Metabolomic Dynamics Reveals Oxidative Stress in Spongy Tissue Disorder During Ripening of Mangifera indica L. Fruit


Abstract
:1. Introduction
2. Results
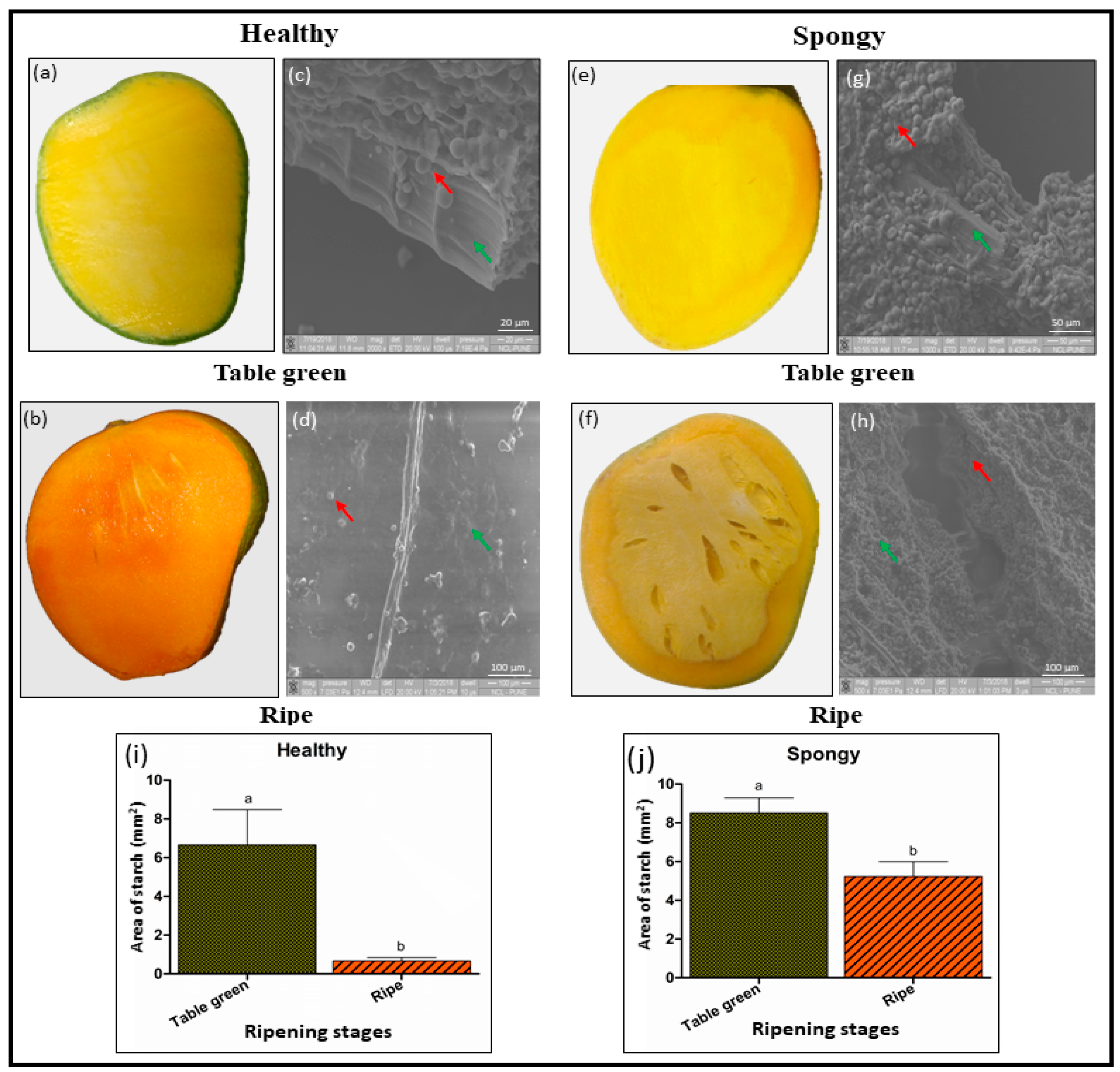
2.1. Accumulation of Starch Granules and Intact Parenchyma Cells Characterize the Spongy Mesocarp in Mango
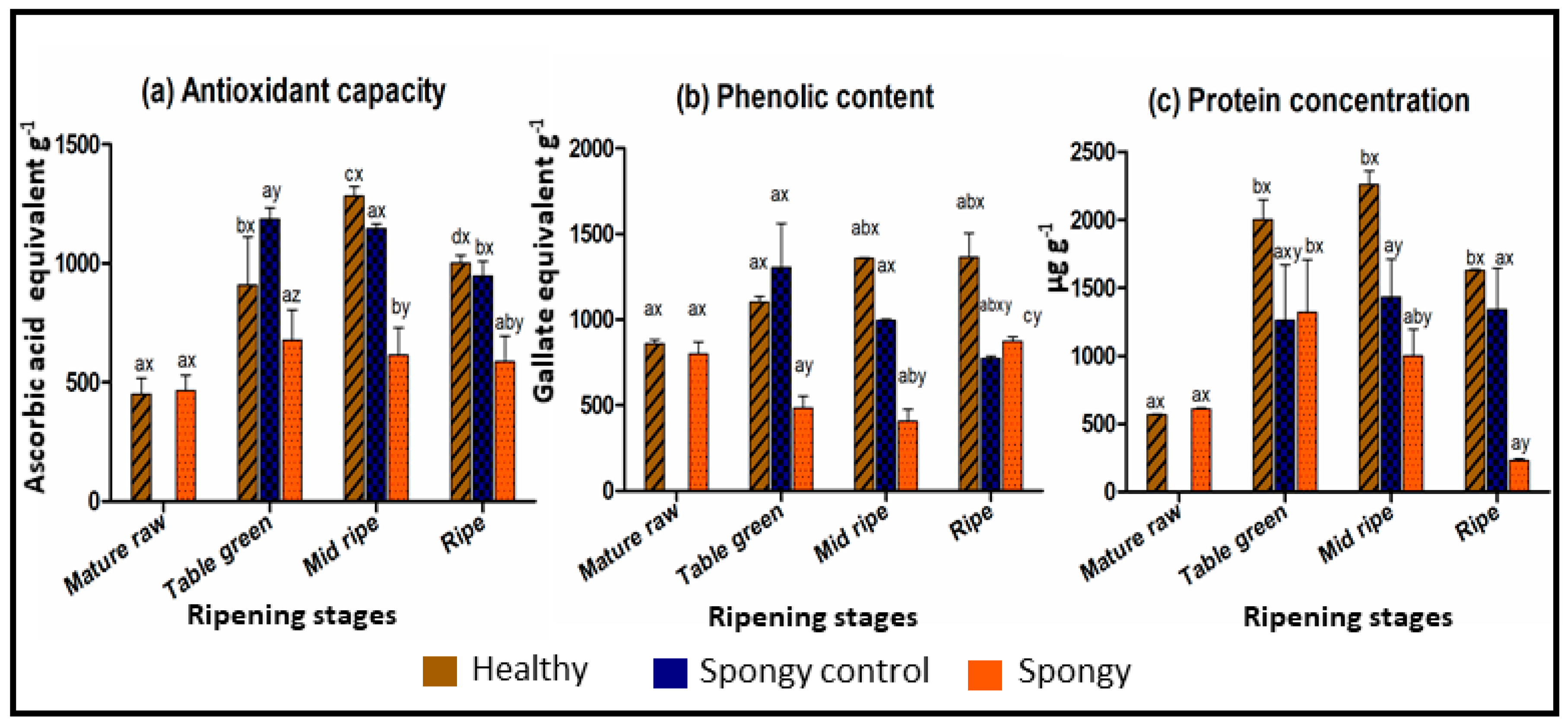
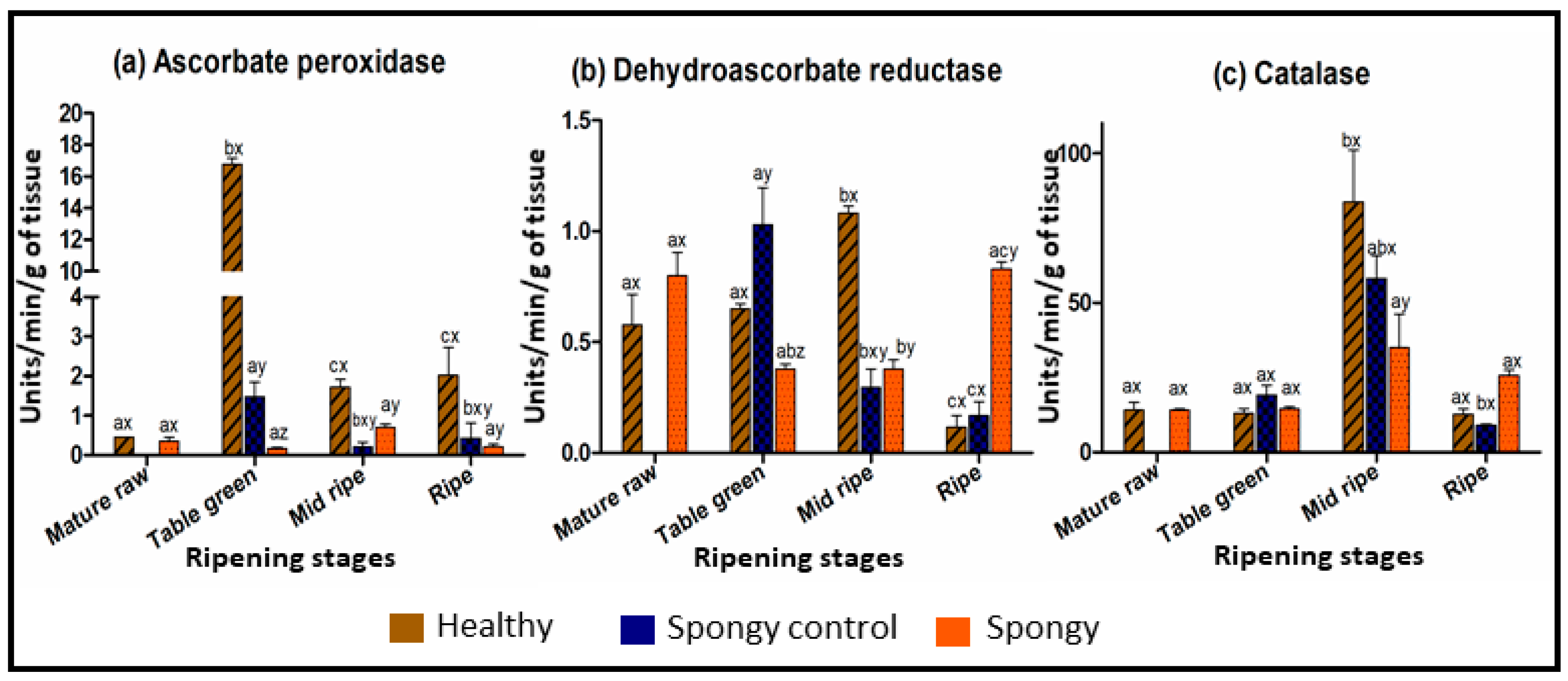
2.2. Altered Biochemical Composition of Fruit Mesocarp during Spongy Tissue Development
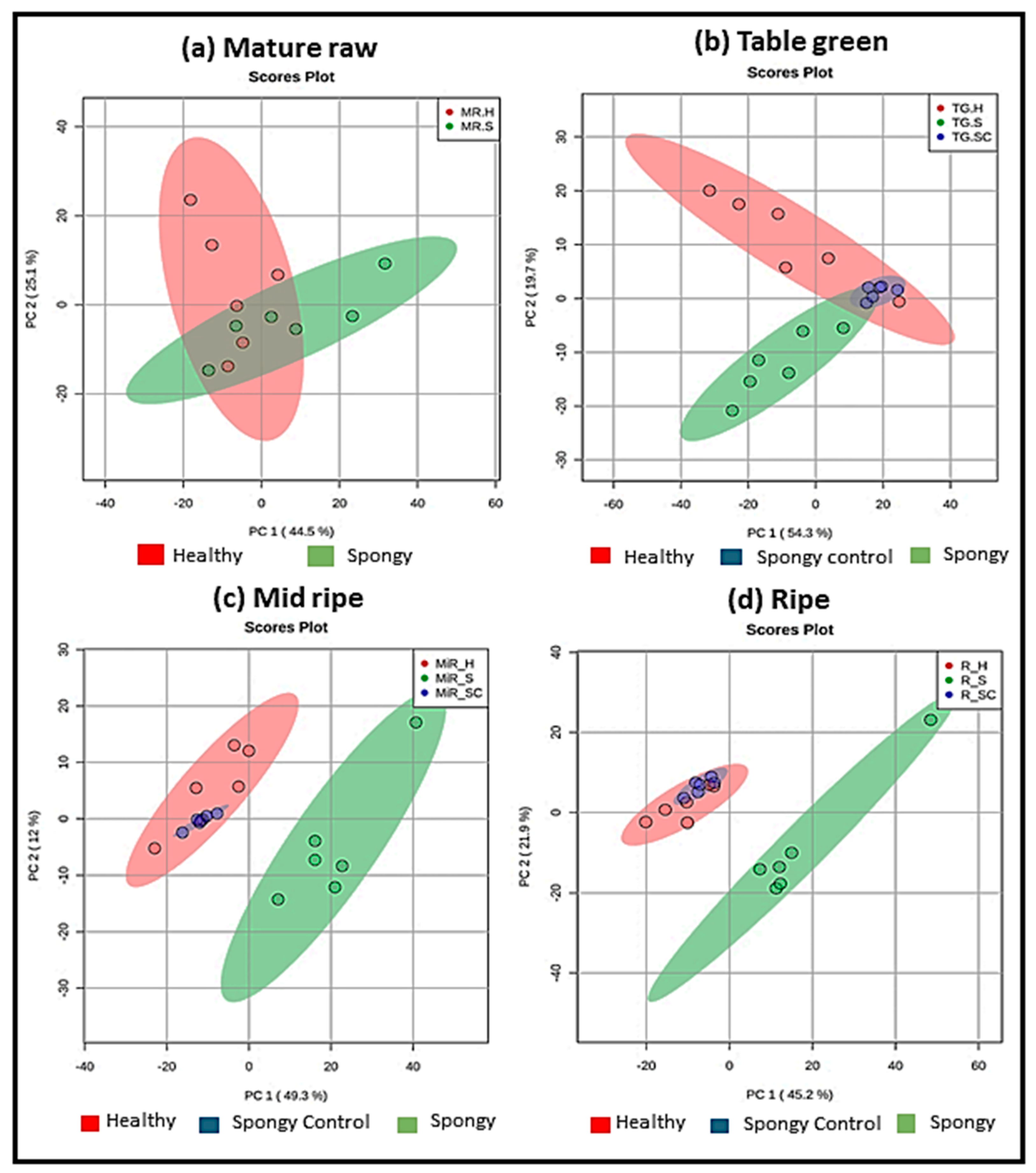
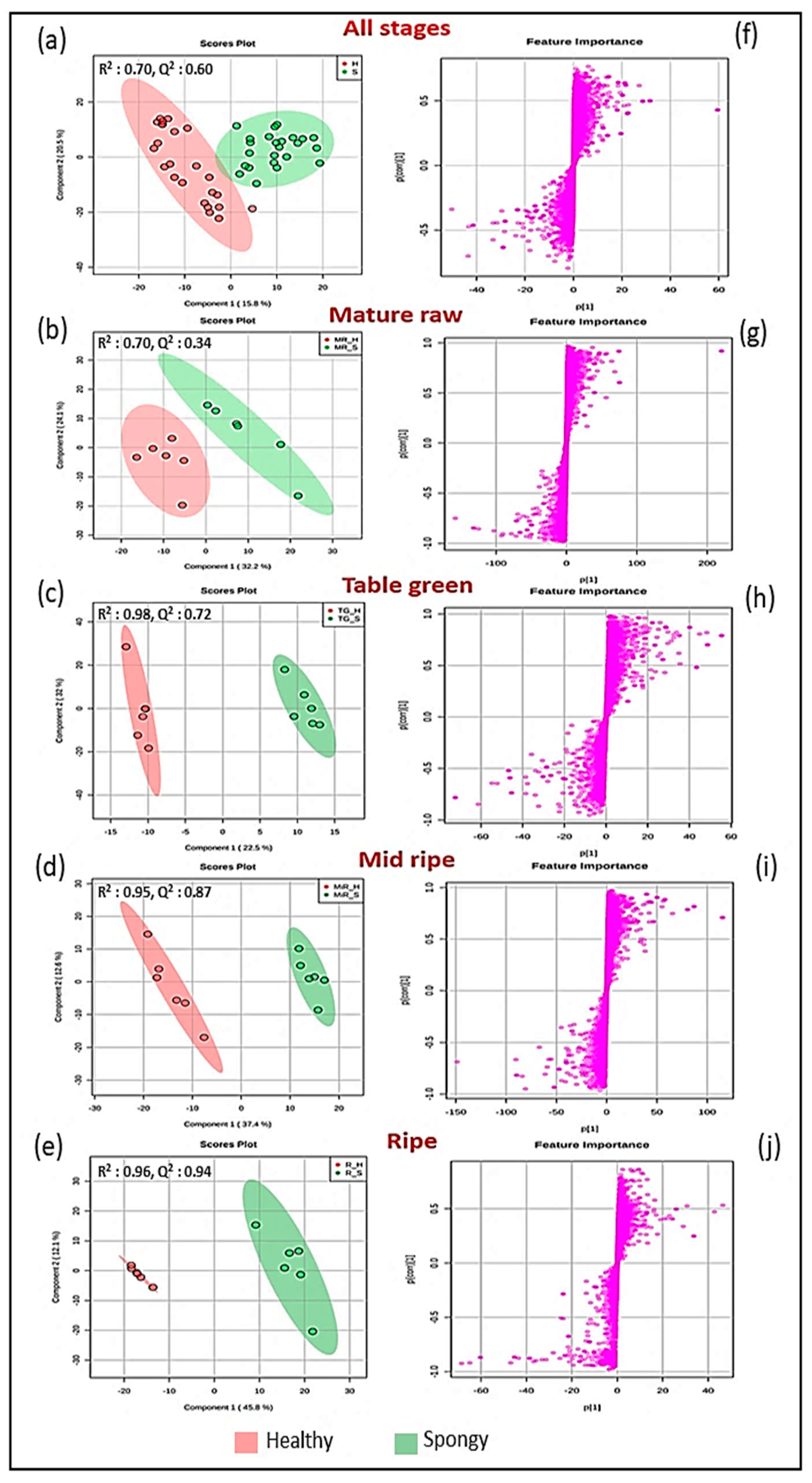
2.3. Plethora of Metabolites Involved in Spongy Tissue Formation
2.4. Metabolic Alterations in Spongy Tissue Formation
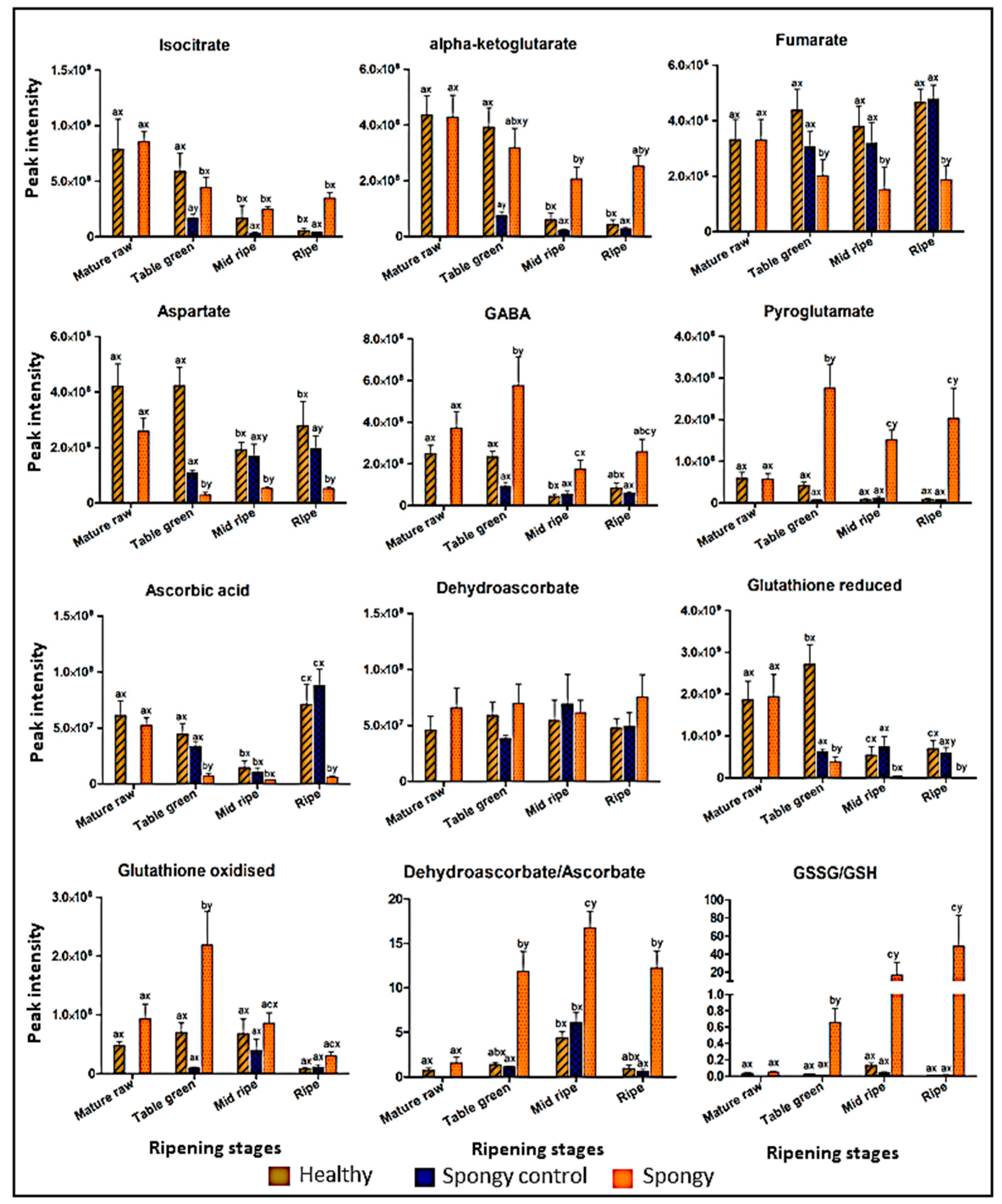
2.5. Profiling of Metabolites Involved in Primary Metabolism
2.6. Variation in the Flavor Metabolites in Spongy Tissue
3. Discussion
3.1. Cellular Changes in Spongy Mesocarp Hamper the Internal Physiology of Fruit
3.2. Deviation in Primry Metabolism in Spongy Tissue Disorder Highlights the Stress Conditions in the Fruit
3.3. Abiotic Stress Induces Cellular Oxidative Damage Leading to Spongy Tissue Formation in Alphonso
3.4. Modulation in Secondary Metabolites Affects the Flavor of Fruit in the Spongy Tissue Disorder
4. Materials and Methods
4.1. Plant Material
4.2. Scanning Electron Microscopy
4.3. Biochemical Analysis of Spongy Mesocarp Mango Tissues
4.4. Enzyme Assays
4.5. Metabolite Extraction and UHPLC-Q-Exactive-Orbitrap Mass Spectrometer Analysis
4.6. Data Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jha, S.N.; Narsaiah, K.; Sharma, A.D.; Singh, M.; Bansal, S.; Kumar, R. Quality parameters of mango and potential of non-destructive techniques for their measurement—A review. J. Food Sci. Technol.-Mysore 2008, 47, 1–14. [Google Scholar] [CrossRef]
- Kushalappa, A.C.; Vikram, A.; Raghavan, G.S.V. Metabolomics of headspace gas for diagnosing diseases of fruits and vegetables after harvest. Stewart Postharvest Rev. 2008, 2, 1–8. [Google Scholar]
- Katrodia, J.S. The Study in to the Causes of the Development of Spongy Tissue in Mango (Mangifera indica L.) Fruit of Cultivar Alphonso; Marathwada Agricultural University: Parbhani, India, 1979. [Google Scholar]
- Cheema, G.S.; Dani, P.G. Report on the Export of Mangoes to Europe in 1932 and 1933; Department of Agriculture: Bombay, India, 1934; pp. 1–31. [Google Scholar]
- Ravindra, V.; Shivshankar, S. Spongy tissue in Alphonso mango. II. A key evidence for the causative role of seed. Curr. Sci. 2006, 91, 1712–1714. [Google Scholar]
- Pandit, S.S.; Chidley, H.G.; Kulkarni, R.S.; Pujari, K.H.; Giri, A.P.; Gupta, V.S. Cultivar relationships in mango based on fruit volatile profiles. Food Chem. 2009, 114, 363–372. [Google Scholar] [CrossRef]
- Janave, M.T.; Sharma, A. Spongy tissue development in Alphonso mango: Association with Staphylococcus xylosus. Eur. J. Plant Pathol. 2008, 122, 335–348. [Google Scholar] [CrossRef]
- Oak, P.S.; Deshpande, A.B.; Pujari, K.H.; Prabhudesai, S.S.; Giri, A.P.; Gupta, V.S. Data on metabolic profiling of spongy tissue disorder in Mangifera indica cv. Alphonso. Data Brief 2019, 22, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Katrodia, J.S.; Rane, D.A.; Salunkhe, D.K. Spongy tissue in mango—Causes and control measures. Acta Hortic. 1988, 231, 815–826. [Google Scholar] [CrossRef]
- Ravindra, V.; Shivashankar, S. Spongy tissue in Alphonso mango—Significance of in situ seed germination events. Curr. Sci. 2004, 87, 1045–1048. [Google Scholar]
- Pandit, S.S.; Kulkarni, R.S.; Chidley, H.G.; Giri, A.P.; Pujari, K.H.; Köllner, T.G.; Degenhardt, J.; Gershenzon, J.; Gupta, V.S. Changes in volatile composition during fruit development and ripening of ‘Alphonso’ mango. J. Sci. Food Agric. 2009, 89, 2071–2081. [Google Scholar] [CrossRef]
- Kulkarni, R.S.; Chidley, H.G.; Pujari, K.H.; Giri, A.P.; Gupta, V.S. Geographic variation in the flavour volatiles of Alphonso mango. Food Chem. 2012, 130, 58–66. [Google Scholar] [CrossRef]
- Kumar, R.; Bohra, A.; Pandey, A.K.; Pandey, M.K.; Kumar, A. Metabolomics for Plant Improvement: Status and Prospects. Front. Plant Sci. 2017, 8, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, E.D.; Barrett, D.; Jobling, J.; Mitcham, E. Biochemical factorsassociated with a CO2-induced flesh browning disorder of Pink Lady apples. Postharvest Biol. Technol. 2008, 48, 182–191. [Google Scholar] [CrossRef]
- Oikawa, A.; Otsuka, T.; Nakabayashi, R.; Jikumaru, Y.; Isuzugawa, K.; Murayama, H.; Saito, K.; Shiratake, K. Metabolic profiling of developing pear fruits reveals dynamic variation in primary and secondary metabolites, including plant hormones. PLoS ONE 2015, 10, e0131408. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhao, H.; Wang, W.; Xu, M.; Shi, J.; Nie, X.; Yang, G. Identification of conserved and diverse metabolic shift of the stylar, intermediate and peduncular segments of cucumber fruit during development. Int. J. Mol. Sci. 2018, 19, 135. [Google Scholar] [CrossRef]
- Liu, X.; Shiomi, S.; Nakatsuka, A.; Kubo, A.; Nakamura, R.; Inaba, A. Characterization of ethylene biosynthesis associated with ripening in banana fruit. Plant Physiol. 1999, 121, 1257–1265. [Google Scholar] [CrossRef]
- Feng, S.; Niu, L.; Suh, J.H.; Hung, W.L.; Wang, Y. Comprehensive metabolomics analysis of mandarins (citrus reticulata) as a tool for variety, rootstock, and grove discrimination. J. Agric. Food Chem. 2018, 66, 10317–10326. [Google Scholar] [CrossRef]
- Ibanez, A.M.; Martinelli, F.; Reagan, R.L.; Uratsu, S.L.; Vo, A.; Tinoco, M.A.; Phu, M.L.; Chen, Y.; Rocke, D.M.; Dandekar, A.M. Transcriptome and metabolome analysis of citrus fruit to elucidate puffing disorder. Plant Sci. 2014, 217–218, 87–98. [Google Scholar] [CrossRef]
- Sumner, L.; Amberg, A.; Barette, D.; Beale, M.; Beger, R.; Daykin, C.; Fan, T.; Fiehn, O.; Goodacre, R.; Groffin, J.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Akihiro, T.; Koike, S.; Tani, R.; Tominaga, T.; Watanabe, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Ashihara, H.; Matsukura, C.; et al. Biochemical mechanism on GABA accumulation during fruit development in tomato. Plant Cell Physiol. 2008, 49, 1378–1389. [Google Scholar] [CrossRef]
- Hanson, A.D.; Beaudoin, G.A.; McCarty, D.R.; Gregory, J.F., 3rd. Does abiotic stress cause functional B Vitamin deficiency in plants? Plant Physiol. 2016, 172, 2082–2097. [Google Scholar] [CrossRef]
- Rahman, M.A.; Akond, M.; Babar, M.A.; Beecher, C.; Erickson, J.; Thomason, K.; De Jong, F.A.; Mason, R.E. LC-HRMS based non-targeted metabolomic profiling of Wheat (Triticum aestivum L.) under post-anthesis drought stress. Am. J. Plant Sci. 2017, 08, 3024–3061. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Lum, G.B.; Shelp, B.J.; DeEll, J.R.; Bozzo, G.G. Oxidative metabolism is associated with physiological disorders in fruits stored under multiple environmental stresses. Plant Sci. 2016, 245, 143–152. [Google Scholar] [CrossRef]
- Benard, C.; Bernillon, S.; Biais, B.; Osorio, S.; Maucourt, M.; Ballias, P.; Deborde, C.; Colombie, S.; Cabasson, C.; Jacob, D.; et al. Metabolomic profiling in tomato reveals diel compositional changes in fruit affected by source-sink relationships. J. Exp. Bot. 2015, 66, 3391–3404. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.K.; Jung, E.S.; Lee, H.A.; Choi, D.; Lee, C.H. Metabolomic characterization of hot pepper (Capsicum annuum “CM334”) during fruit development. J. Agric. Food Chem. 2015, 63, 9452–9460. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, V.; Prabha, T.N.; Tharanathan, R.N. Fruit ripening phenomena--an overview. Crit. Rev. Food Sci. Nutr. 2007, 47, 1–19. [Google Scholar] [CrossRef]
- Vasanthaiah, H.K.; Ravishankar, K.V.; Shivashankara, K.S.; Anand, L.; Narayanaswamy, P.; Mukunda, G.; Prasad, T.G. Cloning and characterization of differentially expressed genes of internal breakdown in mango fruit (Mangifera indica). J. Plant Physiol. 2006, 163, 671–679. [Google Scholar] [CrossRef]
- Shivashankara, K.S.; Mathai, C.K. Relationship of leaf and fruit transpiration rates to the incidence of spongy tissue disorder in two mango (Mangifera indica L.) cultivars. Sci. Hortic. 1999, 82, 317–323. [Google Scholar] [CrossRef]
- Ravindra, V. Recent understanding of spongy tissue problem in ‘Alphonso’ mango—A Review. Acta Hortic. 2015, 1066, 169–175. [Google Scholar] [CrossRef]
- Takayama, M.; Ezura, H. How and why does tomato accumulate a large amount of GABA in the fruit? Front. Plant Sci. 2015, 6, 612. [Google Scholar] [CrossRef]
- Batushansky, A.; Kirma, M.; Grillich, N.; Toubiana, D.; Pham, P.; Balbo, I.; Fromm, H.; Galili, G.; Fernie, A.; Fait, A. Combined transcriptomics and metabolomics of Arabidopsis thaliana seedlings exposed to exogenous GABA suggest its role in plants is predominantly metabolic. Mol. Plant 2014, 7, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.K. Controlled atmosphere storage. In Fruitand Vegetable Storage. Hypobaric, Hyperbaric and Controlled Atmosphere; Hartel, R.W., Ed.; Springer International Publishing: New York, NY, USA, 2016; pp. 21–36. [Google Scholar]
- Deng, J.C.; Yang, C.Q.; Zhang, J.; Zhang, Q.; Yang, F.; Yang, W.Y.; Liu, J. Organ-specific differential NMR-based metabonomic analysis of soybean [Glycine max (L.) Merr.] Fruit Reveals the Metabolic Shifts and Potential Protection Mechanisms Involved in Field Mold Infection. Front. Plant Sci. 2017, 8, 508. [Google Scholar] [CrossRef] [PubMed]
- Nagamani, J.E.; Shivashankara, K.S.; Roy, T.K. Role of oxidative stress and the activity of ethylene biosynthetic enzymes on the formation of spongy tissue in ‘Alphonso’ mango. J. Food Sci. Technol. 2010, 47, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, S.; Fromm, H. Closing the loop on the GABA shunt in plants: Are GABA metabolism and signaling entwined? Front. Plant Sci. 2015, 6, 419. [Google Scholar] [CrossRef]
- Ceusters, J.; Godts, C.; Peshev, D.; Vergauwen, R.; Dyubankova, N.; Lescrinier, E.; De Proft, M.P.; Van den Ende, W. Sedoheptulose accumulation under CO2 enrichment in leaves of Kalanchoe pinnata: A novel mechanism to enhance C and P homeostasis? J. Exp. Bot. 2013, 64, 1497–1507. [Google Scholar] [CrossRef]
- Vasanthaiah, H.K.N.; Ravishankar, K.V.; Narayanaswamy, P.; Shivashankara, K.S. Influence of temperature on spongy tissue formation in ‘alphonso’ mango. Int. J. Fruit Sci. 2008, 8, 226–234. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef]
- Shivashankar, S.; Sumathi, M.; Roy, T.K. Do seed VLCFAs trigger spongy tissue formation in Alphonso mango by inducing germination? J. Biosci. 2015, 40, 375–387. [Google Scholar] [CrossRef]
- Gunjate, R.T.; Tare, S.L.; Rangwala, A.D.; Limaye, V.P. Calcium content in Alphonso mango fruits in relation to occurrence of spongy tissue. J. Maharashtra Agric. Univ. 1979, 4, 159–161. [Google Scholar]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef]
- Ho, Q.T.; Hertog, M.; Verboven, P.; Ambaw, A.; Rogge, S.; Verlinden, B.E.; Nicolai, B.M. Down-regulation of respiration in pear fruit depends on temperature. J. Exp. Bot. 2018, 69, 2049–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peer, M.; Stegmann, M.; Mueller, M.J.; Waller, F. Pseudomonas syringae infection triggers de novo synthesis of phytosphingosine from sphinganine in Arabidopsis thaliana. FEBS Lett. 2010, 584, 4053–4056. [Google Scholar] [CrossRef] [PubMed]
- Ali, U.; Li, H.; Wang, X.; Guo, L. Emerging roles of sphingolipid signaling in plant response to biotic and abiotic stresses. Mol. Plant 2018, 11, 1328–1343. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.F.; Chai, Y.M.; Li, C.L.; Lu, D.; Luo, J.J.; Qin, L.; Shen, Y.Y. Abscisic acid plays an important role in the regulation of strawberry fruit ripening. Plant Physiol. 2011, 157, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Zhao, F.; Li, R.; Xiao, H. Cross-talk modulation between ABA and ethylene by transcription factor SlZFP2 during fruit development and ripening in tomato. Plant Signal. Behav. 2015, 10, e1107691. [Google Scholar] [CrossRef]
- Chidley, H.G.; Deshpande, A.B.; Oak, P.S.; Pujari, K.H.; Giri, A.P.; Gupta, V.S. Effect of postharvest ethylene treatment on sugar content, glycosidase activity and its gene expression in mango fruit. J. Sci. Food Agric. 2017, 97, 1624–1633. [Google Scholar] [CrossRef]
- Yashoda, H.M.; Prabha, T.N.; Tharanathan, R.N. Mango ripening--chemical and structural characterization of pectic and hemicellulosic polysaccharides. Carbohydr. Res. 2005, 340, 1335–1342. [Google Scholar] [CrossRef]
- Kasangana, P.B.; Haddad, P.S.; Stevanovic, T. Study of polyphenol content and antioxidant capacity of Myrianthus arboreus (cecropiaceae) root bark extracts. Antioxidants 2015, 4, 410–426. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V. Total Phenol Analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Dalton, D.; Russell, R.; Hanus, F.; Pascoet, J.; Evans, H. Enzymatic reactions of ascorbate and glutathione that prevent peroxide damage in soybean root nodules. Proc. Natl. Acad. Sci. USA 1986, 83, 3811–3815. [Google Scholar] [CrossRef] [Green Version]
- Deo, R.; Barvkar, V.; Nadaf, A.; Mundhe, S.; Kadoo, N. Integrative omics analysis in Pandanus odorifer (Forssk.) Kuntze reveals the role of Asparagine synthetase in salinity tolerance. Sci. Rep. 2019, 9, 932. [Google Scholar]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acid Res. 2018, 46, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Metabolomic data processing, analysis, and interpretation using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 14, 1–48. [Google Scholar] [CrossRef]
- Pandit, S. Genetic Analysis of Alphonso Mango Flavor Biogenesis; University of Pune, CSIR-National Chemical Laboratory: Pune, Maharashtra, India, 2008. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative Metabolite | Formula | KEGG | No. of Fragment Matches | RT (min) | Probable Mass | Exact Mass | Adduct Annotation | ESI Mode |
|---|---|---|---|---|---|---|---|---|
| 1-Aminocyclopropane-1-carboxylic acid | C6H7NO2 | C01234 | 5 | 3.02 | 101.0481 | 101.0477 | [M+H]+ | Positive |
| γ-Aminobutyric acid a | C4H9NO2 | C00334 | 6 | 3.02 | 103.0638 | 103.0633 | [M+H]+ | Positive |
| Uracil | C4H4N2O2 | C00106 | 4 | 5.06 | 112.0276 | 112.0273 | [M+H]+ | Positive |
| l-Proline | C5H9NO2 | C00148 | 11 | 3.12 | 115.0636 | 115.0633 | [M+H]+ | Positive |
| l-Threonine | C4H9NO3 | C00188 | 8 | 3 | 119.0584 | 119.0582 | [M+H]+ | Positive |
| Pyroglutamate | C5H7NO3 | C01879 | 15 | 4.62 | 129.0426 | 129.0426 | [M+H]+ | Positive |
| Aspartate | C4H7NO4 | C00049 | 6 | 4.6 | 133.036 | 133.0375 | - | Positive |
| Spermidine | C7H19N3 | C00315 | 9 | 2.82 | 145.1579 | 145.1579 | [M+H]+ | Positive |
| α-Ketoglutaric acid | C5H6O5 | C00026 | 7 | 4.61 | 146.0215 | 146.0215 | [M+H]+ | Positive |
| l-Lysine | C6H14N2O2 | C00047 | 8 | 2.81 | 146.1055 | 146.1055 | [M+H]+ | Positive |
| (S)-2-Hydroxyglutarate | C5H8O5 | C03196 | 10 | 4.59 | 148.036 | 148.0372 | [M-H]− | Negative |
| Methionine | C5H11NO2S | C00073 | 9 | 4.13 | 149.051 | 149.051 | [M+H]+ | Positive |
| Histidine | C6H9N3O2 | C00135 | 12 | 2.83 | 155.0695 | 155.0695 | [M+H]+ | Positive |
| DL-2-Aminoadipate | C6H11NO4 | C00956 | 24 | 3.4 | 161.0688 | 161.0688 | [M+H]+ | Positive |
| 2-Hydroxyadipate | C6H10O5 | C02360 | 25 | 3.14 | 162.0527 | 162.0528 | [M+H]+ | Positive |
| Phenylalanine | C9H11NO2 | C00079 | 20 | 6.77 | 165.079 | 165.079 | [M+H]+ | Positive |
| Dehydroascorbic acid | C6H6O6 | C05422 | 7 | 4.02 | 174.0161 | 174.0164 | [M+H]+ | Positive |
| l-Arginine | C6H14N4O2 | C00062 | 19 | 2.89 | 174.1111 | 174.1117 | [M+H]+ | Positive |
| Ascorbic acid | C6H8O6 | C00072 | 24 | 3.7 | 176.0321 | 176.0321 | [M+H]+ | Positive |
| D-Glucose | C6H12O6 | C00031 | 11 | 3.69 | 180.0624 | 180.0634 | [M+H]+ | Positive |
| Tyrosine | C9H11NO3 | C00082 | 17 | 3.38 | 181.0734 | 181.0739 | [M+H]+ | Positive |
| Isocitric acid | C6H8O7 | C00311 | 16 | 3.66 | 192.0267 | 192.027 | [M-H]− | Negative |
| ferulic acid | C10H10O4 | C01494 | 5 | 7.78 | 194.0583 | 194.0579 | [M+H]+ | Positive |
| N2-Acetyl-l-aminoadipate | C18H13NO5 | C12986 | 35 | 3.06 | 203.0792 | 203.0794 | [M+H]+ | Positive |
| l-Tryptophan | C11H12N2O2 | C00078 | 10 | 7.95 | 204.0899 | 204.0899 | [M+H]+ | Positive |
| Indole acetic acid | C11H11NO3 | C02043 | 6 | 10.1 | 205.0734 | 205.0739 | [M+H]+ | Positive |
| Sedoheptulose | C7H14O7 | C02076 | 19 | 3.23 | 210.0737 | 210.074 | - | Positive |
| Succinylproline | C9H13NO5 | C11711 | 35 | 3.22 | 215.0788 | 215.0794 | [M+H]+ | Positive |
| Pyridoxal Phosphate | C8H10NO6P | C00018 | 1 | 4.62 | 247.0245 | 247.0246 | [M+H]+ | Positive |
| (+)-Abscisic Acid | C15H20O4 | C06082 | 18 | 12.69 | 264.1362 | 264.1362 | [M-H]− | Negative |
| Deidaclin | C12H17NO6 | C08329 | 34 | 3.1 | 271.1032 | 271.1056 | [M+H]+ | Positive |
| Ranunculin | C11H16O8 | C08512 | 19 | 3.08 | 276.082 | 276.0845 | [M+H]+ | Positive |
| Kaempferol | C15H10O6 | C05903 | 2 | 6.24 | 286.0479 | 286.0477 | [M+H]+ | Positive |
| Sphinganine | C18H39NO2 | C00836 | 8 | 14.45 | 301.2979 | 301.2981 | [M+H]+ | Positive |
| Glutathione reduced | C10H17N3O6S | C00051 | 22 | 4.72 | 307.0813 | 307.0838 | [M+H]+ | Positive |
| Phytosphingosine | C18H39NO3 | C12144 | 18 | 14.19 | 317.2929 | 317.293 | [M+H]+ | Positive |
| 4-O-β-d-glucosyl-4-hydroxycinnamate | C15H18O8 | C04415 | 11 | 7.38 | 326.1002 | 326.1002 | [M+2Na-H]+ | Negative |
| 9,10-Dihydroxy-12,13-epoxyoctadecanoic acid | C18H34O5 | C14837 | 11 | 13.3 | 330.2408 | 330.2406 | [M-H]- | Negative |
| 5′-O-β-d-glucosyl-pyridoxine | C14H21NO8 | C03996 | 13 | 4.48 | 331.1265 | 331.1267 | [M+H]+ | Positive |
| Sucrose | C12H22O11 | C00089 | 17 | 3.82 | 342.1162 | 342.1162 | [M-H]− | Negative |
| Chlorogenic acid # | C16H18O9 | C00852 | 15 | 3.45 | 354.09 | 354.0951 | [M-H]− | Negative |
| 1-Feruloyl-d-glucose | C16H20O9 | C17759 | 10 | 8.99 | 356.1092 | 356.1107 | [M-H]− | Negative |
| 3-O-Feruloylquinic acid | C17H20O9 | C02572 | 14 | 3.08 | 368.1123 | 368.1107 | [M-H]− | Negative |
| Vernoflexuoside | C21H28O8 | C09579 | 6 | 9.07 | 408.176 | 408.1784 | [M+H]+ | Positive |
| Mangiferin | C19H18O11 | C10077 | 14 | 8.61 | 422.0848 | 422.0849 | [M+H]+ | Positive |
| Glutathione oxidized | C20H32N6O12S2 | C00127 | 14 | 5.16 | 612.1518 | 612.152 | [M+2H]+ | Positive |
| Metabolite | Fold Change in Healthy Tissue | Fold Change in Spongy Control Tissue | Fold Change in Spongy Tissue | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Table Green | Mid Ripe | Ripe | Table Green | Mid Ripe | Ripe | Mature Raw | Table Green | Mid Ripe | Ripe | |
| 1-Aminocyclopropane-1-carboxylic acid | 1.6 | 1.17 | 1.86 | 0.53 | 1.13 | 2.22 | 0.91 | 0.47 | 0.31 | 0.63 |
| γ-Aminobutyric acid | 1.03 | 0.19 | 0.35 | 0.41 | 0.27 | 0.27 | 1.53 | 2.28 | 0.72 | 1.03 |
| Uracil | 5.06 | 4.23 | 10 | 4.43 | 11.5 | 18.4 | 0.91 | 47.63 | 14.08 | 16.49 |
| l-Proline | 1.34 | 0.29 | 0.34 | 0.29 | 0.25 | 0.4 | 1.54 | 0.86 | 0.4 | 0.88 |
| l-Threonine | 1.72 | 0.58 | 1.32 | 0.56 | 0.61 | 1.06 | 1.41 | 1.21 | 0.68 | 1.23 |
| Pyroglutamate | 1.05 | 0.22 | 0.23 | 0.2 | 0.21 | 0.27 | 1.16 | 5.38 | 5.35 | 4.69 |
| Aspartate | 1.43 | 0.78 | 0.8 | 0.34 | 0.45 | 0.49 | 0.84 | 0.11 | 0.18 | 0.19 |
| Spermidine | 1.43 | 0.12 | 0.83 | 0.07 | 0.28 | 0.52 | 0.76 | 0.32 | 0.1 | 0.17 |
| α-Ketoglutarate | 0.98 | 0.13 | 0.1 | 0.17 | 0.06 | 0.07 | 1.01 | 0.72 | 0.48 | 0.62 |
| l-Lysine | 0.78 | 0.18 | 0.22 | 0.2 | 0.19 | 0.25 | 0.87 | 1.39 | 0.62 | 0.64 |
| 2-Hydroxyglutarate | 2.15 | 0.65 | 0.58 | 0.95 | 0.36 | 0.75 | 1.45 | 1.65 | 1.81 | 2.81 |
| Methionine | 0.73 | 0.24 | 0.25 | 0.36 | 0.32 | 0.44 | 0.8 | 1.24 | 0.38 | 0.43 |
| Histidine | 1.57 | 1.29 | 5.44 | 0.57 | 1.6 | 3.36 | 1.6 | 1.43 | 0.77 | 1.18 |
| DL-2-Aminoadipate | 1.31 | 0.19 | 0.44 | 0.08 | 0.21 | 0.33 | 1.55 | 0.31 | 0.14 | 0.22 |
| 2-Hydroxyadipic acid | 2.4 | 2.66 | 4.62 | 3.41 | 3.48 | 3.96 | 0.9 | 1.83 | 2.25 | 1.65 |
| Phenylalanine | 0.51 | 0.07 | 0.04 | 0.06 | 0.04 | 0.04 | 1.08 | 0.8 | 0.22 | 0.31 |
| Dehydroascorbic acid | 2.04 | 2.26 | 2.79 | 2.28 | 1.65 | 2.33 | 1.28 | 3.47 | 1.9 | 3.58 |
| l-Arginine | 0.98 | 0.02 | 0 | 0.03 | 0.01 | 0.01 | 1.11 | 0.87 | 0.28 | 0.17 |
| Ascorbic acid | 0.73 | 0.22 | 1.31 | 0.63 | 0.28 | 1.62 | 1.01 | 0.14 | 0.05 | 0.23 |
| D-Glucose | 1.07 | 0.44 | 0.67 | 0.48 | 2.22 | 0.69 | 0.83 | 4.4 | 5.42 | 1.24 |
| Tyrosine | 1.51 | 0.4 | 0.86 | 0.73 | 0.66 | 1.47 | 1.32 | 2.47 | 1.92 | 2.65 |
| Isocitric acid | 2.69 | 0.48 | 0.31 | 0.84 | 0.25 | 0.3 | 1.66 | 1.87 | 1.81 | 1.94 |
| ferulic acid | 1.85 | 1.2 | 2.53 | 1.55 | 1.98 | 3.44 | 1.22 | 2.05 | 2.28 | 3.98 |
| N2-Acetyl-L-aminoadipate | 1.85 | 1.36 | 3.19 | 1.95 | 2.4 | 4.53 | 0.94 | 2.52 | 2.6 | 4.26 |
| l-Tryptophan | 1.81 | 0.43 | 0.84 | 0.39 | 0.52 | 1.47 | 1.37 | 2.33 | 0.9 | 1.81 |
| Indole acetic acid | 1.55 | 0.37 | 0.59 | 0.4 | 0.47 | 1.75 | 1.28 | 2.39 | 0.87 | 2.07 |
| Sedoheptulose | 1.51 | 1.87 | 5.05 | 4.11 | 6.59 | 14.31 | 1.76 | 1.03 | 2.29 | 3.32 |
| Succinylproline | 1.54 | 0.59 | 1.39 | 0.91 | 1.44 | 2.01 | 1.13 | 1.89 | 1.32 | 2.34 |
| Pyridoxal Phosphate | 0.99 | 0.43 | 0.45 | 0.53 | 0.27 | 0.38 | 1.04 | 0.98 | 0.84 | 1.45 |
| (+)-Abscisic Acid | 4.03 | 4.12 | 2.83 | 2.23 | 1.54 | 2.1 | 1.13 | 1.21 | 0.44 | 0.35 |
| Deidaclin | 1.87 | 0.59 | 1.53 | 1.86 | 2.74 | 4.53 | 1.02 | 1.09 | 7.69 | 90.34 |
| Ranunculin | 0.97 | 0.86 | 1.7 | 1.47 | 0.76 | 1.71 | 0.91 | 1.54 | 1.59 | 3.34 |
| Kaempherol | 1.05 | 0.65 | 1.5 | 0.47 | 0.82 | 1.47 | 0.88 | 0.28 | 0.46 | 1.89 |
| Sphinganine | 1.23 | 0.46 | 0.01 | 0.16 | 0.21 | 0.01 | 0.21 | 0.21 | 0.22 | 0.01 |
| Glutathione reduced | 1.99 | 0.31 | 0.44 | 0.52 | 0.32 | 0.34 | 1.11 | 0.23 | 0.03 | 0.05 |
| Phytosphingosine | 1.08 | 0.35 | 0.12 | 0.47 | 0.26 | 0.1 | 1.13 | 1.48 | 1.25 | 0.3 |
| 4-O-β-d-glucosyl-4-hydroxycinnamate | 1.27 | 1.36 | 2.97 | 1.65 | 1.26 | 4.24 | 1.33 | 1.32 | 1.5 | 2.22 |
| 9,10-Dihydroxy-12,13-epoxyoctadecanoic acid | 1.07 | 0.57 | 0.69 | 1.16 | 0.83 | 2.01 | 1.01 | 3.37 | 35.87 | 132.76 |
| 5′-O-β-D-glucosyl-pyridoxine | 2.08 | 2.8 | 6.8 | 6.13 | 7.86 | 12.65 | 1.18 | 7.51 | 27.24 | 92.21 |
| Sucrose | 3.8 | 8.23 | 9.38 | 5.65 | 11.74 | 13.56 | 0.84 | 3.54 | 2.47 | 1.44 |
| Chlorogenic acid | 0.8 | 1.14 | 10.8 | 1.8 | 1.26 | 15.04 | 1.22 | 1.47 | 2.68 | 19.97 |
| 1-Feruloyl-d-glucose | 0.81 | 0.41 | 0.01 | 0.45 | 0.23 | 0.01 | 0.72 | 0.01 | 0.02 | 0.04 |
| 3-O-Feruloylquinate | 1.15 | 1.67 | 6.8 | 1.75 | 0.8 | 4.55 | 1.5 | 0.32 | 0.74 | 5.51 |
| Vernoflexuoside | 2.76 | 3.98 | 20.19 | 2.67 | 4.58 | 14.42 | 1.47 | 5.72 | 4.45 | 24.35 |
| Mangiferin | 0.56 | 0.37 | 0.57 | 0.32 | 0.43 | 1.22 | 0.44 | 1.06 | 1.84 | 1.3 |
| Glutathione oxidized | 1.56 | 1.33 | 0.19 | 0.24 | 0.44 | 0.27 | 1.86 | 4.91 | 1.76 | 0.67 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oak, P.; Deshpande, A.; Giri, A.; Gupta, V. Metabolomic Dynamics Reveals Oxidative Stress in Spongy Tissue Disorder During Ripening of Mangifera indica L. Fruit. Metabolites 2019, 9, 255. https://doi.org/10.3390/metabo9110255
Oak P, Deshpande A, Giri A, Gupta V. Metabolomic Dynamics Reveals Oxidative Stress in Spongy Tissue Disorder During Ripening of Mangifera indica L. Fruit. Metabolites. 2019; 9(11):255. https://doi.org/10.3390/metabo9110255
Chicago/Turabian StyleOak, Pranjali, Ashish Deshpande, Ashok Giri, and Vidya Gupta. 2019. "Metabolomic Dynamics Reveals Oxidative Stress in Spongy Tissue Disorder During Ripening of Mangifera indica L. Fruit" Metabolites 9, no. 11: 255. https://doi.org/10.3390/metabo9110255
APA StyleOak, P., Deshpande, A., Giri, A., & Gupta, V. (2019). Metabolomic Dynamics Reveals Oxidative Stress in Spongy Tissue Disorder During Ripening of Mangifera indica L. Fruit. Metabolites, 9(11), 255. https://doi.org/10.3390/metabo9110255