Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium


Abstract
1. Introduction
2. Materials and Methods
2.1. Endophytic Fungi from Cymbidium Aloifolium
2.2. Screening for Production of Siderophore by Endophytic Fungi
2.3. Determination of Siderophore Concentration by CAS Liquid Assay
2.4. Detection of Siderophore Type
2.5. Optimization of Culture Parameters for Maximum Siderophore Production
2.6. Production and Extraction of Siderophore Produced by Endophytic Fungi
2.7. Estimation of Total Protein Concentration in Crude and Solvent Purified Siderophore Extract
2.8. Agar Well Diffusion of Crude and Solvent Extracted Siderophore on CAS Agar
2.9. Atomic Absorption Spectra (AAS) Analysis of Solvent Extracted Siderophore
2.10. Purification and Characterization of Siderophore
2.11. Liquid Chromatography Election Spray Ionization-Mass Spectrometry
2.12. Antibacterial Activity of Siderophores Against Plant Pathogenic Bacteria
2.13. Statistical Analysis
3. Results
3.1. Screening for Production of Siderophore by Endophytic Fungi
3.2. Determination of Siderophore Concentration by CAS Liquid Assay
3.3. FeCl3, Tetrazolium and Csaky Test
3.4. Optimization of Culture parameters for Maximum Siderophore Production by Endophytic Fungi
3.5. Production and Extraction of Siderophore
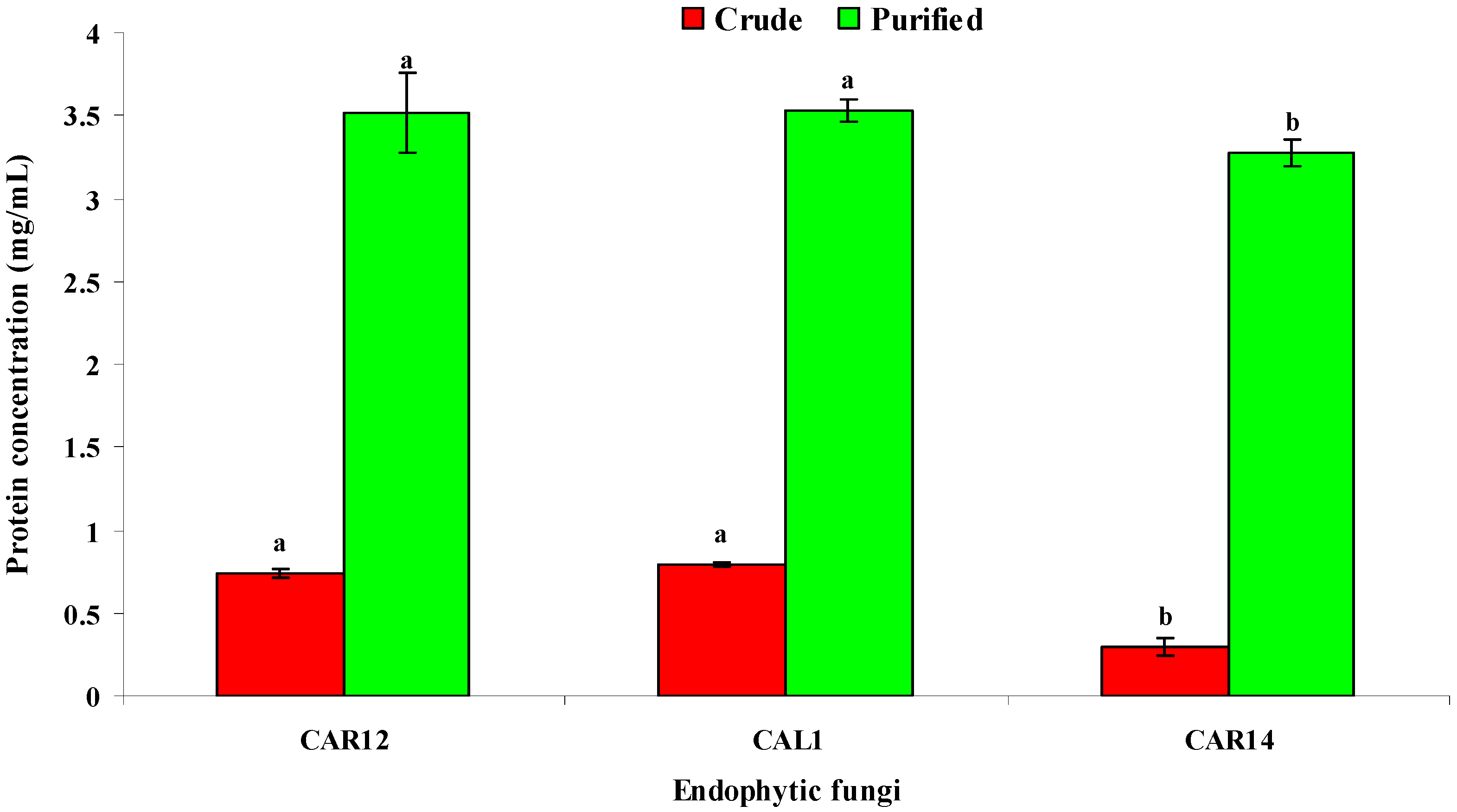
3.6. Estimation of Total Protein Content
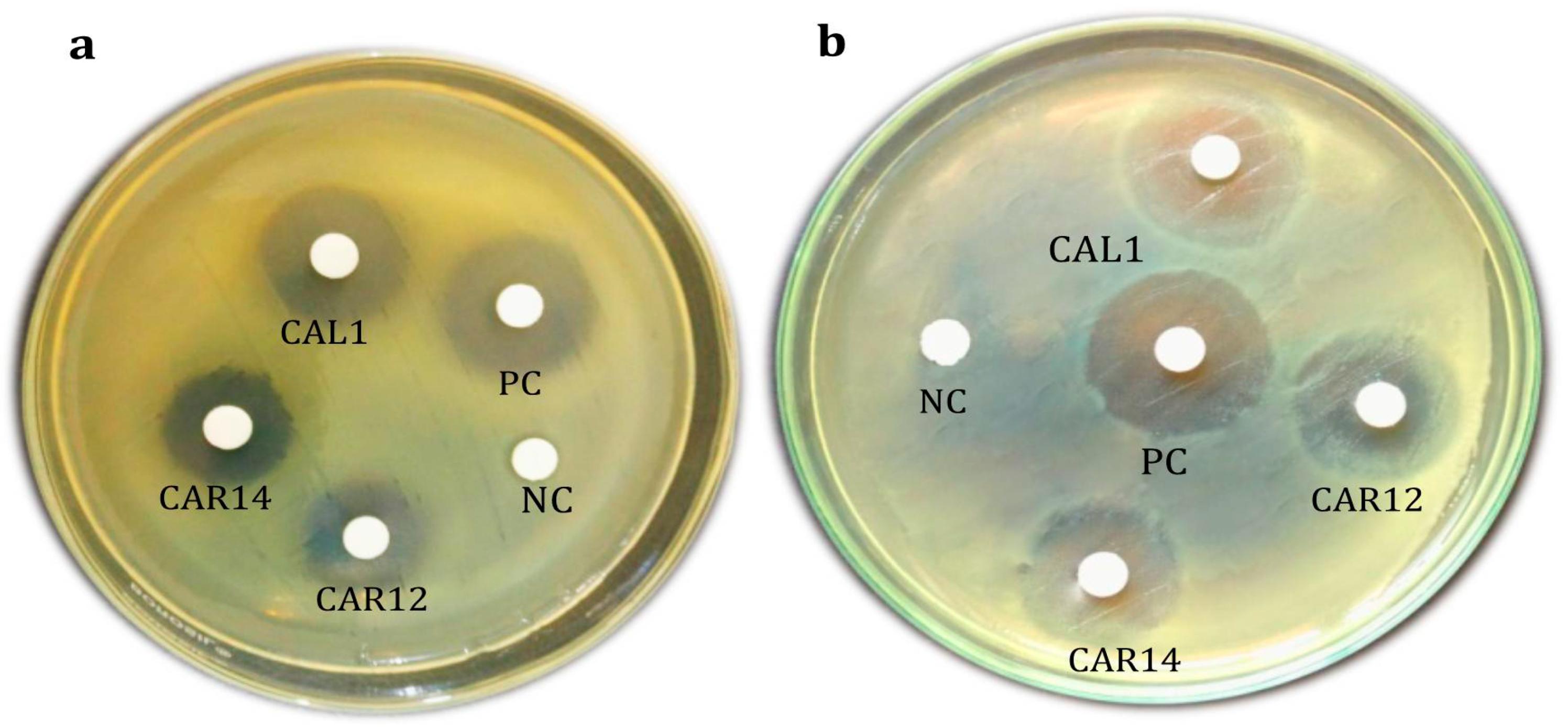
3.7. Agar Well Diffusion of Crude and Solvent Extracted Siderophore on CAS Agar
3.8. Atomic Absorption Spectra (AAS) Analysis
3.9. Purification of Penicillium Chrysogenum (CAL1) Siderophore
3.10. Thin Layer Chromatography of Purified Siderophore
3.11. High-Performance Liquid Chromatography (HPLC) and Liquid Chromatography Electron Spray Ionization Mass Spectrometry (LC-ESI-MS)
3.12. Antibacterial Activity of Fungal Siderophores on Plant Pathogenic Bacteria Ralstonia Solanacearum and Xanthomonas oryzae pv. Oryzae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Praveen, S.; Milan, V.K.; Shivakantkumar, S.A.; Jogaiah, S. Influence of triazole pesticides on tillage soil microbial populations and metabolic changes. Sci. Total Environ. 2019, 651, 2334–2344. [Google Scholar]
- Colombo, C.; Palumbo, G.; He, J.Z.; Pinton, R.; Cesco, S. Review on iron availability in soil: Interaction of Fe minerals, plants, and microbes. J. Soils Sediments 2014, 14, 538–548. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Haas, H. Molecular genetics of fungal siderophore biosynthesis and uptake: The role of siderophores in iron uptake and storage. Appl. Microbiol. Biotechnol. 2003, 62, 316–330. [Google Scholar] [CrossRef]
- Khan, A.; Singh, P.; Srivastava, A. Synthesis, nature and utility of universal iron chelator–Siderophore: A review. Microbiol. Res. 2018, 212, 103–111. [Google Scholar] [CrossRef]
- Sheldon, J.R.; Heinrichs, D.E. Recent developments in understanding the iron acquisition strategies of gram-positive pathogens. FEMS Microbiol. Rev. 2015, 39, 592–630. [Google Scholar] [CrossRef]
- Anupama, N.B.; Jogaiah, S.; Ito, S.; Amruthesh, K.N.; Tran, L.-S.P. Improvement of growth, fruit weight and early blight disease protection of tomato plants by rhizosphere bacteria is correlated with their beneficial traits and induced biosynthesis of antioxidant peroxidase and polyphenol oxidase. Plant Sci. 2015, 231, 62–73. [Google Scholar]
- Jogaiah, S.; Mahantesh, K.; Sharathchnadra, R.G.; Shetty, H.S.; Vedamurthy, A.B.; Tran, L.-S.P. Isolation and evaluation of proteolytic actinomycete isolates as novel inducers of pearl millet downy mildew disease protection. Sci. Rep. 2016, 6, 30789–30802. [Google Scholar] [CrossRef]
- Boukhalfa, H.; Crumbliss, A.L. Chemical aspects of siderophore mediated iron transport. Biometals 2002, 15, 325–339. [Google Scholar] [CrossRef]
- Haselwandter, K.; Winkelmann, G. Siderophores of symbiotic fungi. In Microbial Siderophores; Springer: Berlin/Heidelberg, Germany, 2007; pp. 91–103. [Google Scholar]
- Tripathi, S.; Mishra, S.K.; Varma, A. Mycorrhizal fungi as control agents against plant pathogens. In Mycorrhiza-Nutrient Uptake, Biocontrol, Ecorestoration; Springer: Cham, Switzerland, 2017; pp. 161–178. [Google Scholar]
- Loyevsky, M.; John, C.; Dickens, B.; Hu, V.; Miller, J.H.; Gordeuk, V.R. Chelation of iron within the erythrocytic Plasmodium falciparum parasite by iron chelators. Mol. Biochem. Parasitol. 1999, 101, 43–59. [Google Scholar] [CrossRef]
- Garnerin, T.; Dassonville-Klimpt, A.; Sonnet, P. Fungal Hydroxamate Siderophores: Biosynthesis, chemical synthesis and potential medical applications. In Antimicrobial Research: Novel Bioknowledge and Educational Programs; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2017; pp. 477–488. [Google Scholar]
- Miethke, M.; Marahiel, M.A. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chi, Z. Siderophores from marine microorganisms and their applications. J. Ocean. Univ. China 2004, 3, 40–47. [Google Scholar] [CrossRef]
- Jogaiah, S.; Roopa, K.S.; Pushpalatha, H.G.; Shekar Shetty, H. Evaluation of plant growth-promoting rhizobacteria for their efficiency to promote growth and induce systemic resistance in pearl millet against downy mildew disease. Arch. Phytopath. Plant Prot. 2010, 43, 368–378. [Google Scholar] [CrossRef]
- Bajpai, P. Biological bleaching of chemical pulps. Crit. Rev. Biotechnol. 2004, 24, 1–58. [Google Scholar] [CrossRef]
- Pietro-Souza, W.; de Campos Pereira, F.; Mello, I.S.; Stachack, F.F.F.; Terezo, A.J.; da Cunha, C.N.; White, J.F.; Li, H.; Soares, M.A. Mercury resistance and bioremediation mediated by endophytic fungi. Chemosphere 2020, 240, 124874. [Google Scholar] [CrossRef]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef]
- Wasi, S.; Tabrez, S.; Ahmad, M. Toxicological effects of major environmental pollutants: An overview. Environ. Mon. Ass. 2013, 185, 2585–2593. [Google Scholar] [CrossRef]
- Peralta, D.R.; Adler, C.; Corbalán, N.S.; García, E.C.P.; Pomares, M.F.; Vincent, P.A. Enterobactin as part of the oxidative stress response repertoire. PLoS ONE 2016, 11, 1–15. [Google Scholar] [CrossRef]
- Szebesczyk, A.; Olshvang, E.; Shanzer, A.; Carver, P.L.; Gumienna-Kontecka, E. Harnessing the power of fungal siderophores for the imaging and treatment of human diseases. Coord. Chem. Rev. 2016, 327, 84–109. [Google Scholar] [CrossRef]
- Meyer, J.M.; Stintzi, A.; De Vos, D.; Cornelis, P.; Tappe, R.; Taraz, K.; Budzikiewicz, H. Use of siderophores to type pseudomonads: The three Pseudomonas aeruginosa pyoverdine systems. Microbiology 1997, 143, 35–43. [Google Scholar] [CrossRef]
- Radhika, B.; Murthy, J.V.V.S.N.; Grace, N.D. Preliminary phytochemical analysis & antibacterial activity against clinical pathogens of medicinally important orchid Cymbidium aloifolium (L.) Sw. Int. J. Pharm. Sci. Res. 2013, 4, 3925–3931. [Google Scholar]
- Zhu, G.S.; Yu, Z.N.; Gui, Y.; Liu, Z.Y. A novel technique for isolating orchid mycorrhizal fungi. Fungal. Div. 2008, 33, 123–138. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: St. Paul, MN, USA, 1998; p. 218. [Google Scholar]
- Nagamani, A.; Kunwar, I.K.; Manoharachary, C. Handbook of Soil Fungi; I.K International Publication House, Pvt. Ltd.: New Delhi, India, 2006. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Grimm, P.W.; Allen, P.J. Promotion by zinc of the formation of cytochromes in Ustilago sphaerogena. Plant Physiol. 1954, 29, 369–378. [Google Scholar] [CrossRef]
- Neilands, J.B. Microbial iron compounds. Ann. Rev. Biochem. 1981, 50, 5–31. [Google Scholar] [CrossRef]
- Snow, G.A. Mycobactin. A growth factor for Mycobacterium johnei. Part III. Degradation and tentative structure. J. Chem. Soc. Resumed 1954, 4080–4093. [Google Scholar] [CrossRef]
- Csaky, T.Z. On the estimation of bound hydroxylamine in biological materials. Acta Chem. Scand. 1948, 2, 450–454. [Google Scholar] [CrossRef]
- Jalal, M.A.F.; Van der Helm, D. Isolation and spectroscopic identification of fungal siderophores. In CRC Handbook of Microbial Iron Chelates; CRC Press: Boca Raton, FL, USA, 1991; pp. 235–269. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Karadjova, I.; Izgi, B.; Gucer, S. Fractionation and speciation of Cu, Zn and Fe in wine samples by atomic absorption spectrometry. Spectrochim. Acta Part B At. Spectrosc. 2002, 57, 581–590. [Google Scholar] [CrossRef]
- Murugappan, R.M.; Aravinth, A.; Karthikeyan, M. Chemical and structural characterization of hydroxamate siderophore produced by marine Vibrio harveyi. J. Indian Microbiol. Biotechnol. 2011, 38, 265–273. [Google Scholar] [CrossRef]
- Storey, E.P.; Boghozian, R.; Little, J.L.; Lowman, D.W.; Chakraborty, R. Characterization of ‘Schizokinen’; a dihydroxamate-type siderophore produced by Rhizobium leguminosarum IARI 917. Biometals 2006, 19, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Kajula, M.; Tejesvi, M.V.; Kolehmainen, S.; Mäkinen, A.; Hokkanen, J.; Mattila, S.; Pirttilä, A.M. The siderophore ferricrocin produced by specific foliar endophytic fungi in vitro. Fungal Biol. 2010, 114, 248–254. [Google Scholar] [CrossRef]
- Ayandele, A.A.; Adebiyi, A.O. The phytochemical analysis and antimicrobial screening of extracts of Olax subscorpioidea. Afr. J. Biotechnol. 2007, 6, 868–870. [Google Scholar]
- Neilands, J.B. Siderophores. Arch. Biochem. Biophys. 1993, 302, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Neilands, J.B. Siderophores: Structure and function of microbial iron transport compounds. J. Biol. Chem. 1995, 270, 26723–26726. [Google Scholar] [CrossRef] [PubMed]
- Goodell, B.; Jellison, J.; Liu, J.; Daniel, G.; Paszczynski, A.; Fekete, F.; Krishnamurthy, S.; Jun, L.; Xu, G. Low molecular weight chelators and phenolic compounds isolated from wood decay fungi and their role in the fungal biodegradation of wood. J. Biotechnol. 1997, 53, 133–162. [Google Scholar] [CrossRef]
- Aramsirirujiwet, Y.; Gumlangmak, C.; Kitpreechavanich, V. Studies on antagonistic effect against plant pathogenic fungi from endophytic fungi isolated from Hottuynia Cordata Thunb and screening for Siderophore and indole-3-acetic acid production. Asia Pac. J. Sci. Technol. 2016, 21, 55–66. [Google Scholar]
- Hordt, W.; Römheld, V.; Winkelmann, G. Fusarinines and dimerum acid, mono-and dihydroxamate siderophores from Penicillium chrysogenum, improve iron utilization by strategy I and strategy II plants. Biometals 2000, 13, 37–46. [Google Scholar] [CrossRef]
- Baakza, A.; Vala, A.K.; Dave, B.P.; Dube, H.C. A comparative study of siderophore production by fungi from marine and terrestrial habitats. J. Exp. Mar. Biol. Ecol. 2004, 311, 1–9. [Google Scholar] [CrossRef]
- Calvente, V.; de Orellano, M.E.; Sansone, G.; Benuzzi, D.; de Tosetti, M.I.S. A simple agar plate assay for screening siderophore producer yeasts. J. Microbiol. Met. 2001, 47, 273–279. [Google Scholar] [CrossRef]
- Dave, B.P.; Anshuman, K.; Hajela, P. Siderophores of halophilic Archaea and their chemical characterization. Indian J. Exp. Biol. 2006, 44, 340–344. [Google Scholar] [PubMed]
- Gillam, A.H.; Lewis, A.G.; Andersen, R.J. Quantitative determination of hydroxamic acids. Anal. Chem. 1981, 53, 841–844. [Google Scholar] [CrossRef]
- Volker, C.; Wolf-Gladrow, D.A. Physical limits on iron uptake mediated by siderophores or surface reductases. Mar. Chem. 1999, 65, 227–244. [Google Scholar] [CrossRef]
- Dave, B.P.; Dube, H.C. Chemical characterization of fungal siderophores. Indian J. Exp. Biol. 2000, 38, 56–62. [Google Scholar] [PubMed]
- Roy, N.; Bhattacharyya, P.; Chakrabartty, P.K. Iron acquisition during growth in an iron-deficient medium by Rhizobium sp. isolated from Cicer arietinum. Microbiology 1994, 140, 2811–2820. [Google Scholar] [CrossRef][Green Version]
- Mahmoud, A.L.E.; Abd-Alla, M.H. Siderophore production by some microorganisms and their effect on Bradyrhizobium-Mung Bean symbiosis. Int. J. Agric. Biol. 2001, 3, 157–162. [Google Scholar]
- Sistorm, D.E.; Michlis, L. The effect of D-glucose on utilization of D-mannose and D-fructose by filamentous fungi. J. Bacteriol. 1955, 20, 35–55. [Google Scholar]
- Tailor, A.J.; Joshi, B. Characterization and optimization of siderophore production from Pseudomonas fluorescens strain isolated from sugarcane rhizosphere. J. Environ. Res. Dev. 2012, 6, 688–694. [Google Scholar]
- Aziz, O.A.; Helal, G.A.; Galal, Y.G.M.; Rofaida, A.K.S. Fungal siderophores production in vitro as affected by some abiotic factors. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 210–222. [Google Scholar] [CrossRef]
- Pidacks, C.; Whitehill, A.R.; Pruess, L.M.; Hesseltine, C.W.; Hutchings, B.L.; Bohonos, N.; Williams, J.H. Coprogen, the isolation of a new growth factor required by Pilobolus species. J. Am. Chem. Soc. 1953, 75, 6064–6065. [Google Scholar] [CrossRef]
- Okami, Y.; Kurasawa, S.; Komiyama, T.; Takahashi, A.; Ishizuka, M.; Umezawa, H. Bisucaberin. U.S. Patent 4,870,172 A, 26 September 1989. [Google Scholar]
- Boyer, R. Modern Experimental Biochemistry, 3rd ed.; Addison Wesley Longman: San Francisco, CA, USA, 2000. [Google Scholar]
- Pluhacek, T.; Lemr, K.; Ghosh, D.; Milde, D.; Novak, J.; Havlicek, V. Characterization of microbial siderophores by mass spectrometry. Mass Spectrom. Rev. 2016, 35, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, T.; Clardy, J.A. Chimeric siderophore halts swarming Vibrio. Angew. Chem. Int. Ed. 2014, 53, 3510–3513. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, G.; Schmid, D.G.; Nicholson, G.; Jung, G.; Colquhoun, D.J. Bisucaberin—A dihydroxamate siderophore isolated from Vibrio salmonicida, an important pathogen of farmed Atlantic salmon (Salmo salar). Biometals 2002, 15, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Nakamura, H.; Kameyama, T.; Kurasawa, S.; Naganawa, H.; Okami, Y.; Takeuchi, T.; Umezawa, H.; Iitaka, Y. Bisucaberin, a new siderophore, sensitizing tumor cells to macrophage-mediated cytolysis. J. Antib. 1987, 40, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Dhungana, S.; White, P.S.; Crumbliss, A.L. Crystal structure of ferrioxamine B: A comparative analysis and implications for molecular recognition. J. Biol. Inorg. Chem. 2001, 6, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Ishida, Y. Production of dihydroxamate siderophore alcaligin by Alcaligenes denitrificans sub sp. xylosoxydans. Agric. Biol. Chem. 1990, 54, 1837–1839. [Google Scholar]
- Brickman, T.J.; Hansel, J.G.; Miller, M.J.; Armstrong, S.K. Purification, spectroscopic analysis and biological activity of the macrocyclic dihydroxamate siderophore alcaligin produced by Bordetella pertussis and Bordetella bronchiseptica. Biometals 1996, 9, 191–203. [Google Scholar] [CrossRef]
- Kameyama, T.; Takahashi, A.; Kurasawa, S.; Ishizuka, M.; Okami, Y.; Takeuchi, T.; Umezawa, H. Bisucaberin, A new siderophore, sensitizing tumor cells to macrophage-mediated cytolysis. J. Antibiot. 1987, 40, 1664–1670. [Google Scholar] [CrossRef]
- Saxena, B.; Modi, M.; Modi, V.V. Isolation and characterization of siderophores from Azospirillum lipoferum D-2. Microbiology 1986, 132, 2219–2224. [Google Scholar] [CrossRef]
- Saha, R.; Saha, N.; Donofrio, R.S.; Bestervelt, L.L. Microbial siderophores: A mini review. J. Microbiol. 2012, 52, 1–15. [Google Scholar] [CrossRef]
- Chua, A.C.; Ingram, H.A.; Raymond, K.N.; Baker, E. Multidentate pyridinones inhibit the metabolism of non-transferrin bound iron by hepatocytes and hepatoma cells. Eur. J. Biochem. 2003, 270, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Pseudomonas siderophores: A mechanism explaining disease-suppressive soils. Curr Microbiol. 1980, 4, 317–320. [Google Scholar] [CrossRef]
- Leong, J. Siderophores: Their biochemistry and possible role in the biocontrol of plant pathogens. Ann. Rev. Phytopathol. 1986, 24, 187–209. [Google Scholar] [CrossRef]
- Ahmed, E.; Holmström, S.J. Siderophores in environmental research: Roles and applications. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, H.; Weller, D.M.; Thomashow, L.S. Relative importance of fluorescent siderophores and other factors in biological control of Gaeumannomyces graminis var. tritici by Pseudomonas fluorescens 2-79 and M4-80R. Appl. Environ. Microbiol. 1991, 57, 3270–3277. [Google Scholar]
- McLoughlin, T.J.; Quinn, J.P.; Bettermann, A.; Bookland, R. Pseudomonas cepacia suppression of sunflower wilt fungus and role of antifungal compounds in controlling the disease. Appl. Environ. Microbiol. 1992, 58, 1760–1763. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No | Isolates | Endophytic Fungi | Source | Yellow-Orange Zone on CAS | Mean of Halo Diameter in mm ± SD; n = 3 |
|---|---|---|---|---|---|
| 1 | CAR 1 | Aspergillus japonicus | Root | Positive | 6.33 ± 0.57 b |
| 2 | CAR 2 | Curvularia lunata | Negative | 0 ± 0 a | |
| 3 | CAR 3 | Nigrospora sp. | Negative | 0 ± 0 a | |
| 4 | CAR 4 | C. gloeosporioides | Positive | 9.66 ± 0.57 c | |
| 5 | CAR 5 | Trichoderma sp. | Positive | 7.33 ± 1.15 b | |
| 6 | CAR 6 | Xylaria sp. | Positive | 9.33 ± 2.08 c | |
| 7 | CAR 7 | Rhizoctonia sp. | Negative | 0 ± 0 a | |
| 8 | CAR8 | F. chlamydosporum | Negative | 0 ± 0 a | |
| 9 | CAR 9 | Penicilliumcitrinum | Positive | 9.33 ± 1.52 c | |
| 10 | CAR10 | Helminthosporium sp. | Negative | 0 ± 0 a | |
| 11 | CAR 11 | Curvularia sp. | Positive | 10 ± 1 c | |
| 12 | CAR 12 | Aspergillus sydowii | Positive | 12.33 ± 0.57 e | |
| 13 | CAR 13 | Cladosporium sp. | Positive | 7 ± 1 b | |
| 14 | CAR 14 | Aspergillus terreus | Positive | 12 ± 0 de | |
| 15 | CAR 15 | Alternariaalternata | Positive | 6.66 ± 0.57 b | |
| 16 | CAR 16 | Fusarium oxysporum | Negative | 0 ± 0 a | |
| 17 | CAL1 | P. chrysogenum | Leaf | Positive | 14.33 ± 2.08 f |
| 18 | CAL2 | Aspergillus sydowii | Positive | 7.66 ± 1.52 b | |
| 19 | CAL3 | Trichoderma sp. | Negative | 0 ± 0 a | |
| 20 | CAL4 | Rhizoctonia sp. | Positive | 7.33 ± 1.15 b | |
| 21 | CAL5 | Curvularia lunata | Negative | 0 ± 0 a | |
| 22 | CAL6 | Penicillium citrinum | Negative | 0 ± 0 a | |
| 23 | CAL7 | C.truncatum | Positive | 7 ± 1 b | |
| 24 | CAL8 | Alternaria alternata | Positive | 6.66 ± 0.57 b | |
| 25 | CAL9 | Bipolaris sp. | Negative | 0 ± 0 a | |
| 26 | CAF1 | Fusariumoxysporum | Flower | Positive | 7.66 ± 0.57 b |
| 27 | CAF2 | T. rotundus | Negative | 0 ± 0 a | |
| 28 | CAF3 | P. purpurogenum | Positive | 10.66 ± 1.52 cd | |
| 29 | CAF4 | Cladosporium sp. | Negative | 0 ± 0 a | |
| 30 | CAF5 | Cylindrocephalum sp. | Positive | 7 ± 1 b |
| Sl. No | Fungal Isolate | Endophytic Fungi | % Siderophores ± SD; n = 3 |
|---|---|---|---|
| 1 | CAR1 | Aspergillus japonicus | 27.02 ± 0.45 j |
| 2 | CAR4 | Colletotrichum gloeosporioides | 40.54 ± 0.45 l |
| 3 | CAR5 | Trichoderma sp. | 18.91 ± 0.45 e |
| 4 | CAR6 | Xylaria sp. | 40.99 ± 0.68 l |
| 5 | CAR9 | Penicilliumcitrinum | 25.52 ± 0.68 i |
| 6 | CAR11 | Curvularia sp. | 23.87 ± 0.45 h |
| 7 | CAR12 | Aspergillus sydowii | 48.94 ± 0.68 n |
| 8 | CAR13 | Cladosporium sp. | 35.73 ± 0.68 k |
| 9 | CAR14 | Aspergillus terreus | 46.39 ± 0.45 m |
| 10 | CAR15 | Alternariaalternata | 18.01 ± 0.45 de |
| 11 | CAL1 | Penicillium chrysogenum | 56.6 ± 0.68 o |
| 12 | CAL2 | Aspergillus sydowii | 13.06 ± 0.45 b |
| 13 | CAL4 | Rhizoctonia sp. | 10.35 ± 0.45 a |
| 14 | CAL7 | Colletotrichumtruncatum | 25.22 ± 0.45 i |
| 15 | CAL8 | Alternaria alternata | 21.77 ± 0.68 g |
| 16 | CAF1 | Fusariumoxysporum | 16.06 ± 0.68 c |
| 17 | CAF3 | Penicillium purpurogenum | 17.71 ± 0.68 d |
| 18 | CAF5 | Cylindrocephalum sp. | 20.41 ± 0.69 f |
| Sl. No | Fungal Siderophore | Iron as Fe3+ by AAS (%) | Percentage of Fe3+ with Reference to Control (x) | Percentage of Fe3+ Scavenged by Siderophore: (100 − x) |
|---|---|---|---|---|
| 1 | Control (2% FeCl3) | 0.45 | 100% | - |
| 2 | A. sydowii (CAR12) | 0.13 | 28.88% | 71.12% |
| 3 | P. chrysogenum (CAL1) | 0.076 | 16.88% | 83.12% |
| 4 | A. terreus (CAR14) | 0.12 | 26.66% | 73.34% |
| Sl. No | R. solanacearum Isolate | Zone of Inhibition in mm ± SD; n = 3 | |||
|---|---|---|---|---|---|
| Positive Control | CAR12 | CAR14 | CAL1 | ||
| 1 | APM39 | 9 ± 1 abc | 6.66 ± 0.57 b | 6.33 ± 0.57 b | 8.66 ± 0.57 abc |
| 2 | APM42 | 11.33 ± 1.15 ef | 8.66 ± 0.57 cd | 7.66 ± 0.57 bcd | 9.66 ± 0.57 abcde |
| 3 | APM52 | 19.66 ± 1.52 jk | 9.66 ± 1.52 de | 12.33 ± 1.52 f | 16.33 ± 1.41 hijk |
| 4 | APM53 | 10.33 ± 1.52 bcde | 8.33 ± 0.57 cd | 6.66 ± 1.15 b | 8 ± 1 ab |
| 5 | KAP1 | 10.66 ± 0.57 cde | 0 a | 7.33 ± 0.57 bc | 8.66 ± 0.57 abc |
| 6 | KAP4 | 9.33 ± 0.57 abcd | 0 a | 0 ± 0 a | 7.66 ± 0.57 a |
| 7 | KAP6 | 8.66 ± 0.57 ab | 6.33 ± 0.57 b | 7.33 ± 0.57 bc | 8 ± 1 ab |
| 8 | KAP8 | 12.66 ± 0.57 fg | 8.33 ± 0.57 cd | 7.66 ± 0.57 bcd | 9.33 ± 0.57 abcd |
| 9 | KAP17 | 8.33 ± 0.57 a | 6.66 ± 1.15 b | 7.66 ± 1.15 bcd | 9 ± 1 abcd |
| 10 | KAP18 | 20 ± 1 jk | 10.33 ± 0.57 ef | 11.66 ± 0.57 f | 17 ± 1 ijk |
| 11 | KAP19 | 10.66 ± 0.57 cde | 9.66 ± 0.57 de | 8.33 ± 0.57 cde | 10.66 ± 0.57 abcdef |
| 12 | APH25 | 18.33 ± 0.57 ij | 11.66 ± 0.57 fg | 9.66 ± 0.57 e | 14.66 ± 0.57 ghij |
| 13 | APH26 | 20.33 ± 0.57 k | 14.33 ± 0.57 h | 14.66 ± 0.57 g | 17.33 ± 1.15 jk |
| 14 | APH28 | 12.66 ± 0.57 fg | 9 ± 1 cde | 8.66 ± 0.57 cde | 11.66 ± 0.57 bcdefg |
| 15 | APH36 | 13.66 ± 0.57 gh | 11.33 ± 0.57 fg | 8.66 ± 0.57 cde | 12.66 ± 0.57 defgh |
| 16 | APK9 | 17.66 ± 0.57 i | 14 ± 1 h | 14.33 ± 0.57 g | 16.33 ± 0.57 hijk |
| 17 | APK10 | 13.66 ± 1.15 gh | 11.33 ± 0.57 fg | 12.33 ± 0.57 f | 13.66 ± 0.57 fghij |
| 18 | APA37 | 18.33 ± 0.57 ij | 12.33 ± 0.57 g | 14.66 ± 0.57 g | 16.66 ± 0.57 ijk |
| 19 | APA63 | 11 ± 1 def | 0 a | 8.33 ± 0.57 cde | 8.33 ± 1.15 ab |
| 20 | APP66 | 22.66 ± 1.52 l | 14.33 ± 1.52 h | 12.33 ± 0.57 f | 19.33 ± 0.57 k |
| 21 | APP69 | 13.66 ± 1.15 gh | 9.66 ± 0.57 de | 11 ± 0 f | 13.33 ± 1.52 efghi |
| 22 | APP70 | 12.66 ± 1.52 fg | 0 a | 0 a | 8.66 ± 1.15 abc |
| 23 | APP71 | 15.33 ± 0.57 h | 9.33 ± 0.57 de | 9 ± 1 de | 11.33 ± 0.57 abcdefg |
| 24 | APP73 | 15.33 ± 1.52 h | 7.66 ± 0.57 bc | 9 ± 1 de | 12.33 ± 0.57 cdefg |
| 25 | APP74 | 12.66 ± 0.57 fg | 8.33 ± 1.15 cd | 8.66 ± 1.15 cde | 12.33 ± 0.57 cdefg |
| Sl. No | Xoo Isolate | Zone of Inhibition in mm ± SD; n = 3 | |||
|---|---|---|---|---|---|
| Positive Control | CAR12 | CAR14 | CAL1 | ||
| 1 | MBBT01 | 21.33±1.52 cde | 13.66 ± 1.52 cdef | 16.33 ± 1.52 de | 18.33 ± 0.57 ef |
| 2 | MBBT02 | 23.66 ± 1.52 fg | 18.66 ± 1.52 k | 18.66 ± 1.15 fgh | 21.66 ± 1.52 hi |
| 3 | MBBT03 | 16.33 ± 1.52 ab | 12.33 ± 0.57 c | 13.33 ± 1.15 bc | 14.66 ± 0.57 abc |
| 4 | MBBT04 | 23 ± 1 efg | 15.33 ± 0.57 fgh | 17.33 ± 0.57 ef | 20 ± 1 fgh |
| 5 | MBBT05 | 18 ± 1 b | 12.33 ± 0.57 c | 13.33 ± 0.57 bc | 16.33 ± 0.57 cd |
| 6 | MBBT06 | 16.33 ± 1.52 ab | 9.33 ± 0.57 b | 11.33 ± 0.57 a | 14.33 ± −0.57 ab |
| 7 | MBBT07 | 20.66 ± 0.57 cd | 13.66 ± 1.15 cdef | 14.33 ± 0.57 c | 17.66 ± 1.15 de |
| 8 | MBBT08 | 24 ± 1 g | 18.33 ± 1.52 jk | 19.33 ± 0.57 gh | 21.66 ± 1.52 hi |
| 9 | MBBT09 | 18 ± 1 b | 12.66 ± 0.57 cd | 13.66 ± 0.57 bc | 15.33 ± 0.57 bc |
| 10 | MBBT10 | 23.66 ± 1.52 fg | 16.66 ± 1.15 hij | 20.33 ± 0.57 h | 22 ± 1 i |
| 11 | MBBT11 | 16.33 ± 1.52 ab | 9 ± 1 ab | 12.33 ± 0.57 ab | 14.66 ± 0.57 abc |
| 12 | MBBT12 | 14.66 ± 0.57 a | 7.33 ± 0.57 a | 12.33 ± 0.57 ab | 13.33 ± 0.57 a |
| 13 | MBBT13 | 17.33 ± 0.57 b | 8.66 ± 0.57 ab | 11.33 ± 1.52 a | 15.33 ± 0.57 bc |
| 14 | MBBT14 | 23.66 ± 1.15 fg | 17 ± 1 hijk | 18 ± 1.73 efg | 19.66 ± 0.57 fg |
| 15 | MBBT15 | 24.33 ± 0.57 g | 17.66 ± 2.08 jk | 18.66 ± 1.15 fgh | 23.66 ± 1.15 j |
| 16 | MBBT16 | 22.66 ± 0.57 defg | 16.33 ± 1.52 ghi | 17.66 ± 0.57 efg | 18.66 ± 0.57 ef |
| 17 | MBBT17 | 22.66 ± 1.15 defg | 12.33 ± 0.57 c | 16.33 ± 1.52 de | 20 ± 1 fgh |
| 18 | MBBT18 | 22.66 ± 0.57 defg | 9.66 ± 0.57 b | 14.66 ± 1.52 cd | 18.33 ± 0.57 ef |
| 19 | MBBT19 | 20.66 ± 2.08 cd | 14.66 ± 0.57 efg | 16.33 ± 0.57 de | 17.66 ± 1.52 de |
| 20 | MBBT20 | 20.33 ± 1.52 c | 13.33 ± 0.57 cde | 14.66 ± 0.57 cd | 17.33 ± 1.15 de |
| 21 | MBBT21 | 23.66 ± 0.57 fg | 15.33 ± 0.57 fg | 17.33 ± 057 ef | 20.66 ± 0.57 ghi |
| 22 | MBBT22 | 21.66 ± 0.57 cdef | 12.33 ± 0.57 c | 13.66 ± 0.57 bc | 17.33 ± 1.15 de |
| 23 | MBBT23 | 14.66 ± 0.57 a | 7.33 ± 0.57a | 12.33 ± 0.57ab | 13.33 ± 0.57a |
| 24 | MBBT24 | 21.33 ± 1.52 cde | 14.33 ± 1.52 ef | 16.33 ± 1.52 de | 18.66 ± 0.57 ef |
| 25 | MBBT25 | 23 ± 1 efg | 15.33 ± 0.57 fgh | 17.33 ± 0.57 ef | 20 ± 1 fgh |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chowdappa, S.; Jagannath, S.; Konappa, N.; Udayashankar, A.C.; Jogaiah, S. Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium. Biomolecules 2020, 10, 1412. https://doi.org/10.3390/biom10101412
Chowdappa S, Jagannath S, Konappa N, Udayashankar AC, Jogaiah S. Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium. Biomolecules. 2020; 10(10):1412. https://doi.org/10.3390/biom10101412
Chicago/Turabian StyleChowdappa, Srinivas, Shubha Jagannath, Narasimhamurthy Konappa, Arakere C. Udayashankar, and Sudisha Jogaiah. 2020. "Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium" Biomolecules 10, no. 10: 1412. https://doi.org/10.3390/biom10101412
APA StyleChowdappa, S., Jagannath, S., Konappa, N., Udayashankar, A. C., & Jogaiah, S. (2020). Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium. Biomolecules, 10(10), 1412. https://doi.org/10.3390/biom10101412