
Metabolomic Prediction of Cadmium Nephrotoxicity in the Snail Helix aspersa maxima



Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Housing
2.2. Chemicals
2.3. Cd Exposure
2.4. Hemolymph Sampling
2.5. 1H-NMR Spectroscopy and Spectral Data Analysis
2.5.1. Sample Preparation
2.5.2. Acquisition of Metabolic Profiles
2.5.3. Statistical Analysis
3. Results
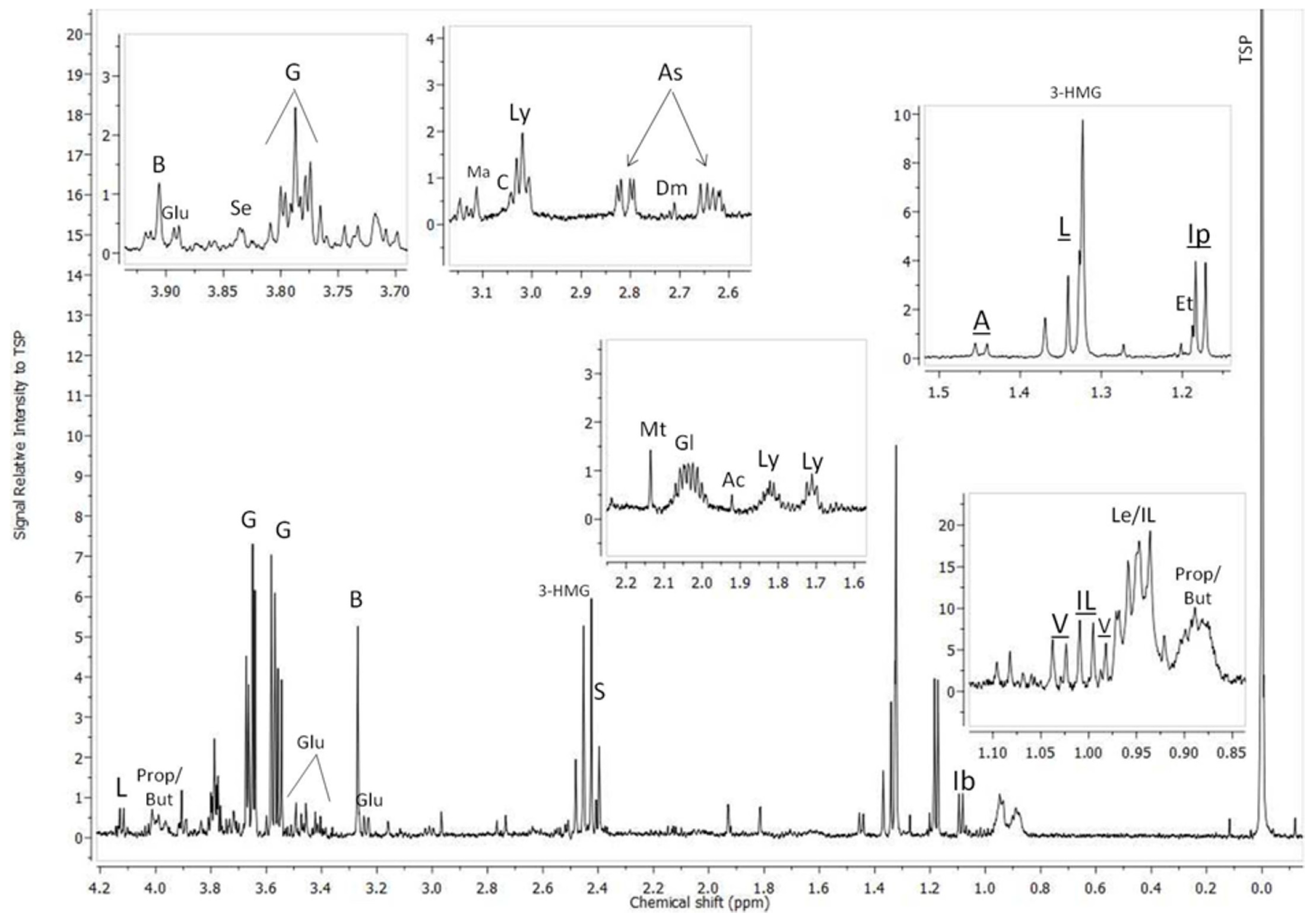
3.1. 1H-NMR Profiles of Kidney Extracts Collected from Cd-Exposed Snails
3.2. 1H-NMR Profiles of Hemolymph Samples Collected from Cd-Exposed Snails
4. Discussion
Metabolic Impact of Cd Exposure in Snails
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ribera, D.; Taberly, J. Mélanges de Polluants, Toxicité, Écotoxicité et Évaluation des Risques. Etude RECORD n°08-0668/1A. 2011. 295p. Available online: https://record-net.org/media/etudes/38/public/synthese/synth-record08-0668-1a.pdf (accessed on 3 July 2024).
- Lavelle, P.; Spain, A.V. Soil Ecology; Kluwer Scientific: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barrot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.-P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Andral, B.; Galgani, F.; Tomasino, C.; Bouchoucha, M.; Blottiere, C.; Scarpato, A.; Benedicto, J.; Deudero, S.; Calvo, M.; Alexandro, C.; et al. Chemical contamination baseline in the Western basin of the Mediterranean Sea based on transplanted mussels. Arch. Environ. Contam. Toxicol. 2011, 61, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Prudente, M.S.; Kanatireklap, S.; Subramanian, A. Mussel watch: Marine pollution monitoring of butyltins and organochlorines in coastal waters of Thailand, Philippines and India. Ocean. Coast. Manag. 2000, 43, 819–839. [Google Scholar] [CrossRef]
- Regoli, F.; Orlando, E. Mytilus galloprovincialis as a bioindicator of lead pollution: Biological variables and cellular responses. Sci. Total Environ. 1993, 134, 1283–1292. [Google Scholar] [CrossRef]
- Conti, M.E.; Botré, F. Honeybees and Their Products as Potential Bio-Indicators of Heavy Metal Contamination. Environ. Monit. Assess. 2001, 69, 267–282. [Google Scholar] [CrossRef]
- Pérès, G.; Vandenbulcke, F.; Guernion, M.; Hedde, M.; Beguiristain, T.; Douay, F.; Houot, S.; Piron, D.; Richard, A.; Bispo, A.; et al. Earthworm indicators as tools for soil monitoring, characterization and risk assessment: An example from the national Bioindicator programme. Pedobiol. Int. J. Soil Biol. 2011, 54, S77–S87. [Google Scholar] [CrossRef]
- Taylor, J.W. Life history of British Helices: Helix (Pomatia) aspersa Müll. J. Conchol. 1883, 4, 94. [Google Scholar]
- Grara, N.; Boucenna, M.; Atailia, A.; Berrebbah, H.; Djebar, M.R. Stress oxydatif des poussières métalliques du complexe sidérurgique d’Annaba (Nord-Est algérien) chez l’escargot Helix aspersa. Environ. Risques Santé 2012, 11, 221–229. [Google Scholar]
- Chabicovsky, M.; Niederstätter, H.; Thaler, R.; Hödl, E.; Parson, W.; Ross Manith, V.; Dallinger, R. Localization and quantification of Cd- and Cu-specific metallothionein isoform mRNA in cells and organs of the terrestrial gastropod Helix pomatia. Toxicol. Appl. Pharm. 2003, 190, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Coeurdassier, M. Utilisation de Mollusques Gastéropodes Pulmonés Terrestres (Helix aspersa) et Aquatiques (Lymnia stagnalis et Lymnia palustris) Comme Indicateurs de Pollution par les Éléments Métalliques et les Xénobiotiques. Ph.D. Thesis, Université de Franche-Comté, Besançon, France, 2001; 281p. [Google Scholar]
- Lagadic, L.; Caquet, T.; Amiard, J.-C. Biomarqueurs en écotoxicologie: Principes et définitions. In Biomarqueurs en Ecotoxicologie: Aspects Fondamentaux; Lagadic, L., Caquet, T., Amiard, J.-C., Ramade, F., Eds.; Collection d’Ecologie; Masson Editeur: Paris, France, 1997; pp. 1–9. [Google Scholar]
- Bocquené, G.; Galgani, F.; Truquet, P. Characterization and assay conditions for use of AChE activity from several marine species in pollution monitoring. Mar. Environ. Res. 1990, 30, 75–89. [Google Scholar] [CrossRef]
- Bucheli, T.D.; Fent, K. Induction of Cytochrome P450 as a Biomarker for Environmental Contamination in Aquatic Ecosystems. Crit. Rev. Environ. Sci. Technol. 1995, 25, 201–268. [Google Scholar] [CrossRef]
- Fasitsas, D.; Theocharis, S.E.; Zoulas, D.; Chrissimou, S.; Deliconstantinos, G. Time-dependent cadmium-neurotoxicity in rat brain synaptosomal plasma membranes. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 100, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.E. Bleeding of pulmonate snails. Lab. Anim. 1994, 28, 277–278. [Google Scholar] [CrossRef]
- Devalckeneer, A.; Marion, B.; Raphaël, C.; Colet, J.-M. Proton Nuclear Magnetic Resonance (1H NMR) profiling of isolated organs in the snail Helix aspersa maxima. Ecol. Indic. 2019, 105, 177–187. [Google Scholar]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W.; Fields, J.H.A.; Mommsen, T.P. Metabolic and enzyme regulation during rest-to-work transition: A mammal versus mollusc comparison. In The Mollusca, Volume 1. Metabolic Biochemistry and Molecular Biomechanics; Hochachka, P.W., Ed.; Elsevier: Amsterdam, The Netherlands, 1983; pp. 55–89. [Google Scholar]
- Stuart, J.A.; Ballantyne, J.S. Subcellular organization of intermediary metabolism in the hepatopancreas of the terrestrial snail, Cepaea nemoralis: A cytosolic β-hydroxybutyrate dehydrogenase. J. Exp. Zool. 1996, 274, 291–299. [Google Scholar] [CrossRef]
- Monteiro, M.S.; Barros, A.S.; Pinto, J.; Carvalho, M.; Pires-Luís, A.S.; Henrique, R.; Jerónimo, C.; Bastos, M.d.L.; Gil, A.M.; de Pinho, P.G. Nuclear Magnetic Resonance metabolomics reveals an excretory metabolic signature of renal cell carcinoma. Sci. Rep. 2016, 6, 37275. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Liu, X.; Zhou, Q.; Hao, X.; Lu, Y.; Guo, S.; Wang, W.; Lin, D.; Chen, N. 1 H NMR spectroscopy analysis of metabolites in the kidneys provides new insight into pathophysiological mechanisms: Applications for treatment with Cordyceps sinensis. Nephrol. Dial. Transplant. 2012, 27, 556–565. [Google Scholar] [CrossRef]
- Qi, S.; Ouyang, X.; Wang, L.; Peng, W.; Wen, J.; Dai, Y. A Pilot Metabolic Profiling Study in Serum of Patients with Chronic Kidney Disease Based on 1H-NMR-Spectroscopy. Clin. Transl. Sci. 2012, 5, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Le Roy, C.I.; Mappley, L.J.; La Ragione, R.M.; Woodward, M.J.; Claus, S.P. NMR-based metabolic characterization of chicken tissues and biofluids: A model for avian research. Metabolomics 2016, 12, 157. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.C.; Hewitt, P. Biomarkers for Drug-Induced Renal Damage and Nephrotoxicity—An Overview for Applied Toxicology. AAPS J. 2011, 13, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Delgado, C.A.; Balbueno Guerreiro, G.B.; Diaz Jacques, C.E.; de Moura Coelho, D.; Sitta, A.; Manfredini, V.; Wajner, M.; Vargas, C.R. Prevention by L-carnitine of DNA damage induced by 3-hydroxy-3-methylglutaric and 3-methylglutaric acids and experimental evidence of lipid and DNA damage in patients with 3-hydroxy-3-methylglutaric aciduria. Arch. Biochem. Biophys. 2019, 668, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Racusen, L.C.; Finn, W.F.; Whelton, A.; Solez, K. Mechanisms of lysine-induced acute renal failure in rats. Kidney Int. 1985, 27, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Grünert, S.C.; Schlatter, S.M.; Schmitt, R.N.; Gemperle-Britschgi, C.; Mrázová, L.; Balcı, M.C.; Bischof, F.; Çoker, M.; Das, A.M.; Demirkol, M.; et al. 3-Hydroxy-3-methylglutaryl-coenzyme A lyase deficiency: Clinical presentation and outcome in a series of 37 patients. Mol. Genet. Metab. 2017, 121, 206–215. [Google Scholar] [CrossRef]
- Da Rosa, M.S.; Seminotti, B.; Amaral, U.; Fernandes, C.G.; Gasparotto, J.; Moreira, C.F.; Gelain, D.P.; Wajner, M.; Leipnitz, G. Redox homeostasis is compromised in vivo by the metabolites accumulating in 3-hydroxy-3-methylglutaryl-CoA lyase deficiency in rat cerebral cortex and liver. Free. Radic. Res. 2013, 47, 1066–1075. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Metabolite | Isoleucine | Valine | Hydroxybut. | Fucose | Lactate | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 140 | 120 | 146 | 140 | 33 | |||||
| 24 h Cd | 73 | 125 | 43 | 45 | 65 | |||||
| Metabolite | Alanine | Putrescine | Acetate | N-acetyl-LY. | Glutamate | |||||
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 90 | 285 | 157 | 114 | 181 | |||||
| 24 h Cd | 208 | 108 | 125 | 112 | 83 | |||||
| Metabolite | Glutamine | Succinate | Riboflavine | Citrate | Aspartate | |||||
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 85 | 80 | 107 | 276 | 95 | |||||
| 24 h Cd | 146 | 162 | 63 | 70 | 178 | |||||
| Metabolite | Choline | Betaïne | Carnitine | β-glc | Glycine | |||||
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 82 | 90 | 146 | 97 | 96 | |||||
| 24 h Cd | 77 | 135 | 45 | 53 | 63 | |||||
| Metabolite | Glycerol | Maltose | β-Galactose | Adenosine | Anserine | |||||
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 120 | 115 | 201 | 147 | 152 | |||||
| 24 h Cd | 73 | 66 | 44 | 214 | 47 | |||||
| Metabolite | 7.52 ppm | Urocanate | Hypoxanthine | |||||||
| Control | 100 | 100 | 100 | |||||||
| 4 h Cd | 71 | 80 | 164 | |||||||
| 24 h Cd | 52 | 225 | 181 | |||||||
| High | 200 | 185 | 150 | 120 | 100 | 80 | 50 | 25 | 0 | Low |
| Metabolite | Isoleucine | Isobutyrate | Isoprop. | Lactate | Alanine | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 60 | 134 | 129 | 96 | 69 | |||||
| 24 h Cd | 149 | 106 | 101 | 42 | 121 | |||||
| Metabolite | Acetate | Glutamate | Succinate | Lysine | Glucose | |||||
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 60 | 81 | 100 | 122 | 79 | |||||
| 24 h Cd | 110 | 76 | 26 | 132 | 73 | |||||
| Metabolite | Glycerol | Serine | Prop/But | Betaïne | 3-HMG | |||||
| Control | 100 | 100 | 100 | 100 | 100 | |||||
| 4 h Cd | 106 | 97 | 73 | 127 | 155 | |||||
| 24 h Cd | 112 | 83 | 142 | 133 | 54 | |||||
| High | 200 | 185 | 150 | 120 | 100 | 80 | 50 | 25 | 0 | Low |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devalckeneer, A.; Bouviez, M.; Gautier, A.; Colet, J.-M. Metabolomic Prediction of Cadmium Nephrotoxicity in the Snail Helix aspersa maxima. Metabolites 2024, 14, 455. https://doi.org/10.3390/metabo14080455
Devalckeneer A, Bouviez M, Gautier A, Colet J-M. Metabolomic Prediction of Cadmium Nephrotoxicity in the Snail Helix aspersa maxima. Metabolites. 2024; 14(8):455. https://doi.org/10.3390/metabo14080455
Chicago/Turabian StyleDevalckeneer, Aude, Marion Bouviez, Amandine Gautier, and Jean-Marie Colet. 2024. "Metabolomic Prediction of Cadmium Nephrotoxicity in the Snail Helix aspersa maxima" Metabolites 14, no. 8: 455. https://doi.org/10.3390/metabo14080455
APA StyleDevalckeneer, A., Bouviez, M., Gautier, A., & Colet, J.-M. (2024). Metabolomic Prediction of Cadmium Nephrotoxicity in the Snail Helix aspersa maxima. Metabolites, 14(8), 455. https://doi.org/10.3390/metabo14080455