
The Impact of Varying Pasture Levels on the Metabolomic Profile of Bovine Ruminal Fluid

 , , , ,
, , , ,  ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Design
2.3. Metabolomic Analysis
2.4. Statistical Analysis
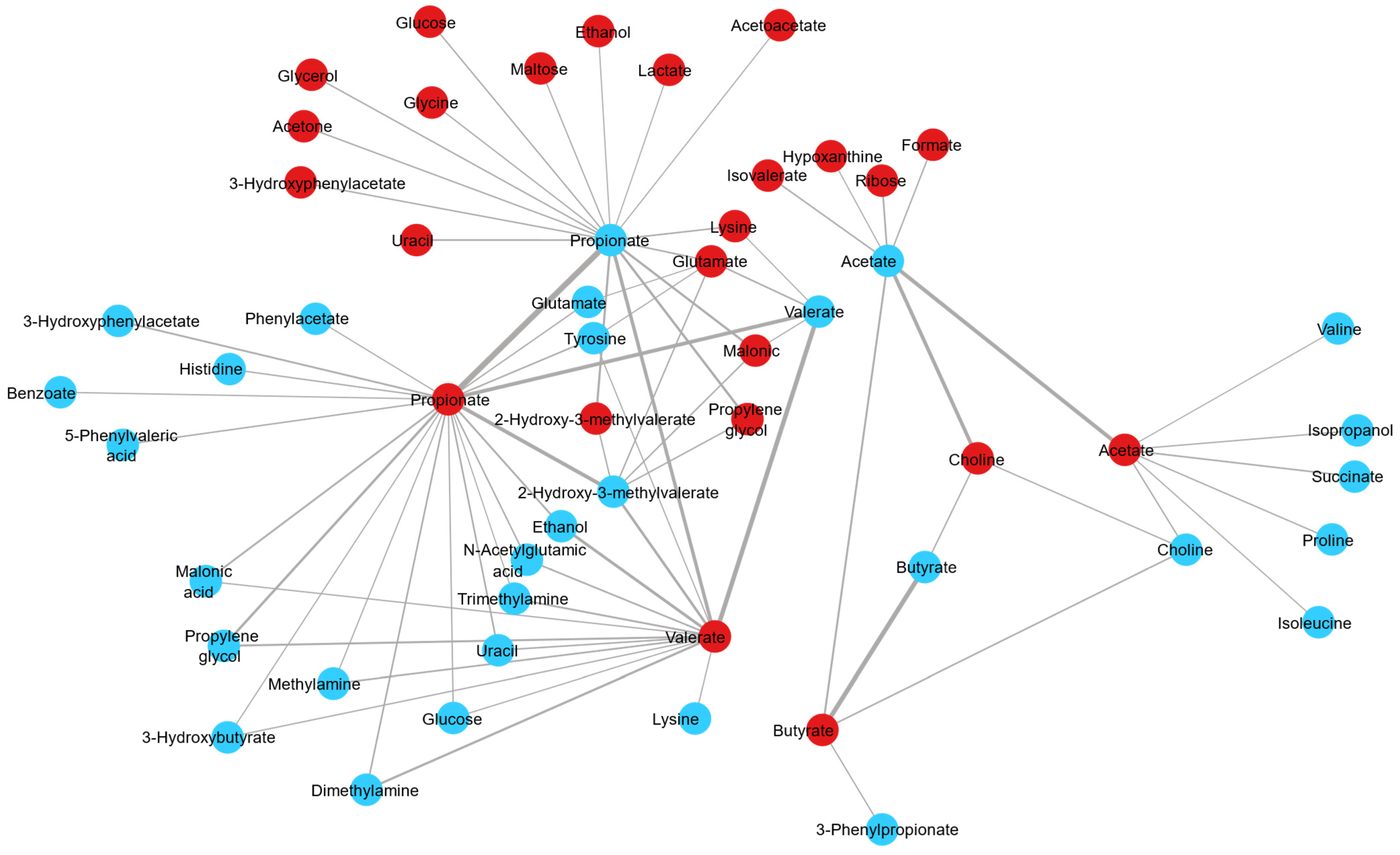
3. Results
3.1. Pasture Feeding Alters the Metabolite Profile of Bovine Ruminal Fluid
3.2. Relationship between Ruminal Fluid Morning and Evening Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lilian, M.; Rawlynce, B.; Charles, G.; Felix, K. Potential role of rumen bacteria in modulating milk production and composition of admixed dairy cows. Lett. Appl. Microbiol. 2023, 76, ovad007. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.-Y.; Sun, H.-Z.; Wu, X.-H.; Liu, J.-X.; Guan, L.L. Multi-omics reveals that the rumen microbiome and its metabolome together with the host metabolome contribute to individualized dairy cow performance. Microbiome 2020, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, S.; Mee, J.F.; Lewis, E.; Galvin, N.; Hennessy, D. Feeding System Effects on Dairy Cow Rumen Function and Milk Production. Animals 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Saleem, F.; Bouatra, S.; Guo, A.C.; Psychogios, N.; Mandal, R.; Dunn, S.M.; Ametaj, B.N.; Wishart, D.S. The Bovine Ruminal Fluid Metabolome. Metabolomics 2013, 9, 360–378. [Google Scholar] [CrossRef]
- Linn, J.; Otterby, D.; Howard, T.W.; Shaver, R. The Ruminant Digestive System. Available online: https://extension.umn.edu/dairy-nutrition/ruminant-digestive-system#stomach-compartments-1000460 (accessed on 14 October 2023).
- Wang, L.; Zhang, G.; Li, Y.; Zhang, Y. Effects of High Forage/Concentrate Diet on Volatile Fatty Acid Production and the Microorganisms Involved in VFA Production in Cow Rumen. Animals 2020, 10, 223. [Google Scholar] [CrossRef]
- McCann, J.C.; Wickersham, T.A.; Loor, J.J. High-throughput Methods Redefine the Rumen Microbiome and Its Relationship with Nutrition and Metabolism. Bioinform. Biol. Insights 2014, 8, BBI-S15389. [Google Scholar] [CrossRef]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. [Google Scholar] [CrossRef]
- de Almeida, R.T.R.; do Prado, R.M.; Porto, C.; dos Santos, G.T.; Huws, S.A.; Pilau, E.J. Exploring the rumen fluid metabolome using liquid chromatography-high-resolution mass spectrometry and Molecular Networking. Sci. Rep. 2018, 8, 17971. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, S.J.; Eom, J.S.; Choi, Y.; Jo, S.U.; Kim, J.; Lee, S.S.; Kim, E.T.; Lee, S.S. Comparison of metabolites in rumen fluid, urine, and feces of dairy cow from subacute ruminal acidosis model measured by proton nuclear magnetic resonance spectroscopy. Anim. Biosci. 2023, 36, 53–62. [Google Scholar] [CrossRef]
- Bica, R.; Palarea-Albaladejo, J.; Lima, J.; Uhrin, D.; Miller, G.A.; Bowen, J.M.; Pacheco, D.; Macrae, A.; Dewhurst, R.J. Methane emissions and rumen metabolite concentrations in cattle fed two different silages. Sci. Rep. 2022, 12, 5441. [Google Scholar] [CrossRef]
- O’Callaghan, T.F.; Vázquez-Fresno, R.; Serra-Cayuela, A.; Dong, E.; Mandal, R.; Hennessy, D.; McAuliffe, S.; Dillon, P.; Wishart, D.S.; Stanton, C.; et al. Pasture Feeding Changes the Bovine Rumen and Milk Metabolome. Metabolites 2018, 8, 27. [Google Scholar] [CrossRef]
- Saleem, F.; Ametaj, B.N.; Bouatra, S.; Mandal, R.; Zebeli, Q.; Dunn, S.M.; Wishart, D.S. A metabolomics approach to uncover the effects of grain diets on rumen health in dairy cows. J. Dairy Sci. 2012, 95, 6606–6623. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Zebeli, Q.; Saleem, F.; Psychogios, N.; Lewis, M.J.; Dunn, S.M.; Xia, J.; Wishart, D.S. Metabolomics reveals unhealthy alterations in rumen metabolism with increased proportion of cereal grain in the diet of dairy cows. Metabolomics 2010, 6, 583–594. [Google Scholar] [CrossRef]
- Zhao, S.; Zhao, J.; Bu, D.; Sun, P.; Wang, J.; Dong, Z. Metabolomics analysis reveals large effect of roughage types on rumen microbial metabolic profile in dairy cows. Lett. Appl. Microbiol. 2014, 59, 79–85. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, W.; Jiang, L.; Mao, S. Comparative metabolome analysis of ruminal changes in Holstein dairy cows fed low- or high-concentrate diets. Metabolomics 2017, 13, 74. [Google Scholar] [CrossRef]
- Joubran, A.M.; Pierce, K.M.; Garvey, N.; Shalloo, L.; O’Callaghan, T.F. Invited review: A 2020 perspective on pasture-based dairy systems and products. J. Dairy Sci. 2021, 104, 7364–7382. [Google Scholar] [CrossRef]
- Timlin, M.; Fitzpatrick, E.; McCarthy, K.; Tobin, J.T.; Murphy, E.G.; Pierce, K.M.; Murphy, J.P.; Hennessy, D.; O’Donovan, M.; Harbourne, N.; et al. Impact of varying levels of pasture allowance on the nutritional quality and functionality of milk throughout lactation. J. Dairy Sci. 2023, 106, 6597–6622. [Google Scholar] [CrossRef]
- Fitzpatrick, E.; Gilliland, T.; Hennessy, D. Grassland at the heart of circular and sustainable food systems. In Proceedings of the 29th General Meeting of the European Grassland Federation, Caen, France, 26–30 June 2022; The Organising Committee of the 29th General Meeting of the European Grassland Federation. INRAE: Paris, France, 2022; Volume 27, pp. 572–574. [Google Scholar] [CrossRef]
- O’Connor, P.J.; Hennessy, D.; Lynch, M.B.; Slattery, H.; Lewis, E. The effect of dicyandiamide on rumen and blood metabolites, diet digestibility and urinary excretion. Livest. Sci. 2013, 155, 30–37. [Google Scholar] [CrossRef]
- Bica, R.; Palarea-Albaladejo, J.; Kew, W.; Uhrin, D.; Pacheco, D.; Macrae, A.; Dewhurst, R.J. Nuclear Magnetic Resonance to Detect Rumen Metabolites Associated with Enteric Methane Emissions from Beef Cattle. Sci. Rep. 2020, 10, 5578. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, M.P.; Lebeuf, Y.; Gervais, R.; Tremblay, G.F.; Vuillemard, J.C.; Fortin, J.; Chouinard, P.Y. Milk volatile organic compounds and fatty acid profile in cows fed timothy as hay, pasture, or silage. J. Dairy Sci. 2013, 96, 7181–7194. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.F.; Kiene, R.P. Microbial Metabolism of Dimethyl Sulfide. In Biogenic Sulfur in the Environment. ACS Symp. Ser. 1989, 393, 202–221. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, H.; Wang, Y.; Li, S.; Cao, Z.; Ji, S.; He, Y.; Zhang, H. Effect of Dietary Forage to Concentrate Ratios on Dynamic Profile Changes and Interactions of Ruminal Microbiota and Metabolites in Holstein Heifers. Front. Microbiol. 2017, 8, 2206. [Google Scholar] [CrossRef]
- Seymour, W.M.; Campbell, D.R.; Johnson, Z.B. Relationships between rumen volatile fatty acid concentrations and milk production in dairy cows: A literature study. Anim. Feed. Sci. Technol. 2005, 119, 155–169. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, H.; Xu, J.; Zhai, Y.; Xia, H.; Xi, Y.; Han, Z. Characteristics of ruminal microbiota and metabolome in Holstein cows differing in milk protein concentrations. J. Anim. Sci. 2022, 100, skac253. [Google Scholar] [CrossRef]
- Urrutia, N.; Bomberger, R.; Matamoros, C.; Harvatine, K.J. Effect of dietary supplementation of sodium acetate and calcium butyrate on milk fat synthesis in lactating dairy cows. J. Dairy Sci. 2019, 102, 5172–5181. [Google Scholar] [CrossRef]
- Mohan, M.S.; O’Callaghan, T.F.; Kelly, P.; Hogan, S.A. Milk fat: Opportunities, challenges and innovation. Crit. Rev. Food. Sci. Nutr. 2021, 61, 2411–2443. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef]
- Brask, M.; Weisbjerg, M.R.; Hellwing, A.L.F.; Bannink, A.; Lund, P. Methane production and diurnal variation measured in dairy cows and predicted from fermentation pattern and nutrient or carbon flow. Animals 2015, 9, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- van Lingen, H.J.; Edwards, J.E.; Vaidya, J.D.; van Gastelen, S.; Saccenti, E.; van den Bogert, B.; Bannink, A.; Smidt, H.; Plugge, C.M.; Dijkstra, J. Diurnal Dynamics of Gaseous and Dissolved Metabolites and Microbiota Composition in the Bovine Rumen. Front. Microbiol. 2017, 8, 425. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maize Silage | Grass Silage | Conc. | Parlour Conc. | Total TMR | Total PMR | |
|---|---|---|---|---|---|---|
| DM (%) | 33.70 | 30.71 | 93.87 | 89.00 | 58.11 | 41.54 |
| CP (g/kg DM) | 76.54 | 152.05 | 220.33 | 133.33 | 146.54 | 177.66 |
| NDF (g/kg DM) | 427.29 | 459.71 | 238.86 | 308.80 | 358.43 | 363.69 |
| Ash (g/kg DM) | 35.65 | 93.92 | 71.68 | 94.53 | 63.36 | 74.59 |
| Starch (%) | 31.19 | N/A | N/A | N/A | 12.06 | 6.59 |
| OM Digestibility (g/kg DM) | 760.17 | 739.00 | 849.00 | 840.6 | 792.90 | 815.66 |
| Gross energy (MJ/kg DM) | 19.22 | 18.64 | 19.57 | 18.49 | 19.19 | 19.20 |
| Pasture | PMR | |
|---|---|---|
| DM (%) | 208.31 | 221.18 |
| CP (g/kg DM) | 845.13 | 843.23 |
| NDF (g/kg DM) | 366.99 | 374.57 |
| Ash (g/kg DM) | 208.42 | 218.67 |
| Starch (%) | 85.39 | 87.64 |
| OM Digestibility (g/kg DM) | 19.22 | 19.26 |
| Gross energy (MJ/kg DM) | 208.31 | 221.18 |
| Metabolite AM | Pasture | PMR | TMR | FDR | |||
|---|---|---|---|---|---|---|---|
| 1,3-Dihydroxyacetone | 0.03 | (0.02) | 0.03 | (0.01) | 0.03 | (0.01) | <0.05 |
| 2-Hydroxy-3-methylvalerate | 0.07 | (0.02) a | 0.09 | (0.03) b | 0.09 | (0.03) b | <0.001 |
| 3-Hydroxybutyrate | 0.03 | (0.01) ab | 0.03 | (0.01) a | 0.03 | (0.01) b | <0.05 |
| 3-Hydroxyphenylacetate | 0.02 | (0.01) a | 0.03 | (0.01) b | 0.03 | (0.01) b | <0.001 |
| 3-Phenylpropionate | 0.51 | (0.09) a | 0.43 | (0.07) b | 0.44 | (0.08) b | <0.001 |
| 5-Phenylvaleric acid | 0.18 | (0.09) a | 0.20 | (0.08) ab | 0.23 | (0.08) b | <0.001 |
| Acetate | 66.37 | (3.05) a | 62.90 | (5.82) b | 64.18 | (3.48) b | <0.001 |
| Acetoacetate | 0.03 | (0.01) | 0.03 | (0.01) | 0.03 | (0.01) | 0.0.095 |
| Acetone | 0.03 | (0.04) | 0.03 | (0.02) | 0.04 | (0.03) | 0.610 |
| Alanine | 0.17 | (0.06) a | 0.19 | (0.04) ab | 0.20 | (0.05) b | <0.01 |
| Aspartate | 0.10 | (0.05) a | 0.12 | (0.04) b | 0.11 | (0.04) ab | <0.01 |
| Benzoate | 0.04 | (0.03) | 0.03 | (0.01) | 0.04 | (0.02) | <0.05 |
| Butyrate | 10.50 | (1.10) a | 8.99 | (1.19) b | 9.41 | (1.84) b | <0.001 |
| Cadaverine | 0.05 | (0.02) a | 0.05 | (0.02) ab | 0.06 | (0.02) b | <0.01 |
| Choline | 0.02 | (0.01) a | 0.01 | (0.00) b | 0.02 | (0.01) ab | <0.05 |
| Dimethyl sulfone | 0.02 | (0.01) a | 0.02 | (0.00) b | 0.01 | (0.00) c | <0.001 |
| Dimethylamine | 0.01 | (0.01) | 0.01 | (0.01) | 0.01 | (0.01) | 0.498 |
| Ethanol | 0.10 | (0.08) | 0.12 | (0.09) | 0.10 | (0.04) | 0.433 |
| Formate | 0.09 | (0.03) a | 0.09 | (0.02) ab | 0.07 | (0.02) b | <0.01 |
| Fumarate | 0.01 | (0.01) a | 0.01 | (0.00) ab | 0.01 | (0.00) b | <0.05 |
| Glucose | 0.44 | (0.34) a | 0.44 | (0.33) ab | 0.75 | (0.44) b | <0.001 |
| Glutamate | 0.26 | (0.07) a | 0.43 | (0.13) b | 0.35 | (0.14) c | <0.001 |
| Glycerol | 0.06 | (0.04) | 0.06 | (0.02) | 0.06 | (0.02) | 0.216 |
| Glycine | 0.06 | (0.02) | 0.07 | (0.02) | 0.07 | (0.02) | 0.181 |
| Histidine | 0.02 | (0.01) | 0.02 | (0.01) | 0.02 | (0.01) | 0.528 |
| Hypoxanthine | 0.07 | (0.03) | 0.06 | (0.02) | 0.07 | (0.03) | 0.139 |
| Imidazole | 0.03 | (0.02) | 0.03 | (0.01) | 0.03 | (0.01) | 0.098 |
| Isobutyrate | 0.96 | (0.20) a | 0.86 | (0.09) b | 0.98 | (0.17) ab | <0.001 |
| Isoleucine | 0.04 | (0.02) | 0.04 | (0.01) | 0.04 | (0.01) | 0.056 |
| Isopropanol | 0.03 | (0.01) a | 0.05 | (0.03) b | 0.04 | (0.02) ab | <0.001 |
| Isovalerate | 0.61 | (0.17) a | 0.54 | (0.08) b | 0.57 | (0.11) ab | <0.05 |
| Lactate | 0.07 | (0.07) | 0.07 | (0.03) | 0.07 | (0.02) | 0.941 |
| Leucine | 0.05 | (0.02) | 0.05 | (0.01) | 0.05 | (0.01) | 0.289 |
| Lysine | 0.07 | (0.03) | 0.07 | (0.03) | 0.08 | (0.02) | 0.941 |
| Malonic acid | 0.02 | (0.01) a | 0.02 | (0.01) ab | 0.02 | (0.01) b | <0.001 |
| Maltose | 0.09 | (0.07) a | 0.10 | (0.06) a | 0.22 | (0.13) b | <0.001 |
| Methanol | 0.01 | (0.02) | 0.01 | (0.01) | 0.01 | (0.01) | 0.611 |
| Methylamine | 0.03 | (0.03) | 0.02 | (0.03) | 0.02 | (0.02) | 0.277 |
| N-Acetylglutamic acid | 0.20 | (0.08) a | 0.23 | (0.08) a | 0.27 | (0.07) b | <0.001 |
| Phenylacetate | 0.25 | (0.14) | 0.20 | (0.09) | 0.20 | (0.11) | 0.052 |
| Proline | 0.12 | (0.07) | 0.12 | (0.04) | 0.12 | (0.04) | 0.871 |
| Propionate | 16.10 | (2.22) a | 20.23 | (4.26) b | 18.19 | (3.30) c | <0.001 |
| Propylene glycol | 0.03 | (0.02) a | 0.04 | (0.03) ab | 0.04 | (0.03) b | <0.01 |
| Ribose | 0.24 | (0.15) ab | 0.22 | (0.10) a | 0.29 | (0.12) b | <0.05 |
| Succinate | 0.05 | (0.04) ab | 0.05 | (0.03) a | 0.04 | (0.03) b | 0.064 |
| Trimethylamine | 0.03 | (0.04) a | 0.03 | (0.05) ab | 0.02 | (0.02) b | 0.181 |
| Tyrosine | 0.04 | (0.02) | 0.05 | (0.02) | 0.05 | (0.01) | <0.05 |
| Uracil | 0.07 | (0.03) | 0.07 | (0.03) | 0.08 | (0.03) | 0.155 |
| Valerate | 1.48 | (0.76) a | 2.38 | (1.50) b | 2.06 | (0.82) b | <0.001 |
| Valine | 0.06 | (0.02) | 0.06 | (0.02) | 0.06 | (0.02) | 0.575 |
| Metabolite AM | Pasture | PMR | TMR | FDR | |||
|---|---|---|---|---|---|---|---|
| 1,3-Dihydroxyacetone | 0.02 | (0.01) a | 0.03 | (0.01) b | 0.03 | (0.01) b | <0.001 |
| 2-Hydroxy-3-methylvalerate | 0.09 | (0.02) a | 0.10 | (0.03) ab | 0.10 | (0.04) b | <0.05 |
| 3-Hydroxybutyrate | 0.05 | (0.02) a | 0.05 | (0.01) b | 0.04 | (0.01) c | <0.001 |
| 3-Hydroxyphenylacetate | 0.06 | (0.04) a | 0.06 | (0.02) a | 0.03 | (0.02) b | <0.001 |
| 3-Phenylpropionate | 0.83 | (0.20) a | 0.85 | (0.13) a | 0.37 | (0.11) b | <0.001 |
| 5-Phenylvaleric acid | 0.22 | (0.09) a | 0.22 | (0.08) a | 0.27 | (0.11) b | <0.001 |
| Acetate | 61.04 | (4.46) a | 61.11 | (3.20) a | 58.75 | (3.92) b | <0.01 |
| Acetoacetate | 0.03 | (0.01) a | 0.03 | (0.01) a | 0.04 | (0.02) b | <0.001 |
| Acetone | 0.07 | (0.08) a | 0.04 | (0.02) b | 0.09 | (0.09) a | <0.001 |
| Alanine | 0.18 | (0.06) a | 0.20 | (0.06) a | 0.32 | (0.27) b | <0.001 |
| Aspartate | 0.10 | (0.04) | 0.10 | (0.04) | 0.10 | (0.05) | 0.225 |
| Benzoate | 0.06 | (0.02) ab | 0.07 | (0.08) a | 0.04 | (0.02) b | <0.05 |
| Butyrate | 11.53 | (1.47) a | 10.01 | (2.13) b | 10.27 | (1.96) b | <0.001 |
| Cadaverine | 0.07 | (0.04) a | 0.07 | (0.02) a | 0.10 | (0.03) b | <0.001 |
| Choline | 0.02 | (0.01) | 0.02 | (0.01) | 0.02 | (0.01) | 0.074 |
| Dimethyl sulfone | 0.02 | (0.01) a | 0.01 | (0.00) b | 0.02 | (0.00) b | <0.001 |
| Dimethylamine | 0.03 | (0.02) | 0.03 | (0.03) | 0.03 | (0.03) | 0.385 |
| Ethanol | 0.32 | (0.41) a | 0.22 | (0.21) a | 0.93 | (1.05) b | <0.001 |
| Formate | 0.06 | (0.02) ab | 0.06 | (0.02) a | 0.05 | (0.02) b | <0.001 |
| Fumarate | 0.01 | (0.01) | 0.01 | (0.00) | 0.01 | (0.01) | 0.381 |
| Glucose | 0.88 | (0.46) a | 0.66 | (0.32) a | 1.61 | (0.70) b | <0.001 |
| Glutamate | 0.20 | (0.05) a | 0.27 | (0.09) b | 0.25 | (0.10) b | <0.001 |
| Glycerol | 0.07 | (0.03) a | 0.07 | (0.02) a | 0.10 | (0.05) b | <0.001 |
| Glycine | 0.10 | (0.06) a | 0.07 | (0.02) a | 0.18 | (0.12) b | <0.001 |
| Histidine | 0.03 | (0.01) | 0.02 | (0.01) | 0.02 | (0.01) | 0.257 |
| Hypoxanthine | 0.09 | (0.03) ab | 0.08 | (0.03) a | 0.10 | (0.03) b | <0.01 |
| Imidazole | 0.04 | (0.02) | 0.04 | (0.02) | 0.04 | (0.02) | 0.257 |
| Isobutyrate | 0.87 | (0.21) | 0.87 | (0.17) | 0.89 | (0.14) | 0.712 |
| Isoleucine | 0.06 | (0.02) a | 0.04 | (0.01) b | 0.08 | (0.03) c | <0.001 |
| Isopropanol | 0.05 | (0.02) ab | 0.04 | (0.01) a | 0.06 | (0.03) b | <0.01 |
| Isovalerate | 0.57 | (0.16) | 0.54 | (0.11) | 0.54 | (0.12) | 0.434 |
| Lactate | 0.08 | (0.03) | 0.07 | (0.02) | 0.08 | (0.04) | 0.113 |
| Leucine | 0.05 | (0.02) a | 0.06 | (0.02) a | 0.07 | (0.04) b | <0.01 |
| Lysine | 0.10 | (0.05) a | 0.08 | (0.04) a | 0.13 | (0.07) b | <0.001 |
| Malonic acid | 0.02 | (0.01) a | 0.02 | (0.01) a | 0.03 | (0.01) b | <0.001 |
| Maltose | 0.18 | (0.09) a | 0.16 | (0.10) a | 0.61 | (0.23) b | <0.001 |
| Methanol | 0.02 | (0.02) a | 0.02 | (0.01) a | 0.03 | (0.03) b | <0.01 |
| Methylamine | 0.17 | (0.11) ab | 0.19 | (0.14) a | 0.13 | (0.08) b | <0.05 |
| N-Acetylglutamic acid | 0.26 | (0.08) a | 0.27 | (0.07) a | 0.31 | (0.05) b | <0.01 |
| Phenylacetate | 0.32 | (0.18) a | 0.28 | (0.11) a | 0.21 | (0.08) b | <0.001 |
| Proline | 0.11 | (0.05) | 0.11 | (0.04) | 0.12 | (0.05) | 0.090 |
| Propionate | 18.43 | (2.87) a | 19.87 | (2.09) b | 19.24 | (3.42) | <0.05 |
| Propylene glycol | 0.04 | (0.02) a | 0.04 | (0.01) a | 0.10 | (0.06) b | <0.001 |
| Ribose | 0.27 | (0.11) a | 0.28 | (0.09) a | 0.34 | (0.13) b | <0.01 |
| Succinate | 0.02 | (0.02) | 0.02 | (0.01) | 0.01 | (0.01) | 0.101 |
| Trimethylamine | 0.11 | (0.14) | 0.06 | (0.06) | 0.10 | (0.13) | 0.080 |
| Tyrosine | 0.05 | (0.02) a | 0.05 | (0.02) a | 0.06 | (0.02) b | <0.001 |
| Uracil | 0.10 | (0.03) | 0.09 | (0.03) | 0.11 | (0.04) | <0.05 |
| Valerate | 1.88 | (0.65) a | 2.30 | (1.10) a | 2.77 | (1.00) b | <0.001 |
| Valine | 0.06 | (0.02) a | 0.06 | (0.02) a | 0.09 | (0.08) b | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Connolly, C.; Timlin, M.; Hogan, S.A.; O’Callaghan, T.F.; Brodkorb, A.; O’Donovan, M.; Hennessy, D.; Fitzpatrick, E.; McCarthy, K.; Murphy, J.P.; et al. The Impact of Varying Pasture Levels on the Metabolomic Profile of Bovine Ruminal Fluid. Metabolites 2024, 14, 476. https://doi.org/10.3390/metabo14090476
Connolly C, Timlin M, Hogan SA, O’Callaghan TF, Brodkorb A, O’Donovan M, Hennessy D, Fitzpatrick E, McCarthy K, Murphy JP, et al. The Impact of Varying Pasture Levels on the Metabolomic Profile of Bovine Ruminal Fluid. Metabolites. 2024; 14(9):476. https://doi.org/10.3390/metabo14090476
Chicago/Turabian StyleConnolly, Claire, Mark Timlin, Sean A. Hogan, Tom F. O’Callaghan, André Brodkorb, Michael O’Donovan, Deirdre Hennessy, Ellen Fitzpatrick, Kieran McCarthy, John P. Murphy, and et al. 2024. "The Impact of Varying Pasture Levels on the Metabolomic Profile of Bovine Ruminal Fluid" Metabolites 14, no. 9: 476. https://doi.org/10.3390/metabo14090476
APA StyleConnolly, C., Timlin, M., Hogan, S. A., O’Callaghan, T. F., Brodkorb, A., O’Donovan, M., Hennessy, D., Fitzpatrick, E., McCarthy, K., Murphy, J. P., & Brennan, L. (2024). The Impact of Varying Pasture Levels on the Metabolomic Profile of Bovine Ruminal Fluid. Metabolites, 14(9), 476. https://doi.org/10.3390/metabo14090476