Effects of Copper and pH on the Growth and Physiology of Desmodesmus sp. AARLG074


Abstract
:1. Introduction
2. Results
2.1. Copper Removal Efficiency
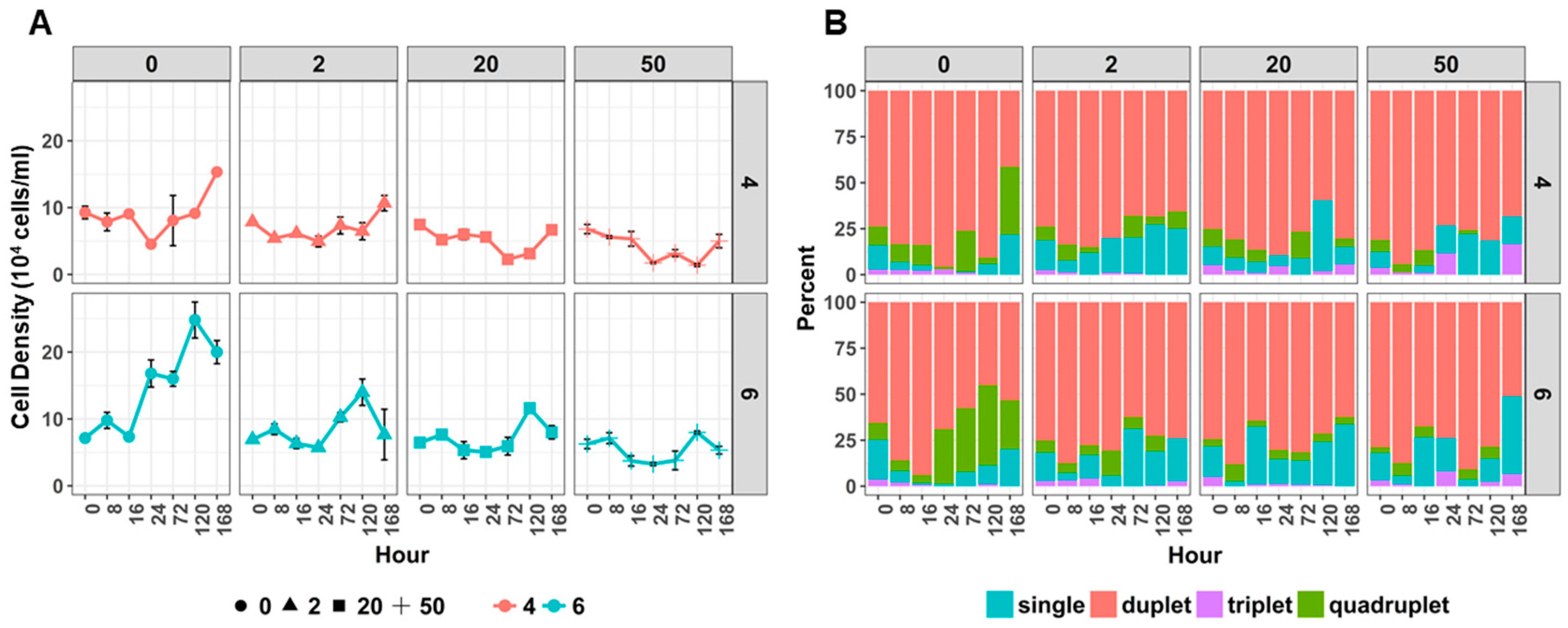
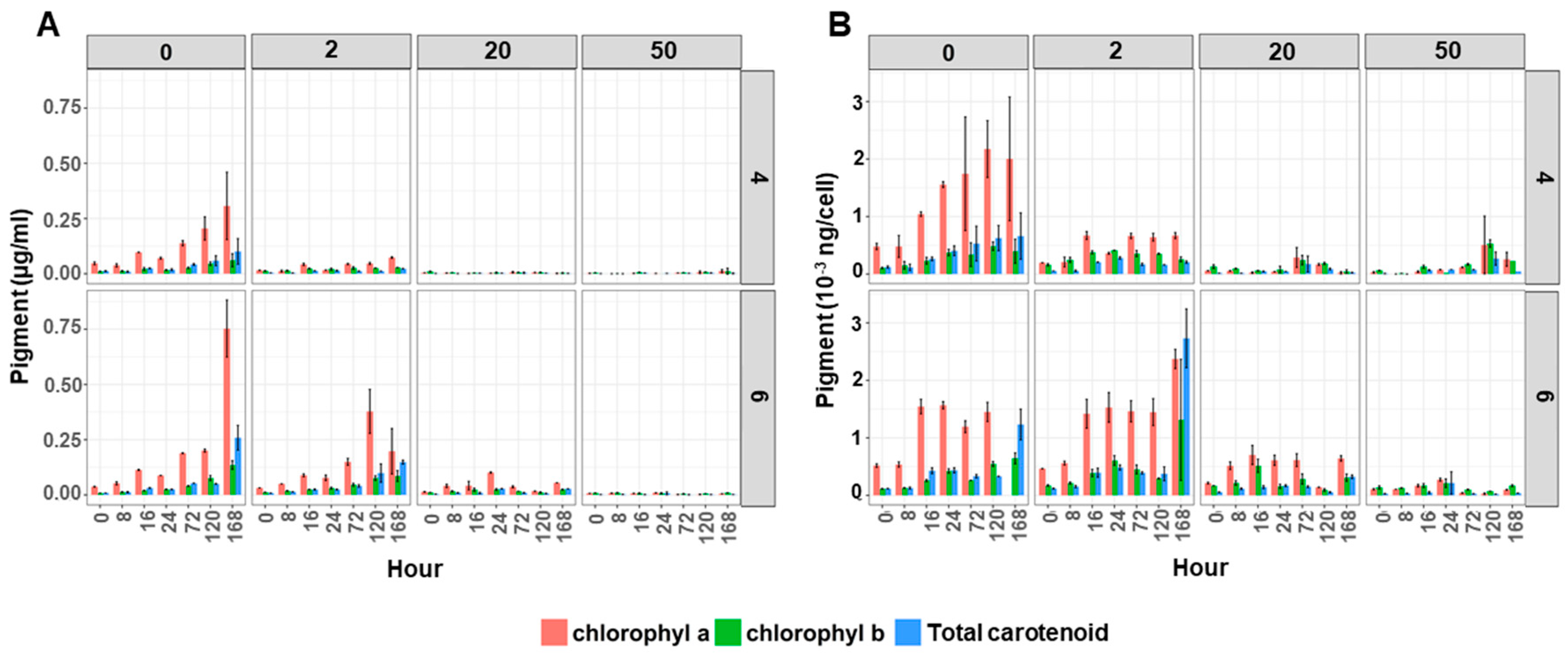
2.2. Copper and Acid Exposure Affected the Cell Density, Pigment Content and Ultrastructure in Desmodesmus sp. AARLG074
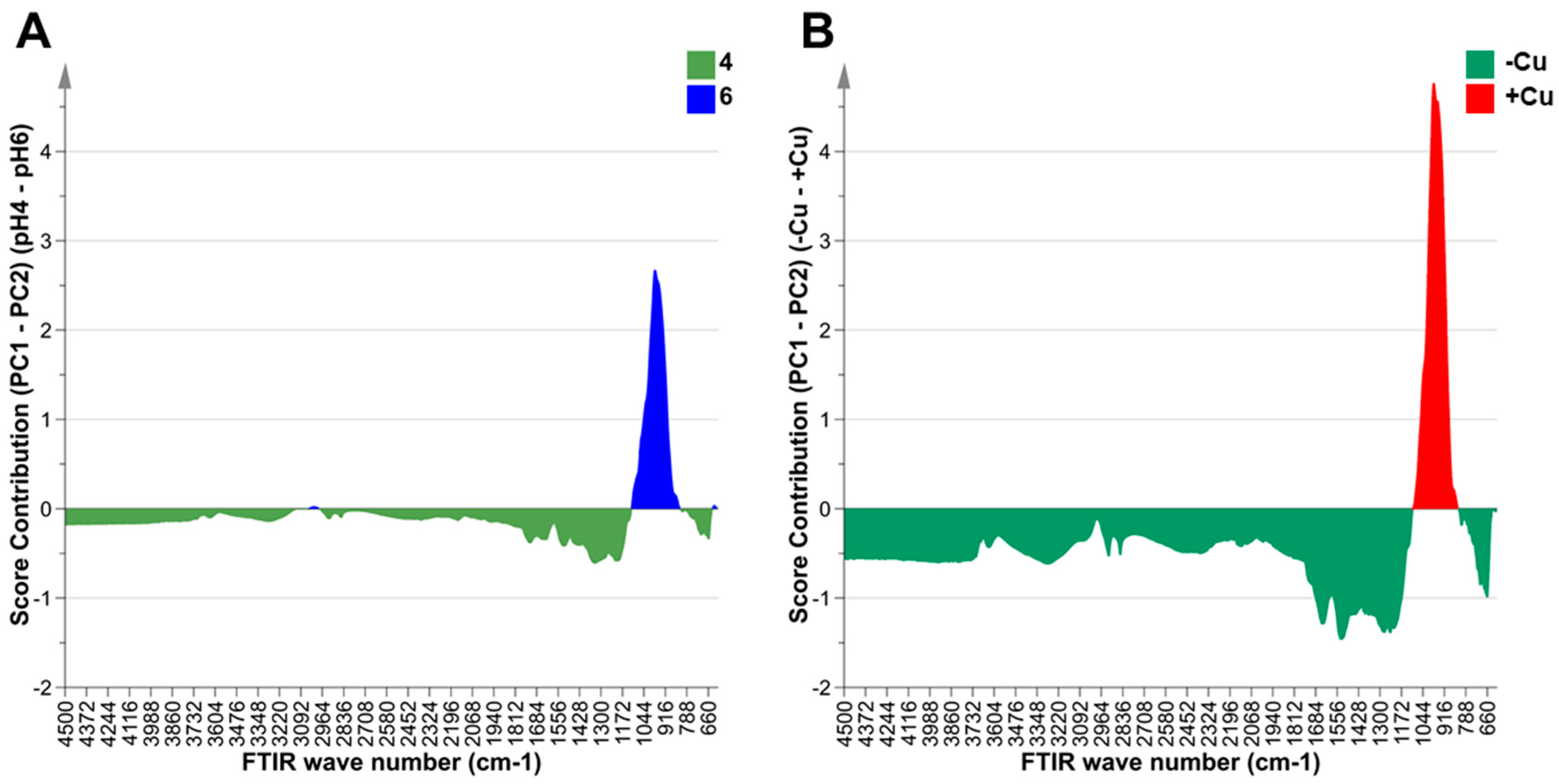
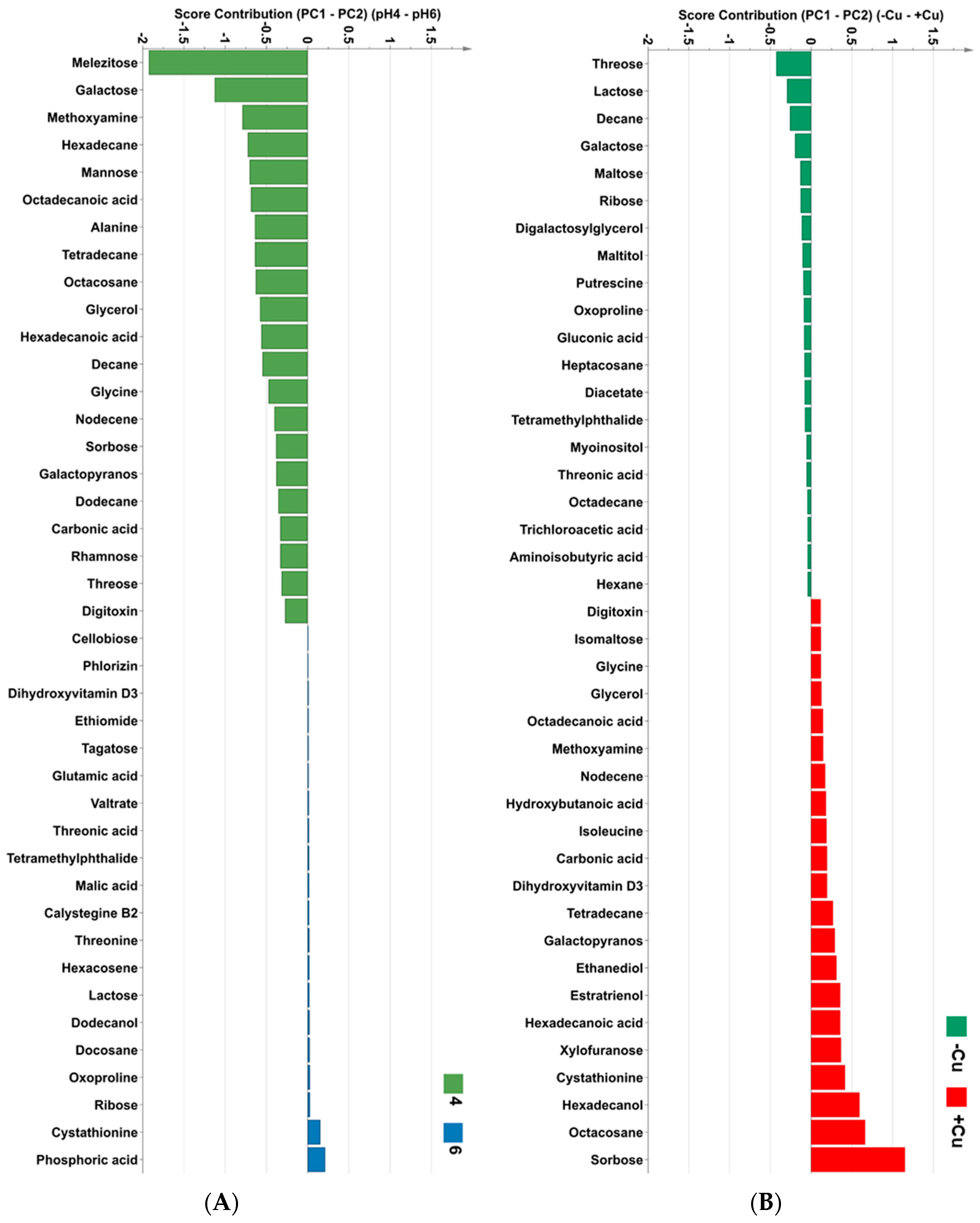
2.3. The Effects of Copper and pH on Metabolite Composition of Desmodesmus sp. AARLG074
3. Discussion
3.1. Tolerance of High Cu Exposure and Acidity in Desmodesmus sp. AARLG074—Growth and Structural Alterations Associated with Cu Removal
3.2. Metabolic Alterations Associated with Exposure to Cu and Acidity in Desmodesmus sp. AARLG074
4. Materials and Methods
4.1. Organism and Culture Conditions
4.2. Copper Absorption Efficiency
4.3. The Effects of Copper and pH on Cell Density, Pigments and Ultrastructure
4.4. Metabolite Analysis
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Al-saydeh, S.A.; El-naas, M.H.; Zaidi, S.J. Copper removal from industrial wastewater: A comprehensive review. J. Ind. Eng. Chem. 2017, 56, 35–44. [Google Scholar] [CrossRef]
- Elshkaki, A.; Graedel, T.E.; Ciacci, L.; Reck, B.K. Copper demand, supply, and associated energy use to 2050. Glob. Environ. Chang. 2016, 39, 305–315. [Google Scholar] [CrossRef]
- Breedveld, G.D.; Klimpel, F.; Kvennås, M.; Okkenhaug, G. Stimulation of natural attenuation of metals in acid mine drainage through water and sediment management at abandoned copper mines. In Proceedings of the International Mine Water Association 2017 Congress (IMWA 2017)-Mine Water and Circular Economy, Lappeenrante, Finland, 25–30 June 2017; pp. 1133–1137. [Google Scholar]
- Liang-qi, L.; Ci-an, S. Acid mine drainage and heavy metal contamination in groundwater of metal sulfide mine at arid territory (BS mine, Western Australia). Trans. Nonferrous Met. Soc. China 2010, 20, 1488–1493. [Google Scholar]
- Ehrlich, H.L. Microorganisms in Acid Drainage from a Copper Mine. J. Bacteriol. 1963, 86, 350–352. [Google Scholar]
- Basha, C.A.; Bhadrinarayana, N.S.; Anantharaman, N.; Sheriffa, K.M.M. Heavy metal removal from copper smelting effluent using electrochemical cylindrical flow reactor. J. Hazard Mater. 2008, 152, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.; Nosrati, A.; Kaur, S.; Wagner, J.; Baus, U.; Nydén, M. Copper removal from acid mine drainage-polluted water using glutaraldehyde-polyethyleneimine modified diatomaceous earth particles. Heliyon 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Pang, Q.; Zhang, A.; Lin, L.; Li, N.; Yan, X. Excess copper induced proteomic changes in the marine brown algae Sargassum fusiforme. Ecotoxicol. Environ. Saf. 2015, 111, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Anu, P.R.; Bijoy Nandan, S.; Jayachandran, P.R.; Don Xavier, N.D. Toxicity effects of copper on the marine diatom, Chaetoceros calcitrans. Reg. Stud. Mar. Sci. 2016, 8, 498–504. [Google Scholar] [CrossRef]
- Debelius, B.; Forja, J.M.; DelValls, Á.; Lubián, L.M. Toxicity and bioaccumulation of copper and lead in five marine microalgae. Ecotoxicol. Environ. Saf. 2009, 72, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.L.; Stauber, J.L.; Adams, M.S.; Jolley, D.F. Copper and zinc tolerance of two tropical microalgae after copper acclimation. Environ. Toxicol. 2007, 22, 234–244. [Google Scholar] [CrossRef]
- Singh, R.; Gautam, N.; Mishra, A.; Gupta, R. Heavy metals and living systems: An overview. Indian J. Pharmacol. 2011, 43, 246–253. [Google Scholar] [CrossRef]
- Usak, V.I.H. Copper and copper-containing pesticides: Metabolisms, toxicity and oxidative stress. J. Vasyl. Stefanyk Precarpathian Natl. Univ. 2015, 2, 38–50. [Google Scholar]
- Hill, K.; Hassett, R.; Kosman, D.; Merchant, S. Regulated copper uptake in Chlamydomonas reinhardtií in response to copper availability. Plant Physiol. 1996, 112, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D. Absorption and Adsorption of Heavy Metals by Microalgae. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2nd ed.; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Oxford, UK, 2013; pp. 602–611. [Google Scholar] [CrossRef]
- Levy, J.L.; Angel, B.M.; Stauber, J.L.; Poon, W.L.; Simpson, S.L.; Cheng, S.H.; Jolley, D.F. Uptake and internalisation of copper by three marine microalgae: Comparison of copper-sensitive and copper-tolerant species. Aquat. Biosyst. 2008, 89, 82–93. [Google Scholar] [CrossRef]
- Cao, D.J.; Xie, P.P.; Deng, J.W.; Zhang, H.M.; Ma, R.X.; Liu, C.; Liu, R.; Liang, Y.; Li, H.; Shi, X. Effects of Cu2+and Zn2+on growth and physiological characteristics of green algae, Cladophora. Environ. Sci. Pollut. Res. 2015, 22, 16535–16541. [Google Scholar] [CrossRef]
- Sabatini, S.E.; Juárez, Á.B.; Eppis, M.R.; Bianchi, L.; Luquet, C.M.; Ríos de Molina M del, C. Oxidative stress and antioxidant defenses in two green microalgae exposed to copper. Ecotoxicol. Environ. Saf. 2009, 72, 1200–1206. [Google Scholar] [CrossRef]
- Volland, S.; Bayer, E.; Baumgartner, V.; Andosch, A.; Lütz, C.; Sima, E.; Lütz-Meindl, U. Rescue of heavy metal effects on cell physiology of the algal model system Micrasterias by divalent ions. J. Plant Physiol. 2014, 171, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Phinyo, K.; Pekkoh, J.; Peerapornpisal, Y. Distribution and ecological habitat of Scenedesmus and related genera in some freshwater resources of Northern and North-Eastern Thailand. Biodiversitas 2017, 18, 1092–1099. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, 486–494. [Google Scholar] [CrossRef]
- Kleinübing, S.J.; Vieira, R.S.; Beppu, M.M.; Guibal, E.; da Silva, M.G.C. Characterization and evaluation of copper and nickel biosorption on acidic algae Sargassum Filipendula. Mater. Res. 2010, 13, 541–550. [Google Scholar] [CrossRef]
- Al-Homaidan, A.A.; Al-Houri, H.J.; Al-Hazzani, A.A.; Elgaaly, G.; Moubayed, N.M.S. Biosorption of copper ions from aqueous solutions by Spirulina a biomass. Arab. J. Chem. 2014, 7, 57–62. [Google Scholar] [CrossRef]
- Rangsayatorn, N.; Upatham, E.S.; Kruatrachue, M.; Pokethitiyook, P.; Lanza, G.R. Phytoremediation potential of Spirulina (Arthrospira) platensis: Biosorption and toxicity studies of cadmium. Environ. Pollut. 2002, 119, 45–53. [Google Scholar] [CrossRef]
- Wilde, K.L.; Stauber, J.L.; Markich, S.J.; Franklin, N.M.; Brown, P.L. The effect of pH on the uptake and toxicity of copper and zinc in a tropical freshwater alga (Chlorella sp.). Arch. Environ. Contam. Toxicol. 2006, 51, 174–185. [Google Scholar] [CrossRef]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R. Potential use of algae for heavy metal bioremediation, a critical review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kováčik, J.; Klejdus, B.; Hedbavny, J.; Bačkor, M. Effect of copper and salicylic acid on phenolic metabolites and free amino acids in Scenedesmus quadricauda (Chlorophyceae). Plant Sci. 2010, 178, 307–311. [Google Scholar] [CrossRef]
- Kondzior, P.; Butarewicz, A. Effect of heavy metals (Cu and Zn) on the content of photosynthetic pigments in the cells of algae Chlorella vulgaris. J. Ecol. Eng. 2018, 19, 18–28. [Google Scholar] [CrossRef]
- Rodrigo, W.; Schmidt, É.C.; Felix, M.R.D.L.; Polo, L.K.; Kreusch, M.; Pereira, D.T.; Costa, G.B.; Simioni, C.; Chow, F.; Ramlov, F.; et al. Bioabsorption of cadmium, copper and lead by the red macroalga Gelidium floridanum: Physiological responses and ultrastructure features. Ecotoxicol. Environ. Saf. 2014, 105, 80–89. [Google Scholar] [CrossRef]
- Surosz, W.; Palinska, K.A. Ultrastructural changes induced by selected cadmium and copper concentrations in the cyanobacterium Phormidium: Interaction with salinity. J. Plant Physiol. 2000, 157, 643–650. [Google Scholar] [CrossRef]
- Rachlin, J.W.; Jensen, T.E.; Baxter, M.; Jani, V. Utilization of Morphometric Analysis in Evaluating Response of Plectonema boryanum (Cyanophyceae) to Exposure to Eight Heavy Metals. Arch. Environ. Contam. Toxicol. 1982, 11, 323–333. [Google Scholar]
- Nie, M.; Wang, Y.; Yu, J.; Xiao, M.; Jiang, L.; Yang, J.; Fang, C.; Chen, J.; Li, B. Understanding Plant-Microbe Interactions for Phytoremediation of Petroleum-Polluted Soil. PLoS ONE 2011, 6, e17961. [Google Scholar] [CrossRef]
- Marchand, J.; Heydarizadeh, P.; Schoefs, B.; Spetea, C. Ion and metabolite transport in the chloroplast of algae: Lessons from land plants. Cell. Mol. Life Sci. 2018, 75, 2153–2176. [Google Scholar] [CrossRef] [PubMed]
- Kropat, J.; Gallaher, S.D.; Urzica, E.I.; Nakamoto, S.S.; Strenkert, D.; Tottey, S.; Andrew ZMason, A.Z.; Merchant, S.S. Copper economy in Chlamydomonas: Prioritized allocation and reallocation of copper to respiration vs. photosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 2644–2651. [Google Scholar] [CrossRef]
- Pfeil, B.E.; Schoefs, B.; Spetea, C. Function and evolution of channels and transporters in photosynthetic membranes. Cell. Mol. Life Sci. 2014, 71, 979–998. [Google Scholar] [CrossRef] [PubMed]
- Cid, A.; Herrero, C.; Torres, E.; Abalde, J. Copper toxicity on the marine microalga Phaeodactylum tricornutum: Effects on photosynthesis related parameters. Aquat. Toxicol. 1995, 31, 165–174. [Google Scholar] [CrossRef]
- Andosch, A.; Höftberger, M.; Lütz, C.; Lütz-meindl, U. Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii. Int. J. Mol. Sci. 2015, 16, 10389–10410. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, K.; Yamakoshi, Y.; Uemura, I.; Tominaga, N. Ultrastructural changes in Chlamydomonas acidophila (Chlorophyta) induced by heavy metals and polyphosphate metabolism. FEMS Microbiol. Ecol. 2003, 44, 253–259. [Google Scholar] [CrossRef]
- Benning, L.G.; Phoenix, V.R.; Yee, N.; Tobin, M.J. Molecular characterization of cyanobacterial silicification using synchrotron infrared micro-spectroscopy. Geochim. Cosmochim. Acta 2004, 68, 729–741. [Google Scholar] [CrossRef]
- Mayers, J.J.; Flynn, K.J.; Shields, R.J. Rapid determination of bulk microalgal biochemical composition by Fourier-Transform Infrared spectroscopy. Bioresour. Technol. 2013, 148, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, A.; Dittami, S.M.; Goulitquer, S.; Correa, J.A.; Boyen, C.; Potin, P. Transcriptomic and metabolomic analysis of copper stress acclimation in Ectocarpus siliculosus highlights signaling and tolerance mechanisms in brown algae. BMC Plant Biol. 2014, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Demmig-adams, B.; Burch, T.A.; Stewart, J.J.; Savage, E.L.; Iii, W.W.A. Algal glycerol accumulation and release as a sink for photosynthetic electron transport. Algal Res. 2017, 21, 161–168. [Google Scholar] [CrossRef]
- Thompson, G.A.J. Lipids and membrane function in green algae. Biochim. Biophys. Acta 1996, 1302, 17–45. [Google Scholar] [CrossRef]
- Pocivavsek, L.; Gavrilov, K.; Cao, K.D.; Chi, E.Y.; Li, D.; Lin, B.; Meron, M.; Majewski, J.; Lee, K.Y. Glycerol-Induced Membrane Stiffening: The Role of Viscous Fluid Adlayers. Biophys. J. 2011, 101, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Gross, W. Ecophysiology of algae living in highly acidic environments. Hydrobiologia 2000, 433, 31–37. [Google Scholar] [CrossRef]
- Eick, K.C. Metabolomic Analysis of the Allelopathic Interactions between Marine Planktonic Microalgae. Ph.D. Dissertation, Friedrich-Schiller-Universität Jena, Jena, Germany, 1987. [Google Scholar]
- Zhao, X.; Wang, W.; Zhang, F.; Deng, J.; Li, Z.; Fu, B. Comparative Metabolite Profiling of Two Rice Genotypes with Contrasting Salt Stress Tolerance at the Seedling Stage. PLoS ONE 2014, 9, 1–7. [Google Scholar] [CrossRef]
- Vílchez, J.I.; Niehaus, K.; Dowling, D.N.; González-lópez, J. Protection of Pepper Plants from Drought by Microbacterium sp. 3J1 by Modulation of the Plant’s Glutamine and α -ketoglutarate Content: A Comparative Metabolomics Approach. Front. Microbiol. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Kumaresan, V.; Nizam, F.; Ravichandran, G.; Viswanathan, K.; Palanisamy, R.; Bhatt, P.; Arasu, M.V.; Al-Dhabi, N.A.; Mala, K.; Arockiaraj, J. Transcriptome changes of blue-green algae, Arthrospira sp. in response to sulfate stress. Algal Res. 2017, 23, 96–103. [Google Scholar] [CrossRef]
- Smith, C.L.; Steele, J.E.; Stauber, J.L.; Jolley, D.F. Copper-induced changes in intracellular thiols in two marine diatoms: Phaeodactylum tricornutum and Ceratoneis closterium. Aquat. Toxicol. 2014, 156, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhang, R.; Xia, M.; Bai, X.; Mou, J.; Zheng, Y.; Wang, M. Effect of aspartic acid and glutamate on metabolism and acid stress resistance of Acetobacter pasteurianus. Microb. Cell Fact. 2017, 16, 1–14. [Google Scholar] [CrossRef]
- Krastel, K.; Senadheera, D.B.; Mair, R.; Downey, J.S.; Goodman, S.D.; Cvitkovitch, D.G. Characterization of a Glutamate Transporter Operon, glnQHMP, in Streptococcus mutans and Its Role in Acid Tolerance. J. Bacteriol. 2010, 192, 984–993. [Google Scholar] [CrossRef]
- Djoko, K.Y.; Phan, M.; Peters, K.M.; Walker, M.J.; Schembri, M.A.; Mcewan, A.G. Interplay between tolerance mechanisms to copper and acid stress in Escherichia coli. Proc. Natl. Acad. Sci. USA 2017, 114, 6818–6823. [Google Scholar] [CrossRef]
- Hamada, A.; Hibino, T.; Nakamura, T.; Takabe, T. Na+/H+ Antiporter from Synechocystis species PCC6803, Homologous to SOS1, Contains an Aspartic Residue and Long C-Terminal Tail Important for the Carrier Activity. Plant Physiol. 2001, 15, 437–446. [Google Scholar] [CrossRef]
- Gralla, J.D.; Vargas, D.R. Potassium glutamate as a transcriptional inhibitor during bacterial osmoregulation. Eur. Mol. Biol. Organ. 2006, 25, 1515–1521. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Siripornadulsil, S.; Traina, S.; Verma, P.S.; Sayre, R.T. Molecular Mechanisms of Proline-Mediated Tolerance to Toxic Heavy Metals in Transgenic Microalgae. Plant Cell 2002, 14, 2837–2847. [Google Scholar] [CrossRef]
- Zhang, L.P.; Mehta, S.K.; Liu, Z.P.; Yang, Z.M. Copper-Induced Proline Synthesis is Associated with Nitric Oxide Generation in Chlamydomonas reinhardtii. Plant Cell Physiol. 2008, 49, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.-T.; Chang, S.-J.; Chou, T.-L. Intracellular proline accumulation in some algae exposed to copper and caddmium. Bot. Bull. Acad. Sin. Taipei 1995, 36, 89–93. [Google Scholar]
- Phinyo, K. Diversity of Microalgae Genus Scenedesmus and Related Genera from Some Freshwater Resources in Thailand and Some Biochemical Properties of Selected Strains. Ph.D. Thesis, Graduate School, Chiang Mai University, Chiang Mai, Thailand, 2018. [Google Scholar]
- Mota, R.; Pereira, S.B.; Meazzini, M.; Fernandes, R.; Santos, A.; Evans, C.A.; Philippisd, R.D.; Wright, P.C.; Tamagnini, P. Effects of heavy metals on Cyanothece sp. CCY 0110 growth, extracellular polymeric substances (EPS) production, ultrastructure and protein profiles. J. Proteom. 2015, 120, 75–94. [Google Scholar] [CrossRef]
- Saijo, Y. Methods for quantitative determination of chlorophyll. Jpn. J. Limonol. 1975, 36, 103–109. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Davey, M.P.; Burrell, M.M.; Woodward, F.I.; Quick, W.P. Population-specific metabolic phenotypes of Arabidopsis lyrata ssp. petraea. New Phytol. 2005, 177, 380–388. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-mcintyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. The Human Serum Metabolome (HUSERMET) Consortium. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.P.; Woodward, F.I.; Quick, W.P. Intraspecfic variation in cold-temperature metabolic phenotypes of Arabidopsis lyrata ssp. petraea. Metabolomics 2009, 5, 138–149. [Google Scholar] [CrossRef]
- Mangiafico, S.S. An R Companion for the Handbook of Biological Statistics, version 1.3.2. 2015. Available online: Rcompanion.org/documents/RCompanionBioStatistics.pdf (accessed on 24 November 2018).
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Percentage of Copper Removal from the Media ± SD (n = 3) | ||||||
|---|---|---|---|---|---|---|
| pH | 4 | 6 | ||||
| Cu (mg/L) | 2 | 20 | 50 | 2 | 20 | 50 |
| 0 h | 0 | 0 | 0 | 0 | 0 | 0 |
| 0.5 h | 20.3 ± 26.6 a | 13.1 ± 1.8 a | 17.7 ± 3.8 a | 57.6 ± 2.8 b | 82.6 ± 5.2 b | 82.5 ± 1.6 b |
| 8 h | 27.6 ± 24.7 a | 15.5 ± 2.2 a | 13.0 ± 1.8 a | 67.2 ± 24.3 b | 82.6 ± 5.2 b | 81.0 ± 1.9 b |
| 16 h | 4.1 ± 5.7 a | 14.0 ± 4.6 a | 14.6 ± 1.7 a | 64.6 ± 1.5 b | 88.9 ± 2.2 d | 77.2 ± 6.2 c |
| 24 h | 10.7 ± 6.1 a | 16.5 ± 4.0 ab | 27.7 ± 9.1 b | 64.2 ± 1.4 c | 92.9 ± 4.0 d | 81.6 ± 2.6 d |
| 72 h | 34.1 ± 19.4 a | 15.2 ± 4.9 a | 16.4 ± 2.9 a | 65.5 ± 0.5 b | 91.3 ± 2.1 c | 77.0 ± 5.0 bc |
| 120 h | 40.6 ± 16.5 b | 18.0 ± 5.6 a | 15.8 ± 1.7 a | 64.6 ± 1.5 c | 83.7 ± 2.7 cd | 85.7 ± 1.2 d |
| 168 h | 39.2 ± 18.9 a | 34.6 ± 0.6 a | 20.3 ± 0.6 a | 93.5 ± 1.2 b | 73.9 ± 1.1 b | 85.9 ± 0.1 b |
| Group of Metabolites | Pathway | pH | Copper | ||||
|---|---|---|---|---|---|---|---|
| Match | Raw P | Impact | Match | Raw P | Impact | ||
| Lipid | Glycerolipid metabolism | 3/13 | 2.86 × 10−5 | 0.08 | - | ||
| Amino acid | Glycine, serine and threonine metabolism | 4/30 | 8.78 × 10−3 | 0.53 | - | ||
| Cyanoamino acid metabolism | 3/11 | 9.88 × 10−3 | 0.00 | - | |||
| Selenoamino acid metabolism | 2/19 | 3.98 × 10−2 | 0.00 | - | |||
| Valine, leucine and isoleucine degradation | 4/34 | 4.02 × 10−2 | 0.00 | - | |||
| Alanine, aspartate and glutamate metabolism | 4/22 | 4.04 × 10−2 | 0.54 | - | |||
| Aminoacyl-tRNA biosynthesis | 10/67 | 1.17 × 10−2 | 0.09 | - | |||
| Nitrogen metabolism | 3/15 | 2.06 × 10−2 | 0.00 | - | |||
| Arginine and proline metabolism | - | 5/38 | 2.15 × 10−6 | 0.27 | |||
| Lysine biosynthesis | - | 1/10 | 1.40 × 10−2 | 0.00 | |||
| beta-Alanine metabolism | - | 1/12 | 1.40 × 10−2 | 0.00 | |||
| Butanoate metabolism | - | 2/18 | 4.01 × 10−2 | 0.00 | |||
| Antioxidant or Homeostasis | Glutathione metabolism | 4/26 | 1.20 × 10−2 | 0.09 | 4/26 | 2.84 × 10−3 | 0.09 |
| Ascorbate and aldarate metabolism | - | 1/15 | 8.24 × 10−3 | 0.00 | |||
| Carbohydrate (Sugar) | Galactose metabolism | 5/26 | 1.65 × 10−2 | 0.09 | 5/26 | 3.03 × 10−3 | 0.09 |
| Inositol phosphate metabolism | - | 1/24 | 8.24 × 10−3 | 0.25 | |||
| Photosynthesis | Carbon fixation | 2/21 | 3.94 × 10−2 | 0.00 | - | ||
| Porphyrin and chlorophyll metabolism | - | 1/29 | 4.32 × 10−3 | 0.00 | |||
| Energy | Pentose phosphate pathway | - | 2/18 | 7.60 × 10−5 | 0.00 | ||
| Glyoxylate and dicarboxylate metabolism | - | 1/17 | 2.78 × 10−3 | 0.00 | |||
| Nicotinate and nicotinamide metabolism | - | 1/12 | 1.40 × 10−2 | 0.00 | |||
| Methane metabolism | 2/11 | 9.98 × 10−3 | 0.17 | - | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buayam, N.; Davey, M.P.; Smith, A.G.; Pumas, C. Effects of Copper and pH on the Growth and Physiology of Desmodesmus sp. AARLG074. Metabolites 2019, 9, 84. https://doi.org/10.3390/metabo9050084
Buayam N, Davey MP, Smith AG, Pumas C. Effects of Copper and pH on the Growth and Physiology of Desmodesmus sp. AARLG074. Metabolites. 2019; 9(5):84. https://doi.org/10.3390/metabo9050084
Chicago/Turabian StyleBuayam, Nattaphorn, Matthew P. Davey, Alison G. Smith, and Chayakorn Pumas. 2019. "Effects of Copper and pH on the Growth and Physiology of Desmodesmus sp. AARLG074" Metabolites 9, no. 5: 84. https://doi.org/10.3390/metabo9050084
APA StyleBuayam, N., Davey, M. P., Smith, A. G., & Pumas, C. (2019). Effects of Copper and pH on the Growth and Physiology of Desmodesmus sp. AARLG074. Metabolites, 9(5), 84. https://doi.org/10.3390/metabo9050084