ma, J.-X.; Yu, L.-H.; Jia, G.-W. Vagus Nerve Stimulation Is a Potential Treatment for Ischemic Stroke. Neural Regener. Res. 2023, 18, 825. [Google Scholar] [CrossRef]
- Muller, P.A.; Schneeberger, M.; Matheis, F.; Wang, P.; Kerner, Z.; Ilanges, A.; Pellegrino, K.; Del Mármol, J.; Castro, T.B.R.; Furuichi, M.; et al. Microbiota Modulate Sympathetic Neurons via a Gut–Brain Circuit. Nature 2020, 583, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Goehler, L.E.; Gaykema, R.P.A.; Opitz, N.; Reddaway, R.; Badr, N.; Lyte, M. Activation in Vagal Afferents and Central Autonomic Pathways: Early Responses to Intestinal Infection with Campylobacter Jejuni. Brain Behav. Immun. 2005, 19, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, F.A.; Keenan, C.M.; Wallace, L.E.; Woods, C.; Cavin, J.-B.; Flockton, A.R.; Macklin, W.B.; Belkind-Gerson, J.; Hirota, S.A.; Sharkey, K.A. Intestinal Microbiota Shapes Gut Physiology and Regulates Enteric Neurons and Glia. Microbiome 2021, 9, 210. [Google Scholar] [CrossRef]
- Yan, Y.; Ramanan, D.; Rozenberg, M.; McGovern, K.; Rastelli, D.; Vijaykumar, B.; Yaghi, O.; Voisin, T.; Mosaheb, M.; Chiu, I.; et al. Interleukin-6 Produced by Enteric Neurons Regulates the Number and Phenotype of Microbe-Responsive Regulatory T Cells in the Gut. Immunity 2021, 54, 499–513.e5. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of Neurotransmitters by the Gut Microbiota and Effects on Cognition in Neurological Disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef] [PubMed]
- Burford, N.; Webster, N.; Cruz-Topete, D. Hypothalamic-Pituitary-Adrenal Axis Modulation of Glucocorticoids in the Cardiovascular System. Int. J. Mol. Sci. 2017, 18, 2150. [Google Scholar] [CrossRef] [PubMed]
- Fassbender, K.; Schmidt, R.; Mössner, R.; Daffertshofer, M.; Hennerici, M. Pattern of Activation of the Hypothalamic-Pituitary-Adrenal Axis in Acute Stroke. Relation to Acute Confusional State, Extent of Brain Damage, and Clinical Outcome. Stroke 1994, 25, 1105–1108. [Google Scholar] [CrossRef]
- Barugh, A.J.; Gray, P.; Shenkin, S.D.; MacLullich, A.M.J.; Mead, G.E. Cortisol Levels and the Severity and Outcomes of Acute Stroke: A Systematic Review. J. Neurol. 2014, 261, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Sugo, N.; Hurn, P.D.; Morahan, M.B.; Hattori, K.; Traystman, R.J.; DeVries, A.C. Social Stress Exacerbates Focal Cerebral Ischemia in Mice. Stroke 2002, 33, 1660–1664. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, E.S.; Chen, P.R.; Kim, E. Dysregulated Hypothalamic–Pituitary–Adrenal Axis Is Associated With Increased Inflammation and Worse Outcomes After Ischemic Stroke in Diabetic Mice. Front. Immunol. 2022, 13, 864858. [Google Scholar] [CrossRef] [PubMed]
- Mracsko, E.; Liesz, A.; Karcher, S.; Zorn, M.; Bari, F.; Veltkamp, R. Differential Effects of Sympathetic Nervous System and Hypothalamic–Pituitary–Adrenal Axis on Systemic Immune Cells after Severe Experimental Stroke. Brain Behav. Immun. 2014, 41, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Sun, W.; Miao, J.; Liang, W.; Qiu, X.; Lan, Y.; Pan, C.; Li, G.; Zhao, X.; et al. Higher Concentration of Adrenocorticotropic Hormone Predicts Post-Stroke Depression. Clin. Interv. Aging 2022, 17, 417–427. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.; Kubo, C.; Koga, Y. Postnatal Microbial Colonization Programs the Hypothalamic–Pituitary–Adrenal System for Stress Response in Mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef]
- Vodička, M.; Ergang, P.; Hrnčíř, T.; Mikulecká, A.; Kvapilová, P.; Vagnerová, K.; Šestáková, B.; Fajstová, A.; Hermanová, P.; Hudcovic, T.; et al. Microbiota Affects the Expression of Genes Involved in HPA Axis Regulation and Local Metabolism of Glucocorticoids in Chronic Psychosocial Stress. Brain Behav. Immun. 2018, 73, 615–624. [Google Scholar] [CrossRef]
- Rehfeld, J.F. A Centenary of Gastrointestinal Endocrinology. Horm. Metab. Res. 2004, 36, 735–741. [Google Scholar] [CrossRef]
- Furness, J.B.; Rivera, L.R.; Cho, H.-J.; Bravo, D.M.; Callaghan, B. The Gut as a Sensory Organ. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-J.; Li, J.-N.; Nie, Y.-Z. Gut Hormones in Microbiota-Gut-Brain Cross-Talk. Chin. Med. J. 2020, 133, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Sternini, C.; De Giorgio, R.; Greenwood-Van Meerveld, B. Enteroendocrine Cells: A Review of Their Role in Brain–Gut Communication. Neurogastroenterol. Motil. 2016, 28, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Bogunovic, M.; Davé, S.H.; Tilstra, J.S.; Chang, D.T.W.; Harpaz, N.; Xiong, H.; Mayer, L.F.; Plevy, S.E. Enteroendocrine Cells Express Functional Toll-like Receptors. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 292, G1770–G1783. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, A.; Matsuda, K.; Kuwahara, Y.; Asano, S.; Inui, T.; Marunaka, Y. Microbiota-Gut-Brain Axis: Enteroendocrine Cells and the Enteric Nervous System Form an Interface between the Microbiota and the Central Nervous System. Biomed. Res. 2020, 41, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Comai, S.; Bertazzo, A.; Brughera, M.; Crotti, S. Tryptophan in Health and Disease. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2020; Volume 95, pp. 165–218. ISBN 978-0-12-821165-6. [Google Scholar]
- Yeung, A.W.S.; Terentis, A.C.; King, N.J.C.; Thomas, S.R. Role of Indoleamine 2,3-Dioxygenase in Health and Disease. Clin. Sci. 2015, 129, 601–672. [Google Scholar] [CrossRef]
- Huang, Y.-S.; Ogbechi, J.; Clanchy, F.I.; Williams, R.O.; Stone, T.W. IDO and Kynurenine Metabolites in Peripheral and CNS Disorders. Front. Immunol. 2020, 11, 388. [Google Scholar] [CrossRef]
- Prendergast, G.C.; Metz, R.; Muller, A.J.; Merlo, L.M.F.; Mandik-Nayak, L. IDO2 in Immunomodulation and Autoimmune Disease. Front. Immunol. 2014, 5, 585. [Google Scholar] [CrossRef]
- Roth, W.; Zadeh, K.; Vekariya, R.; Ge, Y.; Mohamadzadeh, M. Tryptophan Metabolism and Gut-Brain Homeostasis. Int. J. Mol. Sci. 2021, 22, 2973. [Google Scholar] [CrossRef]
- Lamas, B.; Natividad, J.M.; Sokol, H. Aryl Hydrocarbon Receptor and Intestinal Immunity. Mucosal Immunol. 2018, 11, 1024–1038. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, V.; Quintana, F.J. The Aryl Hydrocarbon Receptor: An Environmental Sensor Integrating Immune Responses in Health and Disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef]
- Mrštná, K.; Krčmová, L.K.; Švec, F. Advances in Kynurenine Analysis. Clin. Chim. Acta 2023, 547, 117441. [Google Scholar] [CrossRef] [PubMed]
- Beamer, C.A.; Shepherd, D.M. Role of the Aryl Hydrocarbon Receptor (AhR) in Lung Inflammation. Semin. Immunopathol. 2013, 35, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Lőrinczi, B.; Szatmári, I. KYNA Derivatives with Modified Skeleton; Hydroxyquinolines with Potential Neuroprotective Effect. Int. J. Mol. Sci. 2021, 22, 11935. [Google Scholar] [CrossRef] [PubMed]
- Vécsei, L.; Szalárdy, L.; Fülöp, F.; Toldi, J. Kynurenines in the CNS: Recent Advances and New Questions. Nat. Rev. Drug Discov. 2013, 12, 64–82. [Google Scholar] [CrossRef]
- Lopez, A.; Alegre, E.; Lemaoult, J.; Carosella, E.; Gonzalez, A. Regulatory Role of Tryptophan Degradation Pathway in HLA-G Expression by Human Monocyte-Derived Dendritic Cells. Mol. Immunol. 2006, 43, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Weber, W.P.; Feder-Mengus, C.; Chiarugi, A.; Rosenthal, R.; Reschner, A.; Schumacher, R.; Zajac, P.; Misteli, H.; Frey, D.M.; Oertli, D.; et al. Differential Effects of the Tryptophan Metabolite3-hydroxyanthranilic Acid on the Proliferation of Human CD8 + T Cells Induced by TCR Triggering or Homeostatic Cytokines. Eur. J. Immunol. 2006, 36, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Harvey-Girard, E.; Dunn, R.J. Excitatory Amino Acid Receptors of the Electrosensory System: The NR1/NR2B N-Methyl-d-Aspartate Receptor. J. Neurophysiol. 2003, 89, 822–832. [Google Scholar] [CrossRef]
- Schwarcz, R.; Stone, T.W. The Kynurenine Pathway and the Brain: Challenges, Controversies and Promises. Neuropharmacology 2017, 112, 237–247. [Google Scholar] [CrossRef]
- Pierozan, P.; Biasibetti, H.; Schmitz, F.; Ávila, H.; Parisi, M.M.; Barbe-Tuana, F.; Wyse, A.T.S.; Pessoa-Pureur, R. Quinolinic Acid Neurotoxicity: Differential Roles of Astrocytes and Microglia via FGF-2-Mediated Signaling in Redox-Linked Cytoskeletal Changes. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 3001–3014. [Google Scholar] [CrossRef] [PubMed]
- Dehhaghi, M.; Panahi, H.K.S.; Kavyani, B.; Heng, B.; Tan, V.; Braidy, N.; Guillemin, G.J. The Role of Kynurenine Pathway and NAD + Metabolism in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Aging Dis. 2022, 13, 698. [Google Scholar] [CrossRef] [PubMed]
- Dehhaghi, M.; Tan, V.; Heng, B.; Braidy, N.; Mohammadipanah, F.; Guillemin, G.J. Neuroprotective Effect of Myxobacterial Extracts on Quinolinic Acid-Induced Toxicity in Primary Human Neurons. Neurotox. Res. 2019, 35, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Braidy, N.; Guillemin, G.J. Herpetosiphon Secondary Metabolites Inhibit Amyloid-β Toxicity in Human Primary Astrocytes. J. Alzheimers Dis. 2020, 76, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Pérez, R.; Wanders, R.J.A.; Van Karnebeek, C.D.M.; Houtkooper, R.H. NAD + Homeostasis in Human Health and Disease. EMBO Mol. Med. 2021, 13, e13943. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef]
- Israelyan, N.; Margolis, K.G. Serotonin as a Link between the Gut-Brain-Microbiome Axis in Autism Spectrum Disorders. Pharmacol. Res. 2018, 132, 1–6. [Google Scholar] [CrossRef]
- Margolis, K.G.; Stevanovic, K.; Li, Z.; Yang, Q.M.; Oravecz, T.; Zambrowicz, B.; Jhaver, K.G.; Diacou, A.; Gershon, M.D. Pharmacological Reduction of Mucosal but Not Neuronal Serotonin Opposes Inflammation in Mouse Intestine. Gut 2014, 63, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.R.; Gershon, M.D.; Margolis, K.G.; Gertsberg, Z.V.; Cowles, R.A. Neuronal Serotonin Regulates Growth of the Intestinal Mucosa in Mice. Gastroenterology 2012, 143, 408–417.e2. [Google Scholar] [CrossRef]
- Li, Z.; Chalazonitis, A.; Huang, Y.-Y.; Mann, J.J.; Margolis, K.G.; Yang, Q.M.; Kim, D.O.; Cote, F.; Mallet, J.; Gershon, M.D. Essential Roles of Enteric Neuronal Serotonin in Gastrointestinal Motility and the Development/Survival of Enteric Dopaminergic Neurons. J. Neurosci. 2011, 31, 8998–9009. [Google Scholar] [CrossRef]
- Wolf, D.; Klasen, M.; Eisner, P.; Zepf, F.D.; Zvyagintsev, M.; Palomero-Gallagher, N.; Weber, R.; Eisert, A.; Mathiak, K. Central Serotonin Modulates Neural Responses to Virtual Violent Actions in Emotion Regulation Networks. Brain Struct. Funct. 2018, 223, 3327–3345. [Google Scholar] [CrossRef] [PubMed]
- Ursin, R. Serotonin and Sleep. Sleep Med. Rev. 2002, 6, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Migliarini, S.; Pacini, G.; Pelosi, B.; Lunardi, G.; Pasqualetti, M. Lack of Brain Serotonin Affects Postnatal Development and Serotonergic Neuronal Circuitry Formation. Mol. Psychiatry 2013, 18, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Van Kleef, E.S.B.; Gaspar, P.; Bonnin, A. Insights into the Complex Influence of 5-HT Signaling on Thalamocortical Axonal System Development. Eur. J. Neurosci. 2012, 35, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Filip, M.; Bader, M. Overview on 5-HT Receptors and Their Role in Physiology and Pathology of the Central Nervous System. Pharmacol. Rep. 2009, 61, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Ciranna, L. Serotonin as a Modulator of Glutamate- and GABA-Mediated Neurotransmission: Implications in Physiological Functions and in Pathology. Curr. Neuropharmacol. 2006, 4, 101–114. [Google Scholar] [CrossRef]
- Browning, K.N. Role of Central Vagal 5-HT3 Receptors in Gastrointestinal Physiology and Pathophysiology. Front. Neurosci. 2015, 9, 413. [Google Scholar] [CrossRef] [PubMed]
- Mawe, G.M.; Hoffman, J.M. Serotonin Signalling in the Gut—Functions, Dysfunctions and Therapeutic Targets. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 473–486. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Grasset, E.; Mannerås Holm, L.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E.; Bäckhed, F. Gut Microbiota Regulates Maturation of the Adult Enteric Nervous System via Enteric Serotonin Networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.M.; Tyler, K.; MacEachern, S.J.; Balemba, O.B.; Johnson, A.C.; Brooks, E.M.; Zhao, H.; Swain, G.M.; Moses, P.L.; Galligan, J.J.; et al. Activation of Colonic Mucosal 5-HT4 Receptors Accelerates Propulsive Motility and Inhibits Visceral Hypersensitivity. Gastroenterology 2012, 142, 844–854.e4. [Google Scholar] [CrossRef]
- Walldorf, J.; Porzner, M.; Neumann, M.; Joodi, G.; Niess, J.H.; Von Boyen, G.; Mäder, K.; Weissbach, J.; Kleger, A.; Seufferlein, T. The Selective 5-HT1A Agonist SR57746A Protects Intestinal Epithelial Cells and Enteric Glia Cells and Promotes Mucosal Recovery in Experimental Colitis. Inflamm. Bowel Dis. 2022, 28, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Mawe, G.M.; Branchek, T.A.; Gershon, M.D. Peripheral Neural Serotonin Receptors: Identification and Characterization with Specific Antagonists and Agonists. Proc. Natl. Acad. Sci. USA 1986, 83, 9799–9803. [Google Scholar] [CrossRef] [PubMed]
- Brzęczek, M.; Słonka, K.; Hyla-Klekot, L. Melatonin—A Pleiotropic Hormone. Pediatr. Med. Rodz. 2016, 12, 127–133. [Google Scholar] [CrossRef]
- Navabi, S.; Gorrepati, V.S.; Yadav, S.; Chintanaboina, J.; Maher, S.; Demuth, P.; Stern, B.; Stuart, A.; Tinsley, A.; Clarke, K.; et al. Influences and Impact of Anxiety and Depression in the Setting of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 2303–2308. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Licht, T.R. Microbial Tryptophan Catabolites in Health and Disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Bose, C.; Mande, S.S. Tryptophan Metabolism by Gut Microbiome and Gut-Brain-Axis: An in Silico Analysis. Front. Neurosci. 2019, 13, 1365. [Google Scholar] [CrossRef] [PubMed]
- Devlin, A.S.; Marcobal, A.; Dodd, D.; Nayfach, S.; Plummer, N.; Meyer, T.; Pollard, K.S.; Sonnenburg, J.L.; Fischbach, M.A. Modulation of a Circulating Uremic Solute via Rational Genetic Manipulation of the Gut Microbiota. Cell Host Microbe 2016, 20, 709–715. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, J. Indole as an Intercellular Signal in Microbial Communities. FEMS Microbiol. Rev. 2010, 34, 426–444. [Google Scholar] [CrossRef]
- Whitfield-Cargile, C.M.; Cohen, N.D.; Chapkin, R.S.; Weeks, B.R.; Davidson, L.A.; Goldsby, J.S.; Hunt, C.L.; Steinmeyer, S.H.; Menon, R.; Suchodolski, J.S.; et al. The Microbiota-Derived Metabolite Indole Decreases Mucosal Inflammation and Injury in a Murine Model of NSAID Enteropathy. Gut Microbes 2016, 7, 246–261. [Google Scholar] [CrossRef]
- Chimerel, C.; Emery, E.; Summers, D.K.; Keyser, U.; Gribble, F.M.; Reimann, F. Bacterial Metabolite Indole Modulates Incretin Secretion from Intestinal Enteroendocrine L Cells. Cell Rep. 2014, 9, 1202–1208. [Google Scholar] [CrossRef]
- Tennoune, N.; Andriamihaja, M.; Blachier, F. Production of Indole and Indole-Related Compounds by the Intestinal Microbiota and Consequences for the Host: The Good, the Bad, and the Ugly. Microorganisms 2022, 10, 930. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Xiao, H.; Lin, C.; Wong, H.L.X.; Lam, Y.Y.; Gong, M.; Wu, G.; Ning, Z.; Huang, C.; Zhang, Y.; et al. Gut Microbiota-Derived Tryptamine and Phenethylamine Impair Insulin Sensitivity in Metabolic Syndrome and Irritable Bowel Syndrome. Nat. Commun. 2023, 14, 4986. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.B.; Van Benschoten, A.H.; Cimermancic, P.; Donia, M.S.; Zimmermann, M.; Taketani, M.; Ishihara, A.; Kashyap, P.C.; Fraser, J.S.; Fischbach, M.A. Discovery and Characterization of Gut Microbiota Decarboxylases That Can Produce the Neurotransmitter Tryptamine. Cell Host Microbe 2014, 16, 495–503. [Google Scholar] [CrossRef]
- Hubbard, T.D.; Murray, I.A.; Perdew, G.H. Indole and Tryptophan Metabolism: Endogenous and Dietary Routes to Ah Receptor Activation. Drug Metab. Dispos. 2015, 43, 1522–1535. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, A.M.; Pacheco, A.R.; Henrick, B.M.; Taft, D.; Xu, G.; Huda, M.N.; Mishchuk, D.; Goodson, M.L.; Slupsky, C.; Barile, D.; et al. Indole-3-Lactic Acid Associated with Bifidobacterium-Dominated Microbiota Significantly Decreases Inflammation in Intestinal Epithelial Cells. BMC Microbiol. 2020, 20, 357. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.; Kolde, R.; d’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37.e6. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Wu, T.; Li, Y.; Zhou, X.; Ruan, Z. Indole-3-Propionic Acid Improved the Intestinal Barrier by Enhancing Epithelial Barrier and Mucus Barrier. J. Agric. Food Chem. 2021, 69, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Connell, E.; Le Gall, G.; Pontifex, M.G.; Sami, S.; Cryan, J.F.; Clarke, G.; Müller, M.; Vauzour, D. Microbial-Derived Metabolites as a Risk Factor of Age-Related Cognitive Decline and Dementia. Mol. Neurodegener. 2022, 17, 43. [Google Scholar] [CrossRef]
- Rothhammer, V.; Mascanfroni, I.D.; Bunse, L.; Takenaka, M.C.; Kenison, J.E.; Mayo, L.; Chao, C.-C.; Patel, B.; Yan, R.; Blain, M.; et al. Type I Interferons and Microbial Metabolites of Tryptophan Modulate Astrocyte Activity and Central Nervous System Inflammation via the Aryl Hydrocarbon Receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef]
- Chyan, Y.-J.; Poeggeler, B.; Omar, R.A.; Chain, D.G.; Frangione, B.; Ghiso, J.; Pappolla, M.A. Potent Neuroprotective Properties against the Alzheimer B-Amyloid by an Endogenous Melatonin-Related Indole Structure, Indole-3-Propionic Acid. J. Biol. Chem. 1999, 274, 21937–21942. [Google Scholar] [CrossRef]
- Hwang, I.K.; Yoo, K.; Li, H.; Park, O.K.; Lee, C.H.; Choi, J.H.; Jeong, Y.; Lee, Y.L.; Kim, Y.; Kwon, Y.; et al. Indole-3-propionic Acid Attenuates Neuronal Damage and Oxidative Stress in the Ischemic Hippocampus. J. Neurosci. Res. 2009, 87, 2126–2137. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-Y.; Li, J.-R.; Wang, Y.-Y.; Lin, S.-Y.; Ou, Y.-C.; Lin, C.-J.; Wang, J.-D.; Liao, S.-L.; Chen, C.-J. Indoxyl Sulfate Caused Behavioral Abnormality and Neurodegeneration in Mice with Unilateral Nephrectomy. Aging 2021, 13, 6681–6701. [Google Scholar] [CrossRef]
- Moloney, G.M.; O’Leary, O.F.; Salvo-Romero, E.; Desbonnet, L.; Shanahan, F.; Dinan, T.G.; Clarke, G.; Cryan, J.F. Microbial Regulation of Hippocampal miRNA Expression: Implications for Transcription of Kynurenine Pathway Enzymes. Behav. Brain Res. 2017, 334, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Yu, D.; Sun, J.; Wu, X.; Xin, Z.; Deng, B.; Fan, L.; Fu, J.; Ge, L.; Ren, W. Characterizing the Influence of Gut Microbiota on Host Tryptophan Metabolism with Germ-Free Pigs. Anim. Nutr. 2022, 11, 190–200. [Google Scholar] [CrossRef]
- Schwarcz, R.; Foo, A.; Sathyasaikumar, K.V.; Notarangelo, F.M. The Probiotic Lactobacillus Reuteri Preferentially Synthesizes Kynurenic Acid from Kynurenine. Int. J. Mol. Sci. 2024, 25, 3679. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, M.; Liu, Y.-X.; Li, L.; Chai, X.; Zheng, W.; Chen, S.; Zhu, X.; Zhao, S. High-Fat Diet-Disturbed Gut Microbiota-Colonocyte Interactions Contribute to Dysregulating Peripheral Tryptophan-Kynurenine Metabolism. Microbiome 2023, 11, 154. [Google Scholar] [CrossRef]
- Mu, C.; Choudhary, A.; Mayengbam, S.; Barrett, K.T.; Rho, J.M.; Shearer, J.; Scantlebury, M.H. Seizure Modulation by the Gut Microbiota and Tryptophan-Kynurenine Metabolism in an Animal Model of Infantile Spasms. eBioMedicine 2022, 76, 103833. [Google Scholar] [CrossRef]
- Zhao, L.-P.; Wu, J.; Quan, W.; Zhou, Y.; Hong, H.; Niu, G.-Y.; Li, T.; Huang, S.-B.; Qiao, C.-M.; Zhao, W.-J.; et al. DSS-Induced Colitis Activates the Kynurenine Pathway in Serum and Brain by Affecting IDO-1 and Gut Microbiota. Front. Immunol. 2023, 13, 1089200. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Li, J.; Gao, X.; Yang, H.; Su, J.; Weng, R.; Gao, Y.; Ni, W.; Gu, Y. Involvement of the Gut-Brain Axis in Vascular Depression via Tryptophan Metabolism: A Benefit of Short Chain Fatty Acids. Exp. Neurol. 2022, 358, 114225. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Fukui, H.; Eda, H.; Kitayama, Y.; Hara, K.; Kodani, M.; Tomita, T.; Oshima, T.; Watari, J.; Miwa, H. Involvement of Gut Microbiota in the Association between Gastrointestinal Motility and 5-HT Expression/M2 Macrophage Abundance in the Gastrointestinal Tract. Mol. Med. Rep. 2017, 16, 3482–3488. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Wang, J.; Jiang, Y.; Zhu, X.; Zhang, Z.; Wang, Y.; Shu, X.; Deng, Y.; Zhang, F. Bifidobacterium Lactis TY-S01 Prevents Loperamide-Induced Constipation by Modulating Gut Microbiota and Its Metabolites in Mice. Front. Nutr. 2022, 9, 890314. [Google Scholar] [CrossRef] [PubMed]
- Xue, K.; Tao, R.; Wu, Q.; Zhang, L.; Sun, Z.; Yu, X.; Meng, J.; Bao, N.; Zhao, J. Enteropathogenic Escherichia Coli Mediates CoCrMo Particle-Induced Peri-Implant Osteolysis by Increasing Peripheral 5-HT. Front. Cell. Infect. Microbiol. 2022, 11, 796679. [Google Scholar] [CrossRef]
- Li, H.; Wang, P.; Huang, L.; Li, P.; Zhang, D. Effects of Regulating Gut Microbiota on the Serotonin Metabolism in the Chronic Unpredictable Mild Stress Rat Model. Neurogastroenterol. Motil. 2019, 31, e13677. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, X.; Wang, K.; Li, Y.; Nagaoka, K.; Li, C. Gut Microbiota Intervention Attenuates Thermogenesis in Broilers Exposed to High Temperature through Modulation of the Hypothalamic 5-HT Pathway. J. Anim. Sci. Biotechnol. 2023, 14, 159. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liang, W.; Zhang, W.; Huang, Z.; Liang, H.; Liu, Q. Fecal Microbiota Transplantation Repairs Intestinal Permeability and Regulates the Expression of 5-HT to Influence Alcohol-Induced Depression-like Behaviors in C57BL/6J Mice. Front. Microbiol. 2024, 14, 1241309. [Google Scholar] [CrossRef]
- Bunnett, N.W. Neuro-humoral Signalling by Bile Acids and the TGR5 Receptor in the Gastrointestinal Tract. J. Physiol. 2014, 592, 2943–2950. [Google Scholar] [CrossRef]
- Chen, Z.; Feng, J.; Hu, S.; Hua, Y.; Ma, S.; Fu, W.; Yang, Q.; Zhang, X. Bacillus Subtilis Promotes the Release of 5-HT to Regulate Intestinal Peristalsis in STC Mice via Bile Acid and Its Receptor TGR5 Pathway. Dig. Dis. Sci. 2022, 67, 4410–4421. [Google Scholar] [CrossRef]
- Cai, J.; Cheung, J.; Cheung, S.W.M.; Chin, K.T.C.; Leung, R.W.K.; Lam, R.S.T.; Sharma, R.; Yiu, J.H.C.; Woo, C.W. Butyrate Acts as a Positive Allosteric Modulator of the 5-HT Transporter to Decrease Availability of 5-HT in the Ileum. Br. J. Pharmacol. 2024, 181, 1654–1670. [Google Scholar] [CrossRef]
- Lin, F.; Wang, X.; Luo, R.; Yuan, B.; Ye, S.; Yang, T.; Xiao, L.; Chen, J. Maternal LPS Exposure Enhances the 5-HT Level in the Prefrontal Cortex of Autism-like Young Offspring. Brain Sci. 2023, 13, 958. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yang, Y.; Song, W.; Jiang, C.; Zhang, Y.; Zhang, A.; Lin, Z.; Ke, X. Untargeted Metabonomic Analysis of a Cerebral Stroke Model in Rats: A Study Based on UPLC–MS/MS. Front. Neurosci. 2023, 17, 1084813. [Google Scholar] [CrossRef] [PubMed]
- Saccaro, L.F.; Pico, F.; Chadenat, M.-L.; Richard, O.; Launay, J.-M.; Bastenaire, B.; Jullien, P.; Lambert, J.; Feuga, V.; Macquet, M.; et al. Platelet, Plasma, Urinary Tryptophan-Serotonin-Kynurenine Axis Markers in Hyperacute Brain Ischemia Patients: A Prospective Study. Front. Neurol. 2022, 12, 782317. [Google Scholar] [CrossRef]
- Darlington, L.G.; Mackay, G.M.; Forrest, C.M.; Stoy, N.; George, C.; Stone, T.W. Altered Kynurenine Metabolism Correlates with Infarct Volume in Stroke. Eur. J. Neurosci. 2007, 26, 2211–2221. [Google Scholar] [CrossRef] [PubMed]
- Cuartero, M.I.; Ballesteros, I.; De La Parra, J.; Harkin, A.L.; Abautret-Daly, A.; Sherwin, E.; Fernández-Salguero, P.; Corbí, Á.L.; Lizasoain, I.; Moro, M.A. L-Kynurenine/Aryl Hydrocarbon Receptor Pathway Mediates Brain Damage After Experimental Stroke. Circulation 2014, 130, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Mangas, A.; Yajeya, J.; González, N.; Ruiz, I.; Duleu, S.; Geffard, M.; Coveñas, R. Overexpression of Kynurenic Acid in Stroke: An Endogenous Neuroprotector? Ann. Anat.-Anat. Anz. 2017, 211, 33–38. [Google Scholar] [CrossRef]
- Alme, K.N.; Ulvik, A.; Askim, T.; Assmus, J.; Mollnes, T.E.; Naik, M.; Næss, H.; Saltvedt, I.; Ueland, P.M.; Knapskog, A.B. Neopterin and Kynurenic Acid as Predictors of Stroke Recurrence and Mortality: A Multicentre Prospective Cohort Study on Biomarkers of Inflammation Measured Three Months after Ischemic Stroke. BMC Neurol. 2021, 21, 476. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Fang, Y.; Sherchan, P.; Lu, Q.; Lenahan, C.; Zhang, J.H.; Zhang, J.; Tang, J. Kynurenine/Aryl Hydrocarbon Receptor Modulates Mitochondria-Mediated Oxidative Stress and Neuronal Apoptosis in Experimental Intracerebral Hemorrhage. Antioxid. Redox Signal. 2022, 37, 1111–1129. [Google Scholar] [CrossRef]
- Ganesh, B.P.; Peesh, P.; Blasco, M.P.; Hamamy, A.E.; Khan, R.; Guzman, G.; Honarpisheh, P.; Mohan, E.C.; Goodman, G.W.; Nguyen, J.N.; et al. Restoring a Balanced Pool of Host-Derived and Microbiota-Derived Ligands of the Aryl Hydrocarbon Receptor Is Beneficial after Stroke. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Rothhammer, V.; Borucki, D.M.; Tjon, E.C.; Takenaka, M.C.; Chao, C.-C.; Ardura-Fabregat, A.; De Lima, K.A.; Gutiérrez-Vázquez, C.; Hewson, P.; Staszewski, O.; et al. Microglial Control of Astrocytes in Response to Microbial Metabolites. Nature 2018, 557, 724–728. [Google Scholar] [CrossRef]
- Xie, Y.; Zou, X.; Han, J.; Zhang, Z.; Feng, Z.; Ouyang, Q.; Hua, S.; Liu, Z.; Li, C.; Cai, Y.; et al. Indole-3-Propionic Acid Alleviates Ischemic Brain Injury in a Mouse Middle Cerebral Artery Occlusion Model. Exp. Neurol. 2022, 353, 114081. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Chen, X.; Yu, C.; Deng, Y.; Zhang, Y.; Chen, S.; Chen, X.; Chen, K.; Yang, Y.; Ling, W. Gut Microbially Produced Indole-3-Propionic Acid Inhibits Atherosclerosis by Promoting Reverse Cholesterol Transport and Its Deficiency Is Causally Related to Atherosclerotic Cardiovascular Disease. Circ. Res. 2022, 131, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Konopelski, P.; Chabowski, D.; Aleksandrowicz, M.; Kozniewska, E.; Podsadni, P.; Szczepanska, A.; Ufnal, M. Indole-3-Propionic Acid, a Tryptophan-Derived Bacterial Metabolite, Increases Blood Pressure via Cardiac and Vascular Mechanisms in Rats. Am. J. Physiol. Regul. Intgr. Comp. Physiol. 2021, 321, R969–R981. [Google Scholar] [CrossRef]
- Yoon, N.; Kim, Y.; Kim, S.-D.; Kim, M.; Jung, B.H.; Song, Y.S. Investigation of Long-Term Metabolic Alteration after Stroke in tMCAO (Transient Middle Cerebral Artery Occlusion) Mouse Model Using Metabolomics Approach. Neurosci. Lett. 2022, 774, 136492. [Google Scholar] [CrossRef] [PubMed]
- Marsiglia, R.; Marangelo, C.; Vernocchi, P.; Scanu, M.; Pane, S.; Russo, A.; Guanziroli, E.; Del Chierico, F.; Valeriani, M.; Molteni, F.; et al. Gut Microbiota Ecological and Functional Modulation in Post-Stroke Recovery Patients: An Italian Study. Microorganisms 2023, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Teunis, C.J.; Stroes, E.S.G.; Boekholdt, S.M.; Wareham, N.J.; Murphy, A.J.; Nieuwdorp, M.; Hazen, S.L.; Hanssen, N.M.J. Tryptophan Metabolites and Incident Cardiovascular Disease: The EPIC-Norfolk Prospective Population Study. Atherosclerosis 2023, 387, 117344. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Gan, J.; Xu, F.; Li, L.; Han, L.; Peng, C.; Bao, Q.; Xiao, L.; Peng, D. RNA Sequencing for Gene Expression Profiles in a Rat Model of Middle Cerebral Artery Occlusion. Biomed. Res. Int. 2018, 2018, 2465481. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Sainz, R.M.; Mayo, J.C.; León, J.; Reiter, R.J. Physiological Ischemia/Reperfusion Phenomena and Their Relation to Endogenous Melatonin Production: An Hypothesis. Endocrine 2005, 27, 149–158. [Google Scholar] [CrossRef]
- Hajsl, M.; Hlavackova, A.; Broulikova, K.; Sramek, M.; Maly, M.; Dyr, J.E.; Suttnar, J. Tryptophan Metabolism, Inflammation, and Oxidative Stress in Patients with Neurovascular Disease. Metabolites 2020, 10, 208. [Google Scholar] [CrossRef]
- Mortensen, J.K.; Kraglund, K.L.; Johnsen, S.P.; Mors, O.; Andersen, G.; Buttenschøn, H.N. The Serotonin Transporter Gene Polymorphisms and Risk of Ischemic Stroke. Cerebrovasc. Dis. 2018, 45, 187–192. [Google Scholar] [CrossRef]
- Aguiar, R.P.D.; Newman-Tancredi, A.; Prickaerts, J.; Oliveira, R.M.W.D. The 5-HT1A Receptor as a Serotonergic Target for Neuroprotection in Cerebral Ischemia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 109, 110210. [Google Scholar] [CrossRef]
- Liu, D.; Tan, S.; Zhou, Z.; Gu, S.; Zuo, H. Trimethylamine N-Oxide, β-Alanine, Tryptophan Index, and Vitamin B6-Related Dietary Patterns in Association with Stroke Risk. Nutr. Metab. Cardiovasc. Dis. 2024, 34, 1179–1188. [Google Scholar] [CrossRef]
- Bhave, V.M.; Ament, Z.; Patki, A.; Gao, Y.; Kijpaisalratana, N.; Guo, B.; Chaudhary, N.S.; Guarniz, A.G.; Gerszten, R.; Correa, A.; et al. Plasma Metabolites Link Dietary Patterns to Stroke Risk. Ann. Neurol. 2023, 93, 500–510. [Google Scholar] [CrossRef]
- Joshi, B.; Panda, S.K.; Jouneghani, R.S.; Liu, M.; Parajuli, N.; Leyssen, P.; Neyts, J.; Luyten, W. Antibacterial, Antifungal, Antiviral, and Anthelmintic Activities of Medicinal Plants of Nepal Selected Based on Ethnobotanical Evidence. Evid.-Based. Compl. Alt. 2020, 2020, 1043471. [Google Scholar] [CrossRef]
- Li, J.; Zhao, T.; Qiao, H.; Li, Y.; Xia, M.; Wang, X.; Liu, C.; Zheng, T.; Chen, R.; Xie, Y.; et al. Research Progress of Natural Products for the Treatment of Ischemic Stroke. J. Integr. Neurosci. 2022, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wang, T.; Wu, Q.; Dong, X.; Shen, F.; Liu, D.; Qin, X.; Yan, L.; Wan, Q. Analysis of Microbiota in Elderly Patients with Acute Cerebral Infarction. PeerJ 2019, 7, e6928. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, X.; Pan, D.; Liu, Y.; Yan, X.; Tang, Y.; Tao, M.; Gong, L.; Zhang, T.; Woods, C.R.; et al. Dysbiosis Characteristics of Gut Microbiota in Cerebral Infarction Patients. Transl. Neurosci. 2020, 11, 124–133. [Google Scholar] [CrossRef]
- Haak, B.W.; Westendorp, W.F.; van Engelen, T.S.; Brands, X.; Brouwer, M.C.; Vermeij, J.D.; Hugenholtz, F.; Verhoeven, A.; Derks, R.J.; Giera, M.; et al. Disruptions of Anaerobic Gut Bacteria Are Associated with Stroke and Post-Stroke Infection: A Prospective Case–Control Study. Transl. Stroke Res. 2020, 12, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Lou, Y.; Liu, L.; Liu, Y.; Zhang, W.; Deng, J.; Guan, Y.; She, M.; You, X.; Liu, M.; et al. Gut Microbiotic Features Aiding the Diagnosis of Acute Ischemic Stroke. Front. Cell. Infect. Microbiol. 2020, 10, 587284. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wu, Q.; Wang, H.; Gao, X.; Xu, R.; Cui, Z.; Zhu, J.; Zeng, X.; Zhou, H.; He, Y.; et al. Dysbiosis of Gut Microbiota and Short-chain Fatty Acids in Acute Ischemic Stroke and the Subsequent Risk for Poor Functional Outcomes. J. Parenter. Enter. Nutr. 2021, 45, 518–529. [Google Scholar] [CrossRef]
- Chen, X.; Wu, Q.; Gao, X.; Wang, H.; Zhu, J.; Xia, G.; He, Y.; Song, W.; Xu, K. Gut Microbial Dysbiosis Associated with Type 2 Diabetes Aggravates Acute Ischemic Stroke. mSystems 2021, 6, e01304-21. [Google Scholar] [CrossRef]
- Chen, L.; Wang, S.; Zhang, Y.; Li, Y.; Zhang, X.; Ma, J.; Zou, X.; Yao, T.; Li, S.; Chen, J.; et al. Multi-Omics Reveals Specific Host Metabolism-Microbiome Associations in Intracerebral Hemorrhage. Front. Cell. Infect. Microbiol. 2022, 12, 999627. [Google Scholar] [CrossRef]
- Li, T.; Sun, Q.; Feng, L.; Yan, D.; Wang, B.; Li, M.; Xiong, X.; Ma, D.; Gao, Y. Uncovering the Characteristics of the Gut Microbiota in Patients with Acute Ischemic Stroke and Phlegm-Heat Syndrome. PLoS ONE 2022, 17, e0276598. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Zhao, H.; Cao, R.; Dang, Y.; Yu, B. Imbalance of Microbacterial Diversity Is Associated with Functional Prognosis of Stroke. Neural Plast. 2023, 2023, 6297653. [Google Scholar] [CrossRef]
- Yu, S.; Chen, J.; Zhao, Y.; Liao, X.; Chen, Q.; Xie, H.; Liu, J.; Sun, J.; Zhi, S. Association Analysis of the Gut Microbiota in Predicting Outcomes for Patients with Acute Ischemic Stroke and H-Type Hypertension. Front. Neurol. 2023, 14, 1275460. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, X.; Mu, X. Systematic Insights into the Relationship between the Microbiota–Gut–Brain Axis and Stroke with the Focus on Tryptophan Metabolism. Metabolites 2024, 14, 399. https://doi.org/10.3390/metabo14080399
Shen X, Mu X. Systematic Insights into the Relationship between the Microbiota–Gut–Brain Axis and Stroke with the Focus on Tryptophan Metabolism. Metabolites. 2024; 14(8):399. https://doi.org/10.3390/metabo14080399
Chicago/Turabian StyleShen, Xinyu, and Xiaoqin Mu. 2024. "Systematic Insights into the Relationship between the Microbiota–Gut–Brain Axis and Stroke with the Focus on Tryptophan Metabolism" Metabolites 14, no. 8: 399. https://doi.org/10.3390/metabo14080399