The Metabolome and Osteoarthritis: Possible Contributions to Symptoms and Pathology

Abstract
:1. Introduction
2. The Local and Systemic Metabolomes of Osteoarthritis
3. Osteoarthritis Phenotypes and Impact on Metabolome
3.1. Pain
3.2. Muscle Strength
3.3. Obesity
3.4. Depression
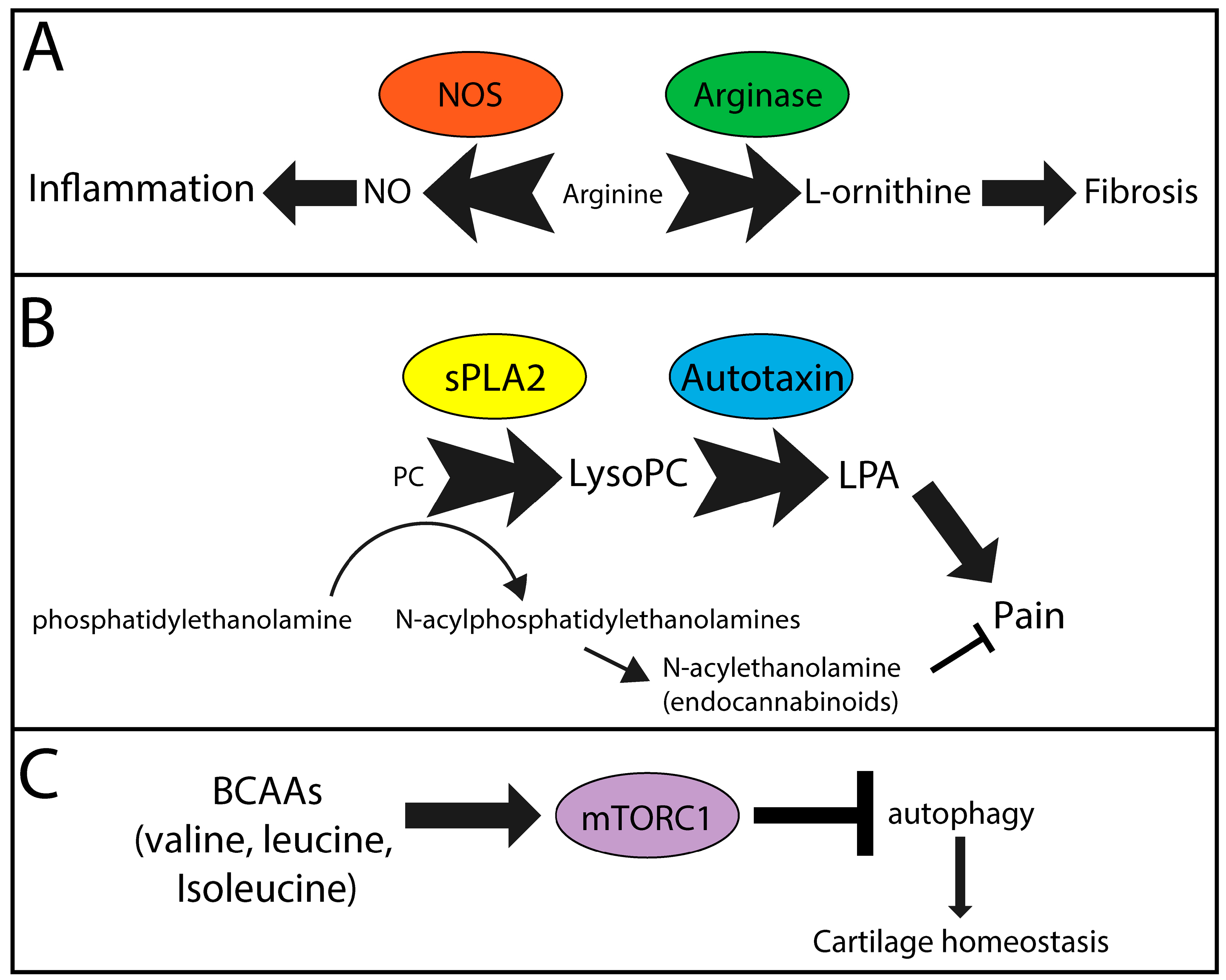
4. Metabolites and Pathways Likely Contributing to Osteoarthritis
4.1. PC-lysoPC-LPA
4.2. BCAA-mTOR
4.3. Arginine-NO/l-ornithine
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beswick, A.D.; Wylde, V.; Gooberman-Hill, R.; Blom, A.; Dieppe, P. What proportion of patients report long-term pain after total hip or knee replacement for osteoarthritis? A systematic review of prospective studies in unselected patients. BMJ Open 2012, 2, e000435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maradit Kremers, H.; Larson, D.R.; Crowson, C.S.; Kremers, W.K.; Washington, R.E.; Steiner, C.A.; Jiranek, W.A.; Berry, D.J. Prevalence of Total Hip and Knee Replacement in the United States. J. Bone Joint Surg. Am. 2015, 97, 1386–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.A.; Lewallen, D.G. Time trends in the characteristics of patients undergoing primary total knee arthroplasty. Arthrit. Care Res. 2014, 66, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Oliveria, S.A.; Felson, D.T.; Reed, J.I.; Cirillo, P.A.; Walker, A.M. Incidence of symptomatic hand, hip, and knee osteoarthritis among patients in a health maintenance organization. Arthritis Rheum. 1995, 38, 1134–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apold, H.; Meyer, H.E.; Nordsletten, L.; Furnes, O.; Baste, V.; Flugsrud, G.B. Risk factors for knee replacement due to primary osteoarthritis, a population based, prospective cohort study of 315,495 individuals. BMC Musculoskelet. Disord. 2014, 15, 217. [Google Scholar] [CrossRef]
- Silverwood, V.; Blagojevic-Bucknall, M.; Jinks, C.; Jordan, J.L.; Protheroe, J.; Jordan, K.P. Current evidence on risk factors for knee osteoarthritis in older adults: A systematic review and meta-analysis. Osteoarthritis Cartilage 2015, 23, 507–515. [Google Scholar] [CrossRef]
- Eymard, F.; Parsons, C.; Edwards, M.H.; Petit-Dop, F.; Reginster, J.Y.; Bruyere, O.; Richette, P.; Cooper, C.; Chevalier, X. Diabetes is a risk factor for knee osteoarthritis progression. Osteoarthritis Cartilage 2015, 23, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.F.; London, D.A.; Husni, E.M.; Navaneethan, S.; Kashyap, S.R. Type 2 diabetes and osteoarthritis: A systematic review and meta-analysis. J. Diabetes Complicat. 2016, 30, 944–950. [Google Scholar] [CrossRef]
- van der Esch, M.; Knoop, J.; van der Leeden, M.; Roorda, L.D.; Lems, W.F.; Knol, D.L.; Dekker, J. Clinical phenotypes in patients with knee osteoarthritis: A study in the Amsterdam osteoarthritis cohort. Osteoarthritis Cartilage 2015, 23, 544–549. [Google Scholar] [CrossRef]
- Dell’Isola, A.; Allan, R.; Smith, S.L.; Marreiros, S.S.; Steultjens, M. Identification of clinical phenotypes in knee osteoarthritis: A systematic review of the literature. BMC Musculoskelet. Disord. 2016, 17, 425. [Google Scholar] [CrossRef] [PubMed]
- Knoop, J.; van der Leeden, M.; Thorstensson, C.A.; Roorda, L.D.; Lems, W.F.; Knol, D.L.; Steultjens, M.P.; Dekker, J. Identification of phenotypes with different clinical outcomes in knee osteoarthritis: Data from the Osteoarthritis Initiative. Arthrit. Care Res. 2011, 63, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Jaggard, M.K.J.; Boulange, C.L.; Akhbari, P.; Vaghela, U.; Bhattacharya, R.; Williams, H.R.T.; Lindon, J.C.; Gupte, C.M. A systematic review of the small molecule studies of osteoarthritis using nuclear magnetic resonance and mass spectroscopy. Osteoarthritis Cartilage 2018. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.R.; Phelan, M.M.; Clegg, P.D.; Peffers, M.J.; Rubio-Martinez, L.M. Synovial Fluid Metabolites Differentiate between Septic and Nonseptic Joint Pathologies. J. Proteome Res. 2018, 17, 2735–2743. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.K.; Rawle, R.A.; Adams, E.; Greenwood, M.C.; Bothner, B.; June, R.K. Application of global metabolomic profiling of synovial fluid for osteoarthritis biomarkers. Biochem. Biophys. Res. Commun. 2018, 499, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Hinata, M.; Imai, S.; Sanaki, T.; Tsuchida, J.; Yoshioka, T.; Higashino, K.; Yamamoto, M.; Imai, M.; Soga, M.; Horita, N.; et al. Sensitization of transient receptor potential vanilloid 4 and increasing its endogenous ligand 5,6-epoxyeicosatrienoic acid in rats with monoiodoacetate-induced osteoarthritis. Pain 2018, 159, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sun, G.; Aitken, D.; Likhodii, S.; Liu, M.; Martin, G.; Furey, A.; Randell, E.; Rahman, P.; Jones, G.; et al. Lysophosphatidylcholines to phosphatidylcholines ratio predicts advanced knee osteoarthritis. Rheumatology (Oxford) 2016, 55, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Woo, D.C.; Jahng, G.H. In vivo H1 MR spectroscopy using 3 Tesla to investigate the metabolic profiles of joint fluids in different types of knee diseases. J. Appl. Clin. Med. Phys. 2016, 17, 561–572. [Google Scholar] [CrossRef] [Green Version]
- Loeser, R.F.; Pathmasiri, W.; Sumner, S.J.; McRitchie, S.; Beavers, D.; Saxena, P.; Nicklas, B.J.; Jordan, J.; Guermazi, A.; Hunter, D.J.; et al. Association of urinary metabolites with radiographic progression of knee osteoarthritis in overweight and obese adults: An exploratory study. Osteoarthritis Cartilage 2016, 24, 1479–1486. [Google Scholar] [CrossRef]
- Mickiewicz, B.; Shin, S.Y.; Pozzi, A.; Vogel, H.J.; Clark, A.L. Serum Metabolite Profiles Are Altered by Erlotinib Treatment and the Integrin alpha1-Null Genotype but Not by Post-Traumatic Osteoarthritis. J. Proteome Res. 2016, 15, 815–825. [Google Scholar] [CrossRef]
- Hu, T.; Zhang, W.; Fan, Z.; Sun, G.; Likhodi, S.; Randell, E.; Zhai, G. Metabolomics Differential Correlation Network Analysis of Osteoarthritis. Proc. Pac. Symp. 2016, 21, 120–131. [Google Scholar]
- Zhang, W.; Sun, G.; Likhodii, S.; Liu, M.; Aref-Eshghi, E.; Harper, P.E.; Martin, G.; Furey, A.; Green, R.; Randell, E.; et al. Metabolomic analysis of human plasma reveals that arginine is depleted in knee osteoarthritis patients. Osteoarthritis Cartilage 2016, 24, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Tufts, L.; Shet Vishnudas, K.; Fu, E.; Kurhanewicz, J.; Ries, M.; Alliston, T.; Li, X. Correlating high-resolution magic angle spinning NMR spectroscopy and gene analysis in osteoarthritic cartilage. NMR Biomed. 2015, 28, 523–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Likhodii, S.; Aref-Eshghi, E.; Zhang, Y.; Harper, P.E.; Randell, E.; Green, R.; Martin, G.; Furey, A.; Sun, G.; et al. Relationship between blood plasma and synovial fluid metabolite concentrations in patients with osteoarthritis. J. Rheumatol. 2015, 42, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Wang-Sattler, R.; Hart, D.J.; Arden, N.K.; Hakim, A.J.; Illig, T.; Spector, T.D. Serum branched-chain amino acid to histidine ratio: A novel metabolomic biomarker of knee osteoarthritis. Ann. Rheum. Dis. 2010, 69, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.R.; Ribeiro, L.R.; Downey-Jones, M.; Needham, M.R.; Oakley, C.; Wardale, J. Ligands for retinoic acid receptors are elevated in osteoarthritis and may contribute to pathologic processes in the osteoarthritic joint. Arthritis Rheum. 2009, 60, 1722–1732. [Google Scholar] [CrossRef]
- Lamers, R.J.; van Nesselrooij, J.H.; Kraus, V.B.; Jordan, J.M.; Renner, J.B.; Dragomir, A.D.; Luta, G.; van der Greef, J.; DeGroot, J. Identification of an urinary metabolite profile associated with osteoarthritis. Osteoarthritis Cartilage 2005, 13, 762–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Whiteman, M.; Mattey, D.L.; Halliwell, B. Raised levels of F(2)-isoprostanes and prostaglandin F(2alpha) in different rheumatic diseases. Ann. Rheum. Dis. 2001, 60, 627–631. [Google Scholar] [CrossRef]
- Wang, X.; Hunter, D.; Xu, J.; Ding, C. Metabolic triggered inflammation in osteoarthritis. Osteoarthritis Cartilage 2015, 23, 22–30. [Google Scholar] [CrossRef]
- Kosinska, M.K.; Liebisch, G.; Lochnit, G.; Wilhelm, J.; Klein, H.; Kaesser, U.; Lasczkowski, G.; Rickert, M.; Schmitz, G.; Steinmeyer, J. Sphingolipids in human synovial fluid—A lipidomic study. PLoS ONE 2014, 9, e91769. [Google Scholar] [CrossRef]
- Mickiewicz, B.; Kelly, J.J.; Ludwig, T.E.; Weljie, A.M.; Wiley, J.P.; Schmidt, T.A.; Vogel, H.J. Metabolic analysis of knee synovial fluid as a potential diagnostic approach for osteoarthritis. J. Orthop. Res. 2015, 33, 1631–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, K.; Shen, N.; Chen, H.; Ni, S.; Zhang, T.; Hu, M.; Wang, J.; Sun, L.; Yang, X. Global and targeted metabolomics of synovial fluid discovers special osteoarthritis metabolites. J. Orthop. Res. 2017, 35, 1973–1981. [Google Scholar] [CrossRef] [PubMed]
- Mickiewicz, B.; Heard, B.J.; Chau, J.K.; Chung, M.; Hart, D.A.; Shrive, N.G.; Frank, C.B.; Vogel, H.J. Metabolic profiling of synovial fluid in a unilateral ovine model of anterior cruciate ligament reconstruction of the knee suggests biomarkers for early osteoarthritis. J. Orthop. Res. 2015, 33, 71–77. [Google Scholar] [CrossRef]
- Hugle, T.; Kovacs, H.; Heijnen, I.A.; Daikeler, T.; Baisch, U.; Hicks, J.M.; Valderrabano, V. Synovial fluid metabolomics in different forms of arthritis assessed by nuclear magnetic resonance spectroscopy. Clin. Exp. Rheumatol. 2012, 30, 240–245. [Google Scholar] [PubMed]
- Anderson, J.R.; Chokesuwattanaskul, S.; Phelan, M.M.; Welting, T.J.M.; Lian, L.Y.; Peffers, M.J.; Wright, H.L. 1H NMR Metabolomics Identifies Underlying Inflammatory Pathology in Osteoarthritis and Rheumatoid Arthritis Synovial Joints. J. Proteome Res. 2018. [Google Scholar] [CrossRef]
- Kang, K.Y.; Lee, S.H.; Jung, S.M.; Park, S.H.; Jung, B.H.; Ju, J.H. Downregulation of Tryptophan-related Metabolomic Profile in Rheumatoid Arthritis Synovial Fluid. J. Rheumatol. 2015, 42, 2003–2011. [Google Scholar] [CrossRef]
- Kellgren, J.H.; Lawrence, J.S. Radiological assessment of osteo-arthrosis. Ann. Rheum. Dis. 1957, 16, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hwang, J.; Kim, J.; Ahn, J.K.; Cha, H.S.; Kim, K.H. Metabolite profiles of synovial fluid change with the radiographic severity of knee osteoarthritis. Joint Bone Spine 2017, 84, 605–610. [Google Scholar] [CrossRef]
- Cicuttini, F.M.; Wluka, A.E. Osteoarthritis: Is OA a mechanical or systemic disease? Nat. Rev. Rheumatol. 2014, 10, 515–516. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.; Zhang, Y.; Parousis, A.; Sharma, A.; Rossomacha, E.; Endisha, H.; Wu, B.; Kacprzak, I.; Mahomed, N.N.; Gandhi, R.; et al. High-fat diet-induced acceleration of osteoarthritis is associated with a distinct and sustained plasma metabolite signature. Sci. Rep. 2017, 7, 8205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Liu, J.; Qin, X.J.; Chen, Y.Y.; Gao, J.R.; Meng, M.; Wang, Y.; Wang, T. Gas chromatography-time of flight/mass spectrometry-based metabonomics of changes in the urinary metabolic profile in osteoarthritic rats. Exp. Ther. Med. 2018, 15, 2777–2785. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Su, X.; Wang, N.; Li, Y.; Yin, H.; Li, L.; Li, L. Chemical Isotope Labeling LC-MS for Monitoring Disease Progression and Treatment in Animal Models: Plasma Metabolomics Study of Osteoarthritis Rat Model. Sci. Rep. 2017, 7, 40543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tootsi, K.; Kals, J.; Zilmer, M.; Paapstel, K.; Ottas, A.; Martson, A. Medium- and long-chain acylcarnitines are associated with osteoarthritis severity and arterial stiffness in end-stage osteoarthritis patients: A case-control study. Int. J. Rheum. Dis. 2018, 21, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Hawker, G.A.; Croxford, R.; Bierman, A.S.; Harvey, P.J.; Ravi, B.; Stanaitis, I.; Lipscombe, L.L. All-cause mortality and serious cardiovascular events in people with hip and knee osteoarthritis: A population based cohort study. PLoS ONE 2014, 9, e91286. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Shin, J.S.; Lee, J.; Lee, Y.J.; Kim, M.R.; Bae, Y.H.; Park, K.B.; Lee, E.J.; Kim, J.H.; Ha, I.H. Association between Knee Osteoarthritis, Cardiovascular Risk Factors, and the Framingham Risk Score in South Koreans: A Cross-Sectional Study. PLoS ONE 2016, 11, e0165325. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Kamper, S.J.; Wiggers, J.H.; O’Brien, K.M.; Lee, H.; Wolfenden, L.; Yoong, S.L.; Robson, E.; McAuley, J.H.; Hartvigsen, J.; et al. Musculoskeletal conditions may increase the risk of chronic disease: A systematic review and meta-analysis of cohort studies. BMC Med. 2018, 16, 167. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bai, J.; He, B.; Hu, X.; Liu, D. Osteoarthritis and the risk of cardiovascular disease: A meta-analysis of observational studies. Sci. Rep. 2016, 6, 39672. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kopec, J.A.; Anis, A.H.; Cibere, J.; Goldsmith, C.H. Risk of cardiovascular disease in patients with osteoarthritis: A prospective longitudinal study. Arthrit. Care Res. 2013, 65, 1951–1958. [Google Scholar] [CrossRef]
- Hall, A.J.; Stubbs, B.; Mamas, M.A.; Myint, P.K.; Smith, T.O. Association between osteoarthritis and cardiovascular disease: Systematic review and meta-analysis. Eur. J. Prev. Cardiol. 2016, 23, 938–946. [Google Scholar] [CrossRef]
- Rahman, M.M.; Cibere, J.; Anis, A.H.; Goldsmith, C.H.; Kopec, J.A. Risk of Type 2 Diabetes among Osteoarthritis Patients in a Prospective Longitudinal Study. Int. J. Rheumatol. 2014, 2014, 620920. [Google Scholar] [CrossRef]
- Neumann, J.; Hofmann, F.C.; Heilmeier, U.; Ashmeik, W.; Tang, K.; Gersing, A.S.; Schwaiger, B.J.; Nevitt, M.C.; Joseph, G.B.; Lane, N.E.; et al. Type 2 diabetes patients have accelerated cartilage matrix degeneration compared to diabetes free controls: Data from the Osteoarthritis Initiative. Osteoarthritis Cartilage 2018, 26, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Jungmann, P.M.; Kraus, M.S.; Alizai, H.; Nardo, L.; Baum, T.; Nevitt, M.C.; McCulloch, C.E.; Joseph, G.B.; Lynch, J.A.; Link, T.M. Association of metabolic risk factors with cartilage degradation assessed by T2 relaxation time at the knee: Data from the osteoarthritis initiative. Arthrit. Care Res. 2013, 65, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Kleyer, A.; Perricone, C.; Sahinbegovic, E.; Iagnocco, A.; Zwerina, J.; Lorenzini, R.; Aschenbrenner, F.; Berenbaum, F.; D’Agostino, M.A.; et al. Diabetes is an independent predictor for severe osteoarthritis: Results from a longitudinal cohort study. Diabetes Care 2013, 36, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, F. Diabetes-induced osteoarthritis: From a new paradigm to a new phenotype. Ann. Rheum. Dis. 2011, 70, 1354–1356. [Google Scholar] [CrossRef] [PubMed]
- Laakso, M.; Kuusisto, J. Insulin resistance and hyperglycaemia in cardiovascular disease development. Nat. Rev. Endocrinol. 2014, 10, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhai, G.; Singmann, P.; He, Y.; Xu, T.; Prehn, C.; Romisch-Margl, W.; Lattka, E.; Gieger, C.; Soranzo, N.; et al. Human serum metabolic profiles are age dependent. Aging Cell 2012, 11, 960–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, F.; Xu, M.; Bruno, R.S.; Ballard, K.D.; Zhu, J. Targeted High Performance Liquid Chromatography Tandem Mass Spectrometry-based Metabolomics differentiates metabolic syndrome from obesity. Exp. Biol. Med. (Maywood) 2017, 242, 773–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockel, J.S.; Zhang, W.; Shestopaloff, K.; Likhodii, S.; Sun, G.; Furey, A.; Randell, E.; Sundararajan, K.; Gandhi, R.; Zhai, G.; et al. A classification modeling approach for determining metabolite signatures in osteoarthritis. PLoS ONE 2018, 13, e0199618. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Likhodii, S.; Zhang, Y.; Aref-Eshghi, E.; Harper, P.E.; Randell, E.; Green, R.; Martin, G.; Furey, A.; Sun, G.; et al. Classification of osteoarthritis phenotypes by metabolomics analysis. BMJ Open 2014, 4, e006286. [Google Scholar] [CrossRef] [Green Version]
- Finco, G.; Locci, E.; Mura, P.; Massa, R.; Noto, A.; Musu, M.; Landoni, G.; d’Aloja, E.; De-Giorgio, F.; Scano, P.; et al. Can Urine Metabolomics Be Helpful in Differentiating Neuropathic and Nociceptive Pain? A Proof-of-Concept Study. PLoS ONE 2016, 11, e0150476. [Google Scholar] [CrossRef]
- Hadrevi, J.; Bjorklund, M.; Kosek, E.; Hallgren, S.; Antti, H.; Fahlstrom, M.; Hellstrom, F. Systemic differences in serum metabolome: A cross sectional comparison of women with localised and widespread pain and controls. Sci. Rep. 2015, 5, 15925. [Google Scholar] [CrossRef] [PubMed]
- Um, S.Y.; Chung, M.W.; Kim, K.B.; Kim, S.H.; Oh, J.S.; Oh, H.Y.; Lee, H.J.; Choi, K.H. Pattern recognition analysis for the prediction of adverse effects by nonsteroidal anti-inflammatory drugs using 1H NMR-based metabolomics in rats. Anal. Chem. 2009, 81, 4734–4741. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.K.; Yadav, R.; Mukherjee, S.; Sinha, N. Perturbation of muscle metabolism in patients with muscular dystrophy in early or acute phase of disease: In vitro, high resolution NMR spectroscopy based analysis. Clin. Chim. Acta 2018, 478, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Cieslarova, Z.; Lopes, F.S.; do Lago, C.L.; Franca, M.C., Jr.; Colnaghi Simionato, A.V. Capillary electrophoresis tandem mass spectrometry determination of glutamic acid and homocysteine’s metabolites: Potential biomarkers of amyotrophic lateral sclerosis. Talanta 2017, 170, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Patin, F.; Corcia, P.; Vourc’h, P.; Nadal-Desbarats, L.; Baranek, T.; Goossens, J.F.; Marouillat, S.; Dessein, A.F.; Descat, A.; Madji Hounoum, B.; et al. Omics to Explore Amyotrophic Lateral Sclerosis Evolution: The Central Role of Arginine and Proline Metabolism. Mol. Neurobiol. 2017, 54, 5361–5374. [Google Scholar] [CrossRef]
- Files, D.C.; Ilaiwy, A.; Parry, T.L.; Gibbs, K.W.; Liu, C.; Bain, J.R.; Delbono, O.; Muehlbauer, M.J.; Willis, M.S. Lung injury-induced skeletal muscle wasting in aged mice is linked to alterations in long chain fatty acid metabolism. Metabolomics 2016, 12, 134. [Google Scholar] [CrossRef]
- Moaddel, R.; Fabbri, E.; Khadeer, M.A.; Carlson, O.D.; Gonzalez-Freire, M.; Zhang, P.; Semba, R.D.; Ferrucci, L. Plasma Biomarkers of Poor Muscle Quality in Older Men and Women from the Baltimore Longitudinal Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 1266–1272. [Google Scholar] [CrossRef] [Green Version]
- Wuolikainen, A.; Jonsson, P.; Ahnlund, M.; Antti, H.; Marklund, S.L.; Moritz, T.; Forsgren, L.; Andersen, P.M.; Trupp, M. Multi-platform mass spectrometry analysis of the CSF and plasma metabolomes of rigorously matched amyotrophic lateral sclerosis, Parkinson’s disease and control subjects. Mol. Biosyst. 2016, 12, 1287–1298. [Google Scholar] [CrossRef]
- Sengupta, M.; Cheema, A.; Kaminski, H.J.; Kusner, L.L.; Muscle Study, G. Serum metabolomic response of myasthenia gravis patients to chronic prednisone treatment. PLoS ONE 2014, 9, e102635. [Google Scholar] [CrossRef]
- Cirulli, E.T.; Guo, L.; Leon Swisher, C.; Shah, N.; Huang, L.; Napier, L.A.; Kirkness, E.F.; Spector, T.D.; Caskey, C.T.; Thorens, B.; et al. Profound Perturbation of the Metabolome in Obesity Is Associated with Health Risk. Cell Metab. 2018. [Google Scholar] [CrossRef]
- Libert, D.M.; Nowacki, A.S.; Natowicz, M.R. Metabolomic analysis of obesity, metabolic syndrome, and type 2 diabetes: Amino acid and acylcarnitine levels change along a spectrum of metabolic wellness. PeerJ 2018, 6, e5410. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.C.; Playdon, M.C.; Sampson, J.N.; Hoover, R.N.; Trabert, B.; Matthews, C.E.; Ziegler, R.G. A Metabolomics Analysis of Body Mass Index and Postmenopausal Breast Cancer Risk. J. Natl. Cancer Inst. 2018, 110, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Muniandy, M.; Velagapudi, V.; Hakkarainen, A.; Lundbom, J.; Lundbom, N.; Rissanen, A.; Kaprio, J.; Pietilainen, K.H.; Ollikainen, M. Plasma metabolites reveal distinct profiles associating with different metabolic risk factors in monozygotic twin pairs. Int. J. Obes. (Lond.) 2018. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Kim, M.; Kim, M.; Kang, M.; Yoo, H.J.; Lee, N.H.; Kim, Y.H.; Song, M.; Lee, J.H. Metabolites distinguishing visceral fat obesity and atherogenic traits in individuals with overweight. Obesity (Silver Spring) 2017, 25, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Carayol, M.; Leitzmann, M.F.; Ferrari, P.; Zamora-Ros, R.; Achaintre, D.; Stepien, M.; Schmidt, J.A.; Travis, R.C.; Overvad, K.; Tjonneland, A.; et al. Blood Metabolic Signatures of Body Mass Index: A Targeted Metabolomics Study in the EPIC Cohort. J. Proteome Res. 2017, 16, 3137–3146. [Google Scholar] [CrossRef] [PubMed]
- Okekunle, A.P.; Li, Y.; Liu, L.; Du, S.; Wu, X.; Chen, Y.; Li, Y.; Qi, J.; Sun, C.; Feng, R. Abnormal circulating amino acid profiles in multiple metabolic disorders. Diabetes Res. Clin. Pract. 2017, 132, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Bogl, L.H.; Kaye, S.M.; Ramo, J.T.; Kangas, A.J.; Soininen, P.; Hakkarainen, A.; Lundbom, J.; Lundbom, N.; Ortega-Alonso, A.; Rissanen, A.; et al. Abdominal obesity and circulating metabolites: A twin study approach. Metabolism 2016, 65, 111–121. [Google Scholar] [CrossRef]
- Dugas, L.R.; Chorell, E.; Plange-Rhule, J.; Lambert, E.V.; Cao, G.; Cooper, R.S.; Layden, B.T.; Scholten, D.; Olsson, T.; Luke, A.; et al. Obesity-related metabolite profiles of black women spanning the epidemiologic transition. Metabolomics 2016, 12, 45. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, W.; Wang, Y.; Pedram, P.; Cahill, F.; Zhai, G.; Randell, E.; Gulliver, W.; Sun, G. Serum metabolic biomarkers distinguish metabolically healthy peripherally obese from unhealthy centrally obese individuals. Nutr. Metab. (Lond.) 2016, 13, 33. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.E.; Larson, M.G.; Ghorbani, A.; Cheng, S.; Chen, M.H.; Keyes, M.; Rhee, E.P.; Clish, C.B.; Vasan, R.S.; Gerszten, R.E.; et al. Metabolomic Profiles of Body Mass Index in the Framingham Heart Study Reveal Distinct Cardiometabolic Phenotypes. PLoS ONE 2016, 11, e0148361. [Google Scholar] [CrossRef]
- Tulipani, S.; Palau-Rodriguez, M.; Minarro Alonso, A.; Cardona, F.; Marco-Ramell, A.; Zonja, B.; Lopez de Alda, M.; Munoz-Garach, A.; Sanchez-Pla, A.; Tinahones, F.J.; et al. Biomarkers of Morbid Obesity and Prediabetes by Metabolomic Profiling of Human Discordant Phenotypes. Clin. Chim. Acta 2016, 463, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shen, J.; Djukovic, D.; Daniel-MacDougall, C.; Gu, H.; Wu, X.; Chow, W.H. Metabolomics-identified metabolites associated with body mass index and prospective weight gain among Mexican American women. Obes. Sci. Pract. 2016, 2, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulet, M.M.; Chevrier, G.; Grenier-Larouche, T.; Pelletier, M.; Nadeau, M.; Scarpa, J.; Prehn, C.; Marette, A.; Adamski, J.; Tchernof, A. Alterations of plasma metabolite profiles related to adipose tissue distribution and cardiometabolic risk. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E736–E746. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Tseng, Y.J.; Wang, S.Y.; Tsai, Y.S.; Chang, C.S.; Kuo, T.C.; Yao, W.J.; Shieh, C.C.; Wu, C.H.; Kuo, P.H. The metabolome profiling and pathway analysis in metabolic healthy and abnormal obesity. Int. J. Obes. (Lond.) 2015, 39, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Gralka, E.; Luchinat, C.; Tenori, L.; Ernst, B.; Thurnheer, M.; Schultes, B. Metabolomic fingerprint of severe obesity is dynamically affected by bariatric surgery in a procedure-dependent manner. Am. J. Clin. Nutr. 2015, 102, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floegel, A.; Wientzek, A.; Bachlechner, U.; Jacobs, S.; Drogan, D.; Prehn, C.; Adamski, J.; Krumsiek, J.; Schulze, M.B.; Pischon, T.; et al. Linking diet, physical activity, cardiorespiratory fitness and obesity to serum metabolite networks: Findings from a population-based study. Int. J. Obes. (Lond.) 2014, 38, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.C.; Matthews, C.E.; Sampson, J.N.; Stolzenberg-Solomon, R.Z.; Zheng, W.; Cai, Q.; Tan, Y.T.; Chow, W.H.; Ji, B.T.; Liu, D.K.; et al. Human metabolic correlates of body mass index. Metabolomics 2014, 10, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.P.; Montoliu, I.; Collino, S.; Scherer, M.; Guy, P.; Tavazzi, I.; Thorimbert, A.; Moco, S.; Rothney, M.P.; Ergun, D.L.; et al. Topographical body fat distribution links to amino acid and lipid metabolism in healthy obese women [corrected]. PLoS ONE 2013, 8, e73445. [Google Scholar] [CrossRef]
- Batch, B.C.; Shah, S.H.; Newgard, C.B.; Turer, C.B.; Haynes, C.; Bain, J.R.; Muehlbauer, M.; Patel, M.J.; Stevens, R.D.; Appel, L.J.; et al. Branched chain amino acids are novel biomarkers for discrimination of metabolic wellness. Metabolism 2013, 62, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Ali-Sisto, T.; Tolmunen, T.; Viinamaki, H.; Mantyselka, P.; Valkonen-Korhonen, M.; Koivumaa-Honkanen, H.; Honkalampi, K.; Ruusunen, A.; Nandania, J.; Velagapudi, V.; et al. Global arginine bioavailability ratio is decreased in patients with major depressive disorder. J. Affect. Disord. 2018, 229, 145–151. [Google Scholar] [CrossRef]
- Kawamura, N.; Shinoda, K.; Sato, H.; Sasaki, K.; Suzuki, M.; Yamaki, K.; Fujimori, T.; Yamamoto, H.; Osei-Hyiaman, D.; Ohashi, Y. Plasma metabolome analysis of patients with major depressive disorder. Psychiatry Clin. Neurosci. 2018, 72, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Moaddel, R.; Shardell, M.; Khadeer, M.; Lovett, J.; Kadriu, B.; Ravichandran, S.; Morris, P.J.; Yuan, P.; Thomas, C.J.; Gould, T.D.; et al. Plasma metabolomic profiling of a ketamine and placebo crossover trial of major depressive disorder and healthy control subjects. Psychopharmacology (Berl.) 2018, 235, 3017–3030. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zheng, P.; Zhao, L.; Jia, J.; Tang, S.; Xu, P.; Xie, P.; Gao, H. Predictive diagnosis of major depression using NMR-based metabolomics and least-squares support vector machine. Clin. Chim. Acta 2017, 464, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Ali-Sisto, T.; Tolmunen, T.; Toffol, E.; Viinamaki, H.; Mantyselka, P.; Valkonen-Korhonen, M.; Honkalampi, K.; Ruusunen, A.; Velagapudi, V.; Lehto, S.M. Purine metabolism is dysregulated in patients with major depressive disorder. Psychoneuroendocrinology 2016, 70, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yieh, L.; Yang, T.; Drinkenburg, W.; Peeters, P.; Steckler, T.; Narayan, V.A.; Wittenberg, G.; Ye, J. Metabolomic biosignature differentiates melancholic depressive patients from healthy controls. BMC Genom. 2016, 17, 669. [Google Scholar] [CrossRef] [PubMed]
- Rotroff, D.M.; Corum, D.G.; Motsinger-Reif, A.; Fiehn, O.; Bottrel, N.; Drevets, W.C.; Singh, J.; Salvadore, G.; Kaddurah-Daouk, R. Metabolomic signatures of drug response phenotypes for ketamine and esketamine in subjects with refractory major depressive disorder: New mechanistic insights for rapid acting antidepressants. Transl. Psychiatry 2016, 6, e894. [Google Scholar] [CrossRef] [PubMed]
- Setoyama, D.; Kato, T.A.; Hashimoto, R.; Kunugi, H.; Hattori, K.; Hayakawa, K.; Sato-Kasai, M.; Shimokawa, N.; Kaneko, S.; Yoshida, S.; et al. Plasma Metabolites Predict Severity of Depression and Suicidal Ideation in Psychiatric Patients-A Multicenter Pilot Analysis. PLoS ONE 2016, 11, e0165267. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Chen, J.J.; Zhou, C.J.; Zeng, L.; Li, K.W.; Sun, L.; Liu, M.L.; Zhu, D.; Liang, Z.H.; Xie, P. Identification of sex-specific urinary biomarkers for major depressive disorder by combined application of NMR- and GC-MS-based metabonomics. Transl. Psychiatry 2016, 6, e955. [Google Scholar] [CrossRef]
- Woo, H.I.; Chun, M.R.; Yang, J.S.; Lim, S.W.; Kim, M.J.; Kim, S.W.; Myung, W.J.; Kim, D.K.; Lee, S.Y. Plasma amino acid profiling in major depressive disorder treated with selective serotonin reuptake inhibitors. CNS Neurosci. Ther. 2015, 21, 417–424. [Google Scholar] [CrossRef]
- Zheng, P.; Gao, H.C.; Li, Q.; Shao, W.H.; Zhang, M.L.; Cheng, K.; Yang, D.Y.; Fan, S.H.; Chen, L.; Fang, L.; et al. Plasma metabonomics as a novel diagnostic approach for major depressive disorder. J. Proteome Res. 2012, 11, 1741–1748. [Google Scholar] [CrossRef]
- Paige, L.A.; Mitchell, M.W.; Krishnan, K.R.; Kaddurah-Daouk, R.; Steffens, D.C. A preliminary metabolomic analysis of older adults with and without depression. Int. J. Geriatr. Psychiatry 2007, 22, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Dickenson, A.H.; Baron, R. Osteoarthritis pain: Nociceptive or neuropathic? Nat. Rev. Rheumatol. 2014, 10, 374–380. [Google Scholar] [CrossRef]
- La Porta, C.; Bura, S.A.; Negrete, R.; Maldonado, R. Involvement of the endocannabinoid system in osteoarthritis pain. Eur. J. Neurosci. 2014, 39, 485–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the cannabinoid receptor system in synovial tissue and fluid in patients with osteoarthritis and rheumatoid arthritis. Arthritis Res. Ther. 2008, 10, R43. [Google Scholar] [CrossRef] [PubMed]
- Muccioli, G.G. Endocannabinoid biosynthesis and inactivation, from simple to complex. Drug Discov. Today 2010, 15, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Oiestad, B.E.; Juhl, C.B.; Eitzen, I.; Thorlund, J.B. Knee extensor muscle weakness is a risk factor for development of knee osteoarthritis. A systematic review and meta-analysis. Osteoarthritis Cartilage 2015, 23, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Culvenor, A.G.; Ruhdorfer, A.; Juhl, C.; Eckstein, F.; Oiestad, B.E. Knee Extensor Strength and Risk of Structural, Symptomatic, and Functional Decline in Knee Osteoarthritis: A Systematic Review and Meta-Analysis. Arthrit. Care Res. 2017, 69, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ghosh, D.; Singh, R.L. Amyotrophic Lateral Sclerosis and Metabolomics: Clinical Implication and Therapeutic Approach. J. Biomark. 2013, 2013, 538765. [Google Scholar] [CrossRef] [PubMed]
- Blasco, H.; Patin, F.; Madji Hounoum, B.; Gordon, P.H.; Vourc’h, P.; Andres, C.R.; Corcia, P. Metabolomics in amyotrophic lateral sclerosis: How far can it take us? Eur. J. Neurol. 2016, 23, 447–454. [Google Scholar] [CrossRef]
- Edwards, R.R.; Cahalan, C.; Mensing, G.; Smith, M.; Haythornthwaite, J.A. Pain, catastrophizing, and depression in the rheumatic diseases. Nat. Rev. Rheumatol. 2011, 7, 216–224. [Google Scholar] [CrossRef]
- Miyamoto, K.; Ohkawara, B.; Ito, M.; Masuda, A.; Hirakawa, A.; Sakai, T.; Hiraiwa, H.; Hamada, T.; Ishiguro, N.; Ohno, K. Fluoxetine ameliorates cartilage degradation in osteoarthritis by inhibiting Wnt/beta-catenin signaling. PLoS ONE 2017, 12, e0184388. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Nakatani, Y.; Atsumi, G.I.; Inoue, K.; Kudo, I. Regulatory Functions of Phospholipase A2. Crit. Rev. Immunol. 2017, 37, 121–179. [Google Scholar] [CrossRef] [PubMed]
- Jamal, O.S.; Conaghan, P.G.; Cunningham, A.M.; Brooks, P.M.; Munro, V.F.; Scott, K.F. Increased expression of human type IIa secretory phospholipase A2 antigen in arthritic synovium. Ann. Rheum. Dis. 1998, 57, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruzanski, W.; Bogoch, E.; Katz, A.; Wloch, M.; Stefanski, E.; Grouix, B.; Sakotic, G.; Vadas, P. Induction of release of secretory nonpancreatic phospholipase A2 from human articular chondrocytes. J. Rheumatol. 1995, 22, 2114–2119. [Google Scholar] [PubMed]
- Tokumura, A.; Harada, K.; Fukuzawa, K.; Tsukatani, H. Involvement of lysophospholipase D in the production of lysophosphatidic acid in rat plasma. Biochim. Biophys. Acta 1986, 875, 31–38. [Google Scholar] [PubMed]
- Tokumura, A.; Majima, E.; Kariya, Y.; Tominaga, K.; Kogure, K.; Yasuda, K.; Fukuzawa, K. Identification of human plasma lysophospholipase D, a lysophosphatidic acid-producing enzyme, as autotaxin, a multifunctional phosphodiesterase. J. Biol. Chem. 2002, 277, 39436–39442. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Rashid, M.H.; Fujita, R.; Contos, J.J.; Chun, J.; Ueda, H. Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med. 2004, 10, 712–718. [Google Scholar] [CrossRef]
- Uchida, H.; Nagai, J.; Ueda, H. Lysophosphatidic acid and its receptors LPA1 and LPA3 mediate paclitaxel-induced neuropathic pain in mice. Mol. Pain 2014, 10, 71. [Google Scholar] [CrossRef]
- Inoue, M.; Xie, W.; Matsushita, Y.; Chun, J.; Aoki, J.; Ueda, H. Lysophosphatidylcholine induces neuropathic pain through an action of autotaxin to generate lysophosphatidic acid. Neuroscience 2008, 152, 296–298. [Google Scholar] [CrossRef]
- Gustin, C.; Van Steenbrugge, M.; Raes, M. LPA modulates monocyte migration directly and via LPA-stimulated endothelial cells. Am. J. Physiol. Cell Physiol. 2008, 295, C905–C914. [Google Scholar] [CrossRef]
- Zhou, D.; Luini, W.; Bernasconi, S.; Diomede, L.; Salmona, M.; Mantovani, A.; Sozzani, S. Phosphatidic acid and lysophosphatidic acid induce haptotactic migration of human monocytes. J. Biol. Chem. 1995, 270, 25549–25556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wei, J.; Weathington, N.; Jacko, A.M.; Huang, H.; Tsung, A.; Zhao, Y. Lysophosphatidic acid receptor 1 antagonist ki16425 blunts abdominal and systemic inflammation in a mouse model of peritoneal sepsis. Transl. Res. 2015, 166, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldshmit, Y.; Matteo, R.; Sztal, T.; Ellett, F.; Frisca, F.; Moreno, K.; Crombie, D.; Lieschke, G.J.; Currie, P.D.; Sabbadini, R.A.; et al. Blockage of lysophosphatidic acid signaling improves spinal cord injury outcomes. Am. J. Pathol. 2012, 181, 978–992. [Google Scholar] [CrossRef] [PubMed]
- Mabey, T.; Taleongpong, P.; Udomsinprasert, W.; Jirathanathornnukul, N.; Honsawek, S. Plasma and synovial fluid autotaxin correlate with severity in knee osteoarthritis. Clin. Chim. Acta 2015, 444, 72–77. [Google Scholar] [CrossRef] [PubMed]
- McDougall, J.J.; Albacete, S.; Schuelert, N.; Mitchell, P.G.; Lin, C.; Oskins, J.L.; Bui, H.H.; Chambers, M.G. Lysophosphatidic acid provides a missing link between osteoarthritis and joint neuropathic pain. Osteoarthritis Cartilage 2017, 25, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Thirunavukkarasu, K.; Swearingen, C.A.; Oskins, J.L.; Lin, C.; Bui, H.H.; Jones, S.B.; Pfeifer, L.A.; Norman, B.H.; Mitchell, P.G.; Chambers, M.G. Identification and pharmacological characterization of a novel inhibitor of autotaxin in rodent models of joint pain. Osteoarthritis Cartilage 2017, 25, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Vuolteenaho, K.; Koskinen, A.; Moilanen, T.; Moilanen, E. Leptin levels are increased and its negative regulators, SOCS-3 and sOb-R are decreased in obese patients with osteoarthritis: A link between obesity and osteoarthritis. Ann. Rheum. Dis. 2012, 71, 1912–1913. [Google Scholar] [CrossRef]
- Stannus, O.P.; Cao, Y.; Antony, B.; Blizzard, L.; Cicuttini, F.; Jones, G.; Ding, C. Cross-sectional and longitudinal associations between circulating leptin and knee cartilage thickness in older adults. Ann. Rheum. Dis. 2015, 74, 82–88. [Google Scholar] [CrossRef]
- Zhang, P.; Zhong, Z.H.; Yu, H.T.; Liu, B. Significance of increased leptin expression in osteoarthritis patients. PLoS ONE 2015, 10, e0123224. [Google Scholar] [CrossRef]
- Beekhuizen, M.; Gierman, L.M.; van Spil, W.E.; Van Osch, G.J.; Huizinga, T.W.; Saris, D.B.; Creemers, L.B.; Zuurmond, A.M. An explorative study comparing levels of soluble mediators in control and osteoarthritic synovial fluid. Osteoarthritis Cartilage 2013, 21, 918–922. [Google Scholar] [CrossRef] [Green Version]
- Rockel, J.S.; Kapoor, M. Autophagy: Controlling cell fate in rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 193. [Google Scholar] [CrossRef] [PubMed]
- Neishabouri, S.H.; Hutson, S.M.; Davoodi, J. Chronic activation of mTOR complex 1 by branched chain amino acids and organ hypertrophy. Amino Acids 2015, 47, 1167–1182. [Google Scholar] [CrossRef] [PubMed]
- Zhenyukh, O.; Civantos, E.; Ruiz-Ortega, M.; Sanchez, M.S.; Vazquez, C.; Peiro, C.; Egido, J.; Mas, S. High concentration of branched-chain amino acids promotes oxidative stress, inflammation and migration of human peripheral blood mononuclear cells via mTORC1 activation. Free Radic. Biol. Med. 2017, 104, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vasheghani, F.; Li, Y.H.; Blati, M.; Simeone, K.; Fahmi, H.; Lussier, B.; Roughley, P.; Lagares, D.; Pelletier, J.P.; et al. Cartilage-specific deletion of mTOR upregulates autophagy and protects mice from osteoarthritis. Ann. Rheum. Dis. 2015, 74, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Kawakami, Y.; Kobayashi, M.; Greco, N.; Cummins, J.H.; Matsushita, T.; Kuroda, R.; Kurosaka, M.; Fu, F.H.; Huard, J. Local intra-articular injection of rapamycin delays articular cartilage degeneration in a murine model of osteoarthritis. Arthritis Res. Ther. 2014, 16, 482. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef]
- Abramson, S.B.; Amin, A.R.; Clancy, R.M.; Attur, M. The role of nitric oxide in tissue destruction. Best Pract. Res. Clin. Rheumatol. 2001, 15, 831–845. [Google Scholar] [CrossRef]
- Shearer, J.D.; Richards, J.R.; Mills, C.D.; Caldwell, M.D. Differential regulation of macrophage arginine metabolism: A proposed role in wound healing. Am. J. Physiol. 1997, 272, E181–E190. [Google Scholar] [CrossRef]
- Wehling-Henricks, M.; Jordan, M.C.; Gotoh, T.; Grody, W.W.; Roos, K.P.; Tidball, J.G. Arginine metabolism by macrophages promotes cardiac and muscle fibrosis in mdx muscular dystrophy. PLoS ONE 2010, 5, e10763. [Google Scholar] [CrossRef]
- Moore, J.P.; Vinh, A.; Tuck, K.L.; Sakkal, S.; Krishnan, S.M.; Chan, C.T.; Lieu, M.; Samuel, C.S.; Diep, H.; Kemp-Harper, B.K.; et al. M2 macrophage accumulation in the aortic wall during angiotensin II infusion in mice is associated with fibrosis, elastin loss, and elevated blood pressure. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H906–H917. [Google Scholar] [CrossRef]
- Wang, Y.; Wehling-Henricks, M.; Samengo, G.; Tidball, J.G. Increases of M2a macrophages and fibrosis in aging muscle are influenced by bone marrow aging and negatively regulated by muscle-derived nitric oxide. Aging Cell 2015, 14, 678–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce, J.T.; Ramalingam, T.R.; Mentink-Kane, M.M.; Wilson, M.S.; El Kasmi, K.C.; Smith, A.M.; Thompson, R.W.; Cheever, A.W.; Murray, P.J.; Wynn, T.A. Arginase-1-expressing macrophages suppress Th2 cytokine-driven inflammation and fibrosis. PLoS Pathog. 2009, 5, e1000371. [Google Scholar] [CrossRef] [PubMed]
- Barron, L.; Smith, A.M.; El Kasmi, K.C.; Qualls, J.E.; Huang, X.; Cheever, A.; Borthwick, L.A.; Wilson, M.S.; Murray, P.J.; Wynn, T.A. Role of arginase 1 from myeloid cells in th2-dominated lung inflammation. PLoS ONE 2013, 8, e61961. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Nakashima, Y.; Okazaki, K.; Mawatari, T.; Fukushi, J.; Oyamada, A.; Fujimura, K.; Iwamoto, Y.; Yoshikai, Y. Preferential accumulation of activated Th1 cells not only in rheumatoid arthritis but also in osteoarthritis joints. J. Rheumatol. 2011, 38, 1569–1575. [Google Scholar] [CrossRef] [PubMed]
- Dolganiuc, A.; Stavaru, C.; Anghel, M.; Georgescu, E.; Chichos, B.; Olinescu, A. Shift toward T lymphocytes with Th1 and Tc1 cytokine-secterion profile in the joints of patients with osteoarthritis. Roum. Arch. Microbiol. Immunol. 1999, 58, 249–258. [Google Scholar] [PubMed]
- Ishii, H.; Tanaka, H.; Katoh, K.; Nakamura, H.; Nagashima, M.; Yoshino, S. Characterization of infiltrating T cells and Th1/Th2-type cytokines in the synovium of patients with osteoarthritis. Osteoarthritis Cartilage 2002, 10, 277–281. [Google Scholar] [CrossRef]
- Vuolteenaho, K.; Moilanen, T.; Knowles, R.G.; Moilanen, E. The role of nitric oxide in osteoarthritis. Scand. J. Rheumatol. 2007, 36, 247–258. [Google Scholar] [CrossRef]
- Clements, K.M.; Price, J.S.; Chambers, M.G.; Visco, D.M.; Poole, A.R.; Mason, R.M. Gene deletion of either interleukin-1beta, interleukin-1beta-converting enzyme, inducible nitric oxide synthase, or stromelysin 1 accelerates the development of knee osteoarthritis in mice after surgical transection of the medial collateral ligament and partial medial meniscectomy. Arthritis Rheum. 2003, 48, 3452–3463. [Google Scholar]


{kind=link}
| Author | Year | Fluid/Tissue for Metabolite Detection | Species | Study Groups | Metabolite Detection Method | Reference |
|---|---|---|---|---|---|---|
| Anderson et al. | 2018 | synovial fluid | equine | septic vs. non-septic joint pathologies | 1H-NMR | [14] |
| Carlson et al. | 2018 | synovial fluid | human | OA vs. RA vs. healthy | LC-MS | [15] |
| Hinata et al. | 2018 | synovial fluid | rat | control vs. MIA-induced OA, sham vs. meniscectomy-induced OA | LC-MS/MS | [16] |
| human | OA only | |||||
| Zhang et al. | 2016 | plasma | human | primary OA at TKR vs. healthy control | LC-MS/MS | [17] |
| Jin et al. | 2016 | synovial fluid | human | degenerative vs. traumatic vs. infectious vs. inflammatory OA | In vivo 1H-MRS | [18] |
| Loeser et al. | 2016 | urine | human | OA progression vs. stable | 1H-NMR | [19] |
| Mickiewicz et al. | 2016 | serum | mouse | sham vs. DMM; wild type vs. Integrin 1α-null; erlotinib vs. vehicle | 1H-NMR | [20] |
| Hu et al. | 2016 | plasma | human | primary OA at TKR vs. healthy control | LC-MS/MS | [21] |
| Zhang et al. | 2016 | plasma | human | primary OA at TKR vs. healthy control | LC-MS/MS | [22] |
| Tufts et al. | 2015 | knee articular cartilage | human | primary OA at TKR | HRMAS-NMR | [23] |
| Zhang et al. | 2015 | plasma, synovial fluid | human | primary OA at TKR | LC-MS/MS | [24] |
| Zhai et al. | 2010 | serum | human | OA vs. healthy control | LC-MS/MS | [25] |
| Davies et al. | 2009 | synovial fluid, serum, cartilage | human | active OA, inactive OA, post-mortem controls | HPLC | [26] |
| Lamers et al. | 2005 | urine | human | radiographic OA vs. non-OA controls | 1H-NMR | [27] |
| Basu et al. | 2001 | serum, synovial fluid | human | control (serum only) vs. OA vs. RA vs. ReA vs. PsA | radioimmunoassay | [28] |
| Phenotype | Author | Year | Fluid/Tissue for Metabolite Detection | Species | Study Groups | Metabolite Detection Method | Reference |
|---|---|---|---|---|---|---|---|
| Pain | Finco et al. | 2016 | urine | human | nociceptive pain vs. neuropathic pain vs. pain free | 1H-NMR | [60] |
| Hadrevi et al. | 2015 | serum | human | women with chronic neck pain, chronic widespread pain vs. healthy control | GS-MS | [61] | |
| Um et al. | 2009 | urine | rat | celecoxib vs. indomethacin vs. ibuprofen vs. vehicle; gastric damaged vs. undamaged | 1H-NMR | [62] | |
| Muscle Strength | Srivastava et al. | 2018 | skeletal muscle | human | Duchenne muscular dystrophy vs. Becker muscular dystrophy vs. facioscapulohumeral dystrophy vs. limb girdle muscular dystrophy vs. healthy control | 1H-NMR | [63] |
| Cieslarova et al. | 2017 | plasma | human | ALS vs. healthy control | CE-MS/MS | [64] | |
| Patin et al. | 2017 | Muscle and brain (mouse only), plasma | human and mouse | mSOD1*G39A-transgenic mice vs. WT mice; ALS vs. healthy control | 1H-NMR | [65] | |
| Files et al. | 2016 | skeletal muscle | mouse | adult vs. old; sham vs. acute lung injury-induced muscle wasting | GS-MS | [66] | |
| Moaddel et al. | 2016 | plasma | human | low vs. high muscle quality in older men and women | LC-MS/MS | [67] | |
| Wuolikainen et al. | 2016 | CSF and Plasma | human | ALS and Parkinson’s disease vs. healthy control | GC-MS; LC-MS | [68] | |
| Sengupta et al. | 2014 | serum | human | myasthenia gravis prednisone treated vs. baseline | UPLC-ESI-QTOF-MS | [69] | |
| Obesity | Cirulli et al. | 2018 | serum, plasma | human | metabolically obese vs. metabolically overweight vs. metabolically healthy | LC-MS/MS | [70] |
| Libert et al. | 2018 | plasma | human | lean metabolically well vs. obese metabolically well vs. obese metabolically unwell vs. obese metabolically unwell with type II diabetes | LC-MS/MS | [71] | |
| Moore et al. | 2018 | serum | human | correlation of BMI and breast cancer risk to circulating metabolites in postmenopausal women | LC-MS/MS | [72] | |
| Munlandy et al. | 2018 | plasma | human | correlation of metabolites to cardiometabolic risk factors (including BMI, % body fat, visceral fat, subcutaneous fat) in monozygotic twins | LC-MS/MS | [73] | |
| Baek et al. | 2017 | plasma | human | low vs. high visceral fat area in a Korean cohort | LC-MS | [74] | |
| Carayol et al. | 2017 | serum, plasma | human | correlation of BMI to circulating metabolites | LC-MS/MS | [75] | |
| Okekunle et al. | 2017 | serum | human | obese vs. type II diabetes vs. metabolic syndrome vs. healthy control | UPLC-TQ/MS | [76] | |
| Zhong et al. | 2017 | plasma | human | obese vs. metabolic syndrome | LC-MS/MS | [57] | |
| Bogl et al. | 2016 | serum | human | correlation of phenotypic and obesity-related measures to metabolite levels in dizygotic and monozygotic twins | 1H-NMR | [77] | |
| Dugas et al. | 2016 | serum | human | normal vs. obese; black women from U.S. vs. South Africa vs. Ghana | GC-TOF/MS | [78] | |
| Gao et al. | 2016 | serum | human | metabolically unhealthy centrally obese vs. metabolically healthy peripherally obese | LC-MS/MS | [79] | |
| Ho et al. | 2016 | plasma | human | correlation of BMI, waist circumference, and other metabolic traits to circulating metabolites | LC-MS/MS | [80] | |
| Tulipani et al. | 2016 | serum | human | BMI-discordant non-diabetic vs. pre-diabetic monozygotic twins | LC-MS/MS; FIA-MS/MS; ESI-MS/MS | [81] | |
| Zhao et al. | 2016 | plasma | human | correlation of metabolites to BMI and weight gain in Mexican American women | LC-MS/MS | [82] | |
| Boulet et al. | 2015 | plasma | human | lean vs. overweight vs. obese women | ESI-LC-MS/MS, ESI-MS/MS | [83] | |
| Chen et al. | 2015 | serum | human | metabolic healthy obese vs. metabolic unhealthy obese | LC-MS; GC-MS | [84] | |
| Gralka et al. | 2015 | serum | human | obese vs. normal weight | 1H-NMR | [85] | |
| Floegel et al. | 2014 | serum | human | correlation of metabolite networks to different dietary, activity and anthropometric exposures (including BMI and waist circumference) | LC-MS/MS | [86] | |
| Moore et al. | 2014 | serum, plasma | human | correlation of metabolite levels to BMI | LC-MS/MS; GC-MS/MS | [87] | |
| Martin et al. | 2013 | plasma, urine | human | correlation of metabolites to body fat distribution in obese women | LC-MS/MS | [88] | |
| Batch et al. | 2013 | plasma | human | lean vs. overweight vs. obese | LC-MS/MS; ESI-MS/MS | [89] | |
| Depression | Ali-Sisto et al. | 2018 | serum | human | major depressive disorder vs. non-depressed controls, remitted vs. non-remitted patients with major depressive disorder | LC-MS | [90] |
| Kawamura et al. | 2018 | plasma | human | major depressive disorder vs. mentally healthy controls | CE-TOF/MS | [91] | |
| Moaddel et al. | 2018 | plasma | human | major depressive disorder vs. healthy controls, ketamine vs. placebo | LC-MS/MS | [92] | |
| Zheng et al. | 2017 | plasma | human | major depressive disorder vs. healthy controls | 1H-NMR | [93] | |
| Ali-Sisto et al. | 2016 | serum | human | major depressive disorder vs. non-depressed controls | LC-MS/MS | [94] | |
| Liu et al. | 2016 | plasma | human | healthy controls vs. major depressive disorder, melancholic depressed, anxious depressed | LC-MS/MS, GC-MS | [95] | |
| Rotroff et al. | 2016 | plasma | human | baseline vs. post-treatment of patients with major depressive disorder treated with placebo, ketamine, or esketamine | LC-MS/MS, GC-TOF/MS | [96] | |
| Setoyama et al. | 2016 | plasma | human | correlation of metabolites to depression severity in patients with psychiatric disorders, drug-free major depressive disorder, or bipolar disorders; medicated major depressive disorder and bipolar disorders | LC-MS | [97] | |
| Zheng et al. | 2016 | urine | human | major depressive disorder vs. healthy controls, women vs. men | 1H-NMR, GC-MS | [98] | |
| Woo et al. | 2015 | plasma | human | healthy controls vs. major depressive disorder patients baseline vs. major depressive disorder patients 6-weeks post SSRI treatment | LC-MS/MS | [99] | |
| Zheng et al. | 2012 | plasma | human | drug-naïve first episode depression vs. healthy controls | 1H-NMR | [100] | |
| Paige et al. | 2007 | plasma | human | remitted depressed vs. non-remitted depressed vs. non-depressed older adults | GC-MS | [101] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rockel, J.S.; Kapoor, M. The Metabolome and Osteoarthritis: Possible Contributions to Symptoms and Pathology. Metabolites 2018, 8, 92. https://doi.org/10.3390/metabo8040092
Rockel JS, Kapoor M. The Metabolome and Osteoarthritis: Possible Contributions to Symptoms and Pathology. Metabolites. 2018; 8(4):92. https://doi.org/10.3390/metabo8040092
Chicago/Turabian StyleRockel, Jason S., and Mohit Kapoor. 2018. "The Metabolome and Osteoarthritis: Possible Contributions to Symptoms and Pathology" Metabolites 8, no. 4: 92. https://doi.org/10.3390/metabo8040092
APA StyleRockel, J. S., & Kapoor, M. (2018). The Metabolome and Osteoarthritis: Possible Contributions to Symptoms and Pathology. Metabolites, 8(4), 92. https://doi.org/10.3390/metabo8040092