Quenching for Microalgal Metabolomics: A Case Study on the Unicellular Eukaryotic Green Alga Chlamydomonas reinhardtii



Abstract
:1. Introduction
2. Results and Discussion
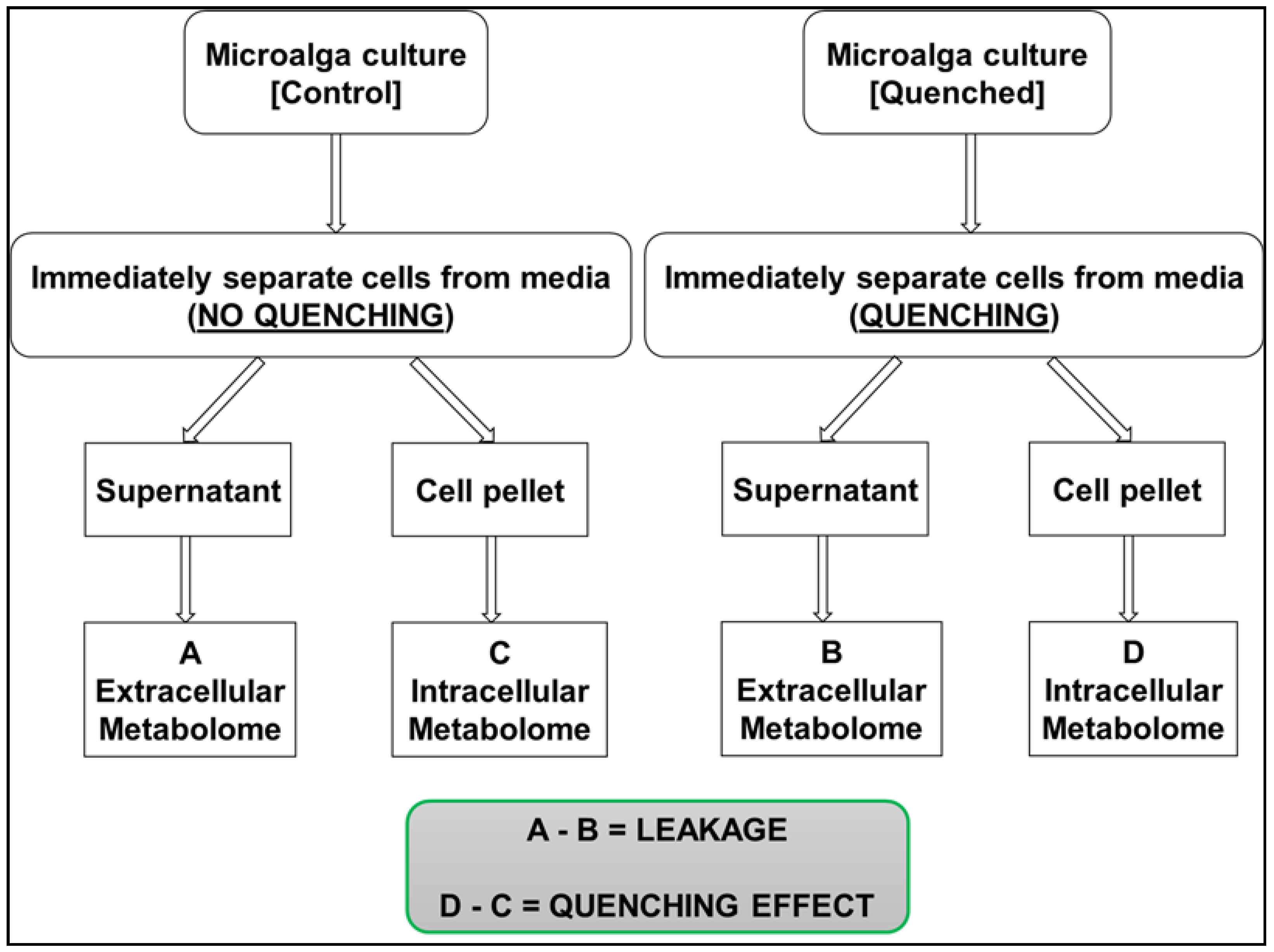
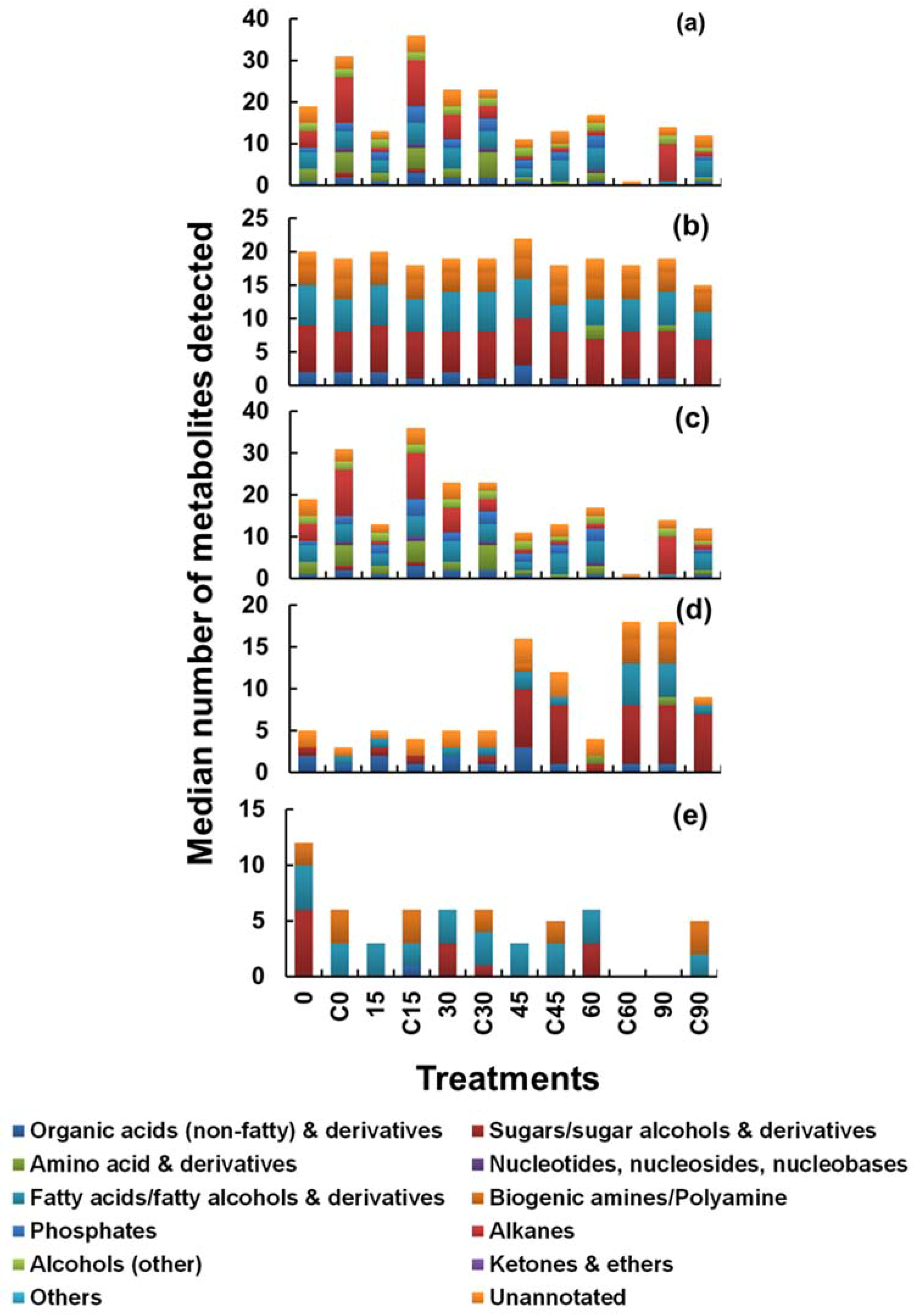
- Response variable 1: Median number of metabolites recovered in;
- (a)
- Cell extracts
- (b)
- In quenching supernatant
- (c)
- Only in cell extract and not in supernatant (unique to cells)
- (d)
- Only in supernatant and not in cell extract (unique to supernatant) and
- (e)
- In both the cell extracts and supernatants (common to both)
- Response variable 2: Recoveries of metabolites within twelve different classes of metabolites with respect to their normalised median peak intensities, and represented by the average metabolite peak intensity for each class.
2.1. Leakage Based on Recovery of Median Metabolite Numbers
2.1.1. Effect of Methanol Concentration and Inclusion of Buffer Additives on Metabolite Leakage (Approach 1)
2.1.2. Effect of Prolonged Exposure to Quenching Solvent on Metabolite Leakage (Approach 2)
2.1.3. Effect of Quenching Solvent to Culture Ratio (Temperature Influence) (Approach 3)
2.2. Leakage Based on Metabolite Peak Intensities
3. Materials and Methods
3.1. Chemicals and Analytical Reagents
3.2. Microalgal Cultivation
3.3. Sampling and Quenching
3.4. Metabolite Extraction
3.5. Metabolite Derivatization and GC-MS Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kapoore, R.V.; Coyle, R.; Staton, C.A.; Brown, N.J.; Vaidyanathan, S. Cell line dependence of metabolite leakage in metabolome analyses of adherent normal and cancer cell lines. Metabolomics 2015, 11, 1743–1755. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Vaidyanathan, S. Towards quantitative mass spectrometry-based metabolomics in microbial and mammalian systems. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, B.; François, J.; Renaud, M. A rapid and reliable method for metabolite extraction in yeast using boiling buffered ethanol. Yeast 1997, 13, 1347–1355. [Google Scholar] [CrossRef] [Green Version]
- Bolten, C.J.; Kiefer, P.; Letisse, F.; Portais, J.-C.; Wittmann, C. Sampling for metabolome analysis of microorganisms. Anal. Chem. 2007, 79, 3843–3849. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Bruheim, P. Cold glycerol–saline: The promising quenching solution for accurate intracellular metabolite analysis of microbial cells. Anal. Biochem. 2007, 370, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Højer-Pedersen, J.; Åkesson, M.; Smedsgaard, J.; Nielsen, J. Global metabolite analysis of yeast: Evaluation of sample preparation methods. Yeast 2005, 22, 1155–1169. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, C.; Krömer, J.O.; Kiefer, P.; Binz, T.; Heinzle, E. Impact of the cold shock phenomenon on quantification of intracellular metabolites in bacteria. Anal. Biochem. 2004, 327, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.D. Cellular metabolomics of Escherchia coli. Expert Rev. Proteom. 2007, 4, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Canelas, A.B.; Ras, C.; ten Pierick, A.; van Dam, J.C.; Heijnen, J.J.; Van Gulik, W.M. Leakage-free rapid quenching technique for yeast metabolomics. Metabolomics 2008, 4, 226–239. [Google Scholar] [CrossRef] [Green Version]
- Buchholz, A.; Takors, R.; Wandrey, C. Quantification of Intracellular Metabolites in Escherichia coli K12 Using Liquid Chromatographic-Electrospray Ionization Tandem Mass Spectrometric Techniques. Anal. Biochem. 2001, 295, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Fatemeh, T.; Tang, L.; Cai, Z. Quantitative metabolic network profiling of Escherichia coli: An overview of analytical methods for measurement of intracellular metabolites. TrAC Trends Anal. Chem. 2016, 75, 141–150. [Google Scholar] [CrossRef]
- Pinu, F.R.; Villas-Boas, S.G.; Aggio, R.J.M. Analysis of Intracellular Metabolites from Microorganisms: Quenching and Extraction Protocols. Metabolites 2017, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Schädel, F.; David, F.; Franco-Lara, E. Evaluation of cell damage caused by cold sampling and quenching for metabolome analysis. Appl. Microbiol. Biotechnol. 2011, 92, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Spura, J.; Christian Reimer, L.; Wieloch, P.; Schreiber, K.; Buchinger, S.; Schomburg, D. A method for enzyme quenching in microbial metabolome analysis successfully applied to gram-positive and gram-negative bacteria and yeast. Anal. Biochem. 2009, 394, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Nambou, K.; Wei, L.; Chen, J.; Imanaka, T.; Hua, Q. Evaluation of metabolome sample preparation methods regarding leakage reduction for the oleaginous yeast Yarrowia lipolytica. Biochem. Eng. J. 2014, 82, 63–70. [Google Scholar] [CrossRef]
- Wellerdiek, M.; Winterhoff, D.; Reule, W.; Brandner, J.; Oldiges, M. Metabolic quenching of Corynebacterium glutamicum: Efficiency of methods and impact of cold shock. Bioprocess Biosyst. Eng. 2009, 32, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Faijes, M.; Mars, A.E.; Smid, E.J. Comparison of quenching and extraction methodologies for metabolome analysis of Lactobacillus plantarum. Microb. Cell Fact. 2007, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Kronthaler, J.; Gstraunthaler, G.; Heel, C. Optimizing high-throughput metabolomic biomarker screening: A study of quenching solutions to freeze intracellular metabolism in CHO cells. Omics J. Integr. Boil. 2012, 16, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Sellick, C.A.; Hansen, R.; Maqsood, A.R.; Dunn, W.B.; Stephens, G.M.; Goodacre, R.; Dickson, A.J. Effective quenching processes for physiologically valid metabolite profiling of suspension cultured mammalian cells. Anal. Chem. 2008, 81, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Tredwell, G.D.; Edwards-Jones, B.; Leak, D.J.; Bundy, J.G. The development of metabolomic sampling procedures for Pichia pastoris, and baseline metabolome data. PLoS ONE 2011, 6, e16286. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, L.P.; Douma, R.D.; Heijnen, J.J.; van Gulik, W.M. Optimization of cold methanol quenching for quantitative metabolomics of Penicillium chrysogenum. Metabolomics 2012, 8, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, D.Y.; Wohlgemuth, G.; Park, H.S.; Fiehn, O.; Kim, K.H. Evaluation and optimization of metabolome sample preparation methods for Saccharomyces cerevisiae. Anal. Chem. 2013, 85, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Fiehn, O. High quality metabolomic data for Chlamydomonas reinhardtii. Plant Methods 2008, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Bölling, C.; Fiehn, O. Metabolite profiling of Chlamydomonas reinhardtii under nutrient deprivation. Plant Physiol. 2005, 139, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Kapoore, R.V.; Coyle, R.; Staton, C.A.; Brown, N.J.; Vaidyanathan, S. Influence of washing and quenching in profiling the metabolome of adherent mammalian cells: A case study with the metastatic breast cancer cell line MDA-MB-231. Analyst 2017, 142, 2038–2049. [Google Scholar] [CrossRef] [PubMed]
- Koning, W.d.; Dam, K.v. A method for the determination of changes of glycolytic metabolites in yeast on a subsecond time scale using extraction at neutral pH. Anal. Biochem. 1992, 204, 118–123. [Google Scholar] [CrossRef]
- Weuster-Botz, D. Die Rolle der Reaktionstechnik in der Mikrobiellen Verfahrensentwicklung; Forschungszentrum Jülich, Zentralbibliothek: Jülich, Germany, 1999. [Google Scholar]
- Britten, R.J.; McClure, F.T. The amino acid pool in Escherichia coli. Bacteriol. Rev. 1962, 26, 292. [Google Scholar] [PubMed]
- Ramautar, R.; Somsen, G.W.; de Jong, G.J. CE-MS in metabolomics. Electrophoresis 2009, 30, 276–291. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Ohashi, Y.; Ueno, Y.; Naraoka, H.; Tomita, M.; Nishioka, T. Quantitative metabolome analysis using capillary electrophoresis mass spectrometry. J. Proteome Res. 2003, 2, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, J.; Jonsson, P.; Nordström, A.; Sjöström, M.; Moritz, T. Design of experiments: An efficient strategy to identify factors influencing extraction and derivatization of Arabidopsis thaliana samples in metabolomic studies with gas chromatography/mass spectrometry. Anal. Biochem. 2004, 331, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W.; Wenzel, K.; Fiehn, O. Process for the integrated extraction, identification and quantification of metabolites, proteins and RNA to reveal their co-regulation in biochemical networks. Proteomics 2004, 4, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Kapoore, R.V. Mass Spectrometry Based Hyphenated Techniques for Microalgal and Mammalian Metabolomics. Ph.D. Thesis, University of Sheffield, Sheffield, UK, 2014. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method Code | Addition of Buffer | Initial Quenching Solvent Temperature (°C) | Initial Concentration of Methanol (v/v) (%) | Sample/Quenching Solvent Ratio | Final Concentration of Methanol after Quenching (v/v) (%) | Final Temperature of Resultant Mixture after Quenching (°C) | |
|---|---|---|---|---|---|---|---|
| Approach 1 | 33M | x | −50 | 33 | 1:1 | 17 | −20 |
| 33A | 0.85% AMBIC | −50 | 33 | 1:1 | 17 | −20 | |
| 60M | x | −50 | 60 | 1:1 | 30 | −30 | |
| 60A | 0.85% AMBIC | −50 | 60 | 1:1 | 30 | −30 | |
| 60H10 | 10 mM HEPES | −50 | 60 | 1:1 | 30 | −30 | |
| 60H70 | 70 mM HEPES | −50 | 60 | 1:1 | 30 | −30 | |
| 70M | x | −50 | 70 | 1:1 | 35 | −35 | |
| 100M | x | −50 | 100 | 1:1 | 50 | −40 | |
| Approach 3 | 60–41 | x | −50 | 60 | 1:4 | 48 | −40 |
| 100–21 | x | −50 | 100 | 1:2 | 77 | −45 | |
| Controls | Control (Unquenched) | x | x | x | x | x | x |
| Media | x | x | x | x | x | x |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapoore, R.V.; Vaidyanathan, S. Quenching for Microalgal Metabolomics: A Case Study on the Unicellular Eukaryotic Green Alga Chlamydomonas reinhardtii. Metabolites 2018, 8, 72. https://doi.org/10.3390/metabo8040072
Kapoore RV, Vaidyanathan S. Quenching for Microalgal Metabolomics: A Case Study on the Unicellular Eukaryotic Green Alga Chlamydomonas reinhardtii. Metabolites. 2018; 8(4):72. https://doi.org/10.3390/metabo8040072
Chicago/Turabian StyleKapoore, Rahul Vijay, and Seetharaman Vaidyanathan. 2018. "Quenching for Microalgal Metabolomics: A Case Study on the Unicellular Eukaryotic Green Alga Chlamydomonas reinhardtii" Metabolites 8, no. 4: 72. https://doi.org/10.3390/metabo8040072
APA StyleKapoore, R. V., & Vaidyanathan, S. (2018). Quenching for Microalgal Metabolomics: A Case Study on the Unicellular Eukaryotic Green Alga Chlamydomonas reinhardtii. Metabolites, 8(4), 72. https://doi.org/10.3390/metabo8040072