Early Growth Response Gene Upregulation in Epstein–Barr Virus (EBV)-Associated Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)

Abstract
:
1. Introduction
2. Early Growth Response (EGR) Gene Upregulation in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)
3. Cellular Immediate Early Gene (IEG) Response
4. Early Growth Response (EGR) Genes
4.1. EGR1
4.2. EGR2 and EGR3
5. Early Growth Response (EGR) Genes in Immunity and Systemic Autoimmunity
6. Early Growth Response (EGR) Genes in Epstein–Barr Virus (EBV) Infection
6.1. EBV Transcription
6.2. Lytic Cycle
6.3. B lymphocyte Transformation
7. Relevance of EGR-Associated Pathology to ME/CFS
8. Conclusions
Funding
Conflicts of Interest
References
- Fukuda, K.; Straus, S.E.; Hickie, I.; Sharpe, M.C.; Dobbins, J.G.; Komaroff, A. The chronic fatigue syndrome: A comprehensive approach to its definition and study. Ann. Intern. Med. 1994, 121, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, B.M.; van de Sande, M.I.; de Meirleir, K.L.; Klimas, N.G.; Broderick, G.; Mitchell, T.; Staines, D.; Powles, A.C.; Speight, N.; Vallings, R.; et al. Myalgic encephalomyelitis: International consensus criteria. J. Intern Med. 2011, 270, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, J.R. Epstein-barr virus induced gene-2 upregulation identifies a particular subtype of chronic fatigue syndrome/myalgic encephalomyelitis. Front. Pediatr. 2019, 3, 59. [Google Scholar] [CrossRef] [PubMed]
- Birkenbach, M.; Josefsen, K.; Yalamanchili, R.; Lenoir, G.; Kieff, E. Epstein-barr virus-induced genes: First lymphocyte-specific G protein-coupled peptide receptors. J. Virol. 1993, 67, 2209–2220. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.R.; Petty, R.; Burke, B.; Gough, J.; Fear, D.; Sinclair, L.I.; Mattey, D.L.; Richards, S.C.; Montgomery, J.; Baldwin, D.A.; et al. Gene expression subtypes in patients with chronic fatigue syndrome/myalgic encephalomyelitis. J. Infect. Dis. 2008, 197, 1171–1184. [Google Scholar] [CrossRef]
- Rutkowska, A.; Dev, K.K.; Sailer, A.W. The role of the Oxysterol/EBI2 pathway in the immune and central nervous systems. Curr. Cancer Drug Targets 2016, 17, 1851–1860. [Google Scholar] [CrossRef]
- Benned-Jensen, T.; Norn, C.; Laurent, S.; Madsen, C.M.; Larsen, H.M.; Arfelt, K.N.; Wolf, R.M.; Frimurer, T.; Sailer, A.W.; Rosenkilde, M.M. Molecular characterization of oxysterol binding to the Epstein-Barr virus-induced gene 2 (GPR183). J. Biol. Chem. 2012, 287, 35470–35483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havis, E.; Duprez, D. EGR1 transcription factor is a multifaceted regulator of matrix production in tendons and other connective tissues. Int. J. Mol. Sci. 2020, 21, 1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herschman, H.R. Primary response genes induced by growth factors and tumor promoters. Annu. Rev. Biochem. 1991, 60, 281–319. [Google Scholar] [CrossRef]
- Brivanlou, A.H.; Darnell, J.E., Jr. Signal transduction and the control of gene expression. Science 2002, 295, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Weake, V.M.; Workman, J.L. Inducible gene expression: Diverse regulatory mechanisms. Nat. Rev. Genet. 2010, 11, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Lau, L.F.; Nathans, D. Expression of a set of growth-related immediate early genes in BALB/c 3T3 cells: Coordinate regulation with c-fos or c-myc. Proc. Natl. Acad. Sci. USA 1987, 84, 1182–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldogh, I.; AbuBakar, S.; Albrecht, T. Activation of proto-oncogenes: An immediate early event in human cytomegalovirus infection. Science 1990, 247, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Angel, P.; Karin, M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim. Biophys. Acta 1991, 1072, 129–157. [Google Scholar] [CrossRef]
- Shaulian, E.; Karin, M. AP-1 as a regulator of cell life and death. Nat. Cell Biol. 2002, 4, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.; Dragunow, M. Induction of immediate-early genes and the control of neurotransmitter-regulated gene expression within the nervous system. Pharmacol. Rev. 1995, 47, 133–178. [Google Scholar] [PubMed]
- Burger, C.; Wick, M.; Brusselbach, S.; Muller, R. Differential induction of ‘metabolic genes’ after mitogen stimulation and during normal cell cycle progression. J. Cell Sci. 1994, 107, 241–252. [Google Scholar] [PubMed]
- Winkles, J.A. Serum- and polypeptide growth factor-inducible gene expression in mouse fibroblasts. Prog. Nucleic Acid Res. Mol. Biol. 1998, 58, 41–78. [Google Scholar]
- Healy, S.; Khan, P.; Davie, J.R. Immediate early response genes and cell transformation. Pharmacol. Ther. 2013, 137, 64–77. [Google Scholar] [CrossRef]
- Levin, W.J.; Press, M.F.; Gaynor, R.B.; Sukhatme, V.P.; Boone, T.C.; Reissmann, P.T.; Figlin, R.A.; Holmes, E.C.; Souza, L.M.; Slamon, D.J. Expression patterns of immediate early transcription factors in human non-small cell lung cancer. The lung cancer study group. Oncogene 1995, 11, 1261–1269. [Google Scholar]
- Dunn, K.L.; Espino, P.S.; Drobic, B.; He, S.; Davie, J.R. The Ras-MAPK signal transduction pathway, cancer and chromatin remodeling. Biochem. Cell. Biol. 2005, 83, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tullai, J.W.; Schaffer, M.E.; Mullenbrock, S.; Shoulder, G.; Kasif, S.; Cooper, G.M. Immediate-early and delayed primary response genes are distinct in function and genomic architecture. J. Biol. Chem. 2007, 282, 23981–23995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, S.; Drablos, F. Gene regulation in the immediate-early response process. Adv. Biol. Regul. 2016, 62, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Sevetson, B.R.; Svaren, J.; Milbrandt, J. A novel activation function for NAB proteins in EGR-dependent transcription of the luteinizing hormone beta gene. J. Biol. Chem. 2000, 275, 9749–9757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, S.B.; Monroe, J.G. The role of early growth response gene 1 (egr-1) in regulation of the immune response. J. Leukoc. Biol. 1996, 60, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Glynne, R.; Ghandour, G.; Rayner, J.; Mack, D.H.; Goodnow, C.C. B-lymphocyte quiescence, tolerance and activation as viewed by global gene expression profiling on microarrays. Immunol. Rev. 2000, 176, 216–246. [Google Scholar]
- Gómez-Martín, D.; Díaz-Zamudio, M.; Galindo-Campos, M.; Alcocer-Varela, J. Early growth response transcription factors and the modulation of immune response: Implications towards autoimmunity. Autoimmun. Rev. 2010, 9, 454–458. [Google Scholar] [CrossRef]
- Taefeshokr, S.; Key, Y.A.; Khakpour, M.; Dadebighlu, P.; Oveisi, A. Early growth response 2 and Egr3 are unique regulators in immune system. Cent. Eur. J. Immunol. 2017, 42, 205–209. [Google Scholar] [CrossRef]
- Schalch, P.; Patejunas, G.; Retuerto, M.; Sarateanu, S.; Milbrandt, J.; Thakker, G.; Kim, D.; Carbray, J.; Crystal, R.G.; Rosengart, T.K. Homozygous deletion of early growth response 1 gene and critical limb ischemia after vascular ligation in mice: Evidence for a central role in vascular homeostasis. J. Thorac. Cardiovasc. Surg. 2004, 128, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Sarateanu, C.S.; Retuerto, M.A.; Beckmann, J.T.; McGregor, L.; Carbray, J.; Patejunas, G.; Nayak, L.; Milbrandt, J.; Rosengart, T.K. An Egr-1 master switch for arteriogenesis: Studies in Egr-1 homozygous negative and wild-type animals. J. Thorac. Cardiovasc. Surg. 2006, 131, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, S.S.; Tocco, G.; Shors, T.J.; Thompson, R.F. Activation of immediate early genes after acute stress. Neuroreport 1991, 2, 17–20. [Google Scholar] [CrossRef]
- Melia, K.R.; Ryabinin, A.E.; Schroeder, R.; Bloom, F.E.; Wilson, M.C. Induction and habituation of immediate early gene expression in rat brain by acute and repeated restraint stress. J. Neurosci. 1994, 14, 5929–5938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Stone, E.; McEwen, B.S. Induction and habituation of c-fos and zif/268 by acute and repeated stressors. Neuroreport 1994, 5, 1321–1324. [Google Scholar] [PubMed]
- Cullinan, W.E.; Herman, J.P.; Battaglia, D.F.; Akil, H.; Watson, S.J. Pattern and time course of immediate early gene expression in rat brain following acute stress. Neuroscience 1995, 64, 477–505. [Google Scholar] [CrossRef]
- Olsson, T.; Hakånsson, A.; Seckl, J.R. Ketanserin selectively blocks acute stress-induced changes in NGFI-A and mineralocorticoid receptor gene expression in hippocampal neurons. Neuroscience 1997, 76, 441–448. [Google Scholar] [CrossRef]
- Knapska, E.; Kaczmarek, L. A gene for neuronal plasticity in the mammalian brain: Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK? Prog. Neurobiol. 2004, 74, 183–211. [Google Scholar] [CrossRef] [PubMed]
- Kozlovsky, N.; Matar, M.A.; Kaplan, Z.; Zohar, J.; Cohen, H. A distinct pattern of intracellular glucocorticoid-related responses is associated with extreme behavioral response to stress in an animal model of post-traumatic stress disorder. Eur. Neuropsychopharmacol. 2009, 19, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Revest, J.-M.; Di Blasi, F.; Kitchener, P.; Rougé-Pont, F.; Desmedt, A.; Turiault, M.; Tronche, F.; Piazza, P.V. The MAPK pathway and Egr-1 mediate stress-related behavioral effects of glucocorticoids. Nat. Neurosci. 2005, 8, 664–672. [Google Scholar] [CrossRef]
- Revest, J.-M.; Kaouane, N.; Mondin, M.; Le Roux, A.; Rougé-Pont, F.; Vallée, M.; Barik, J.; Tronche, F.; Desmedt, A.; Piazza, P.V. The enhancement of stress-related memory by glucocorticoids depends on synapsin-Ia/Ib. Mol. Psychiatry 2010, 1125, 1140–1151. [Google Scholar] [CrossRef] [Green Version]
- Saunderson, E.A.; Spiers, H.; Mifsud, K.R.; Gutierrez-Mecinas, M.; Trollope, A.F.; Shaikh, A.; Mill, J.; Reul, J.M. Stress-induced gene expression and behavior are controlled by DNA methylation and methyl donor availability in the dentate gyrus. Proc. Natl. Acad. Sci. USA 2016, 113, 4830–4835. [Google Scholar] [CrossRef] [Green Version]
- Nelo-Bazán, M.A.; Latorre, P.; Bolado-Carrancio, A.; Pérez-Campo, F.M.; Echenique-Robba, P.; Rodríguez-Rey, J.C.; Carrodeguas, J.A. Early growth response 1 (EGR-1) is a transcriptional regulator of mitochondrial carrier homolog 1 (MTCH 1)/presenilin 1-associated protein (PSAP). Gene 2016, 578, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Irrcher, I.; Hood, D.A. Regulation of Egr-1, SRF, and Sp1 mRNA expression in contracting skeletal muscle cells. J. Appl. Physiol. 2004, 97, 2207–2213. [Google Scholar] [CrossRef] [Green Version]
- Freyssenet, D.; Irrcher, I.; Connor, M.K.; Di Carlo, M.; Hood, D.A. Calcium-regulated changes in mitochondrial phenotype in skeletal muscle cells. Am. J. Physiol. Cell Physiol. 2004, 286, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Covington, H.E., III; Lobo, M.K.; Maze, I.; Vialou, V.; Hyman, J.M.; Zaman, S.; LaPlant, Q.C.; Mouzon, E.; Ghose, S.; Tamminga, C.A.; et al. Antidepressant effect of optogenetic stimulation of the medial prefrontal cortex. J. Neurosci. 2010, 30, 16082–16090. [Google Scholar] [CrossRef]
- Yamada, K.; Gerber, D.J.; Iwayama, Y.; Ohnishi, T.; Ohba, H.; Toyota, T.; Aruga, J.; Minabe, Y.; Tonegawa, S.; Yoshikawa, T. Genetic analysis of the calcineurin pathway identifies members of the EGR gene family, specifically EGR3, as potential susceptibility candidates in schizophrenia. Proc. Natl. Acad. Sci. USA 2007, 104, 2815–2820. [Google Scholar] [CrossRef] [Green Version]
- Kimoto, S.; Bazmi, H.H.; Lewis, D.A. Lower expression of glutamic acid decarboxylase 67 in the prefrontal cortex in schizophrenia: Contribution of altered regulation by Zif268. Am. J. Psychiatry 2014, 171, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Santiago, J.; Diez-Alarcia, R.; Callado, L.F.; Zhang, J.X.; Chana, G.; White, C.H.; Glatt, S.J.; Tsuang, M.T.; Everall, I.P.; Meana, J.J.; et al. A combined analysis of microarray gene expression studies of the human prefrontal cortex identifies genes implicated in schizophrenia. J. Psychiatr. Res. 2012, 46, 1464–1474. [Google Scholar] [CrossRef]
- Northcutt, K.V.; Lonstein, J.S. Social contact elicits immediate early gene expression in dopaminergic cells of the male prairie vole extended olfactory amygdala. Neuroscience 2009, 163, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Ono, K.; Ouchi, H.; Tsushima, R.; Murakami, Y. Social isolation stress down-regulates cortical early growth response 1 (Egr- 1) expression in mice. Neurosci. Res. 2012, 73, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Hodges, T.E.; Green, M.R.; Simone, J.J.; McCormick, C.M. Effects of social context on endocrine function and Zif268 expression in response to an acute stressor in adolescent and adult rats. Int. J. Dev. Neurosci. 2014, 35, 25–34. [Google Scholar] [CrossRef]
- Okada, R.; Fujiwara, H.; Mizuki, D.; Araki, R.; Yabe, T.; Matsumoto, K. Involvement of dopaminergic and cholinergic systems in social isolation-induced deficits in social affiliation and conditional fear memory in mice. Neuroscience 2015, 299, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Okada, R.; Matsumoto, K.; Tsushima, R.; Fujiwara, H.; Tsuneyama, K. Social isolation stress-induced fear memory deficit is mediated by down-regulated neuro-signaling system and Egr-1 expression in the brain. Neurochem. Res. 2014, 39, 875–882. [Google Scholar] [CrossRef]
- Hodges, T.E.; McCormick, C.M. Adolescent and adult male rats habituate to repeated isolation, but only adolescents sensitize to partner unfamiliarity. Horm. Behav. 2015, 69, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Ieraci, A.; Mallei, A.; Popoli, M. Social isolation stress induces anxiousdepressive-like behavior and alterations of neuroplasticity-related genes in adult male mice. Neural Plast. 2016, 2016, 6212983. [Google Scholar] [CrossRef] [Green Version]
- El Rawas, R.; Thiriet, N.; Lardeux, V.; Jaber, M.; Solinas, M. Environmental enrichment decreases the rewarding but not the activating effects of heroin. Psychopharmacology 2009, 203, 561–570. [Google Scholar] [CrossRef]
- Hamlin, A.S.; McNally, G.P.; Osborne, P.B. Induction of c-Fos and zif268 in the nociceptive amygdala parallel abstinence hyperalgesia in rats briefly exposed to morphine. Neuropharmacology 2007, 53, 330–343. [Google Scholar] [CrossRef]
- Ziółkowska, B.; Korostynski, M.; Piechota, M.; Kubik, J.; Przewłocki, R. Effects of morphine on immediate-early gene expression in the striatum of C57BL/6J and DBA/2J mice. Pharmacol. Rep. 2012, 64, 1091–1104. [Google Scholar] [CrossRef]
- Ziółkowska, B.; Gieryk, A.; Solecki, W.; Przewłocki, R. Temporal and anatomic patterns of immediate-early gene expression in the forebrain of C57BL/6 and DBA/2 mice after morphine administration. Neuroscience 2015, 284, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Thiriet, N.; Aunis, D.; Zwiller, J. C-fos and egr-1 immediate-early gene induction by cocaine and cocaethylene in rat brain: A comparative study. Ann. N. Y. Acad. Sci. 2000, 914, 46–57. [Google Scholar] [CrossRef]
- Faria, R.R.; Rueda, A.V.L.; Sayuri, C.; Soares, S.L.; Malta, M.B.; Carrara-Nascimento, P.F.; Alves, A.D.S.; Marcourakis, T.; Yonamine, M.; Scavone, C.; et al. Environmental modulation of ethanol-induced locomotor activity: Correlation with neuronal activity in distinct brain regions of adolescent and adult Swiss mice. Brain Res. 2008, 1239, 127–140. [Google Scholar] [CrossRef]
- Hansson, A.C.; Rimondini, R.; Neznanova, O.; Sommer, W.H.; Heilig, M. Neuroplasticity in brain reward circuitry following a history of ethanol dependence. Eur. J. Neurosci. 2008, 27, 1912–1922. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, J.; Guitart-Masip, M.; Hassankhali, H.; Landgren, S.; Nicoleau, C.; Giménez-Llort, L.; Terenius, L.; Ögren, S.O.; Johansson, B. Effects of naltrexone and acamprosate on alcohol-induced NGFI-A expression in mouse brain. Neurochem. Res. 2008, 33, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Crews, F.T. Adolescent intermittent ethanol exposure enhances ethanol activation of the nucleus accumbens while blunting the prefrontal cortex responses in adult rat. Neuroscience 2015, 293, 92–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.T.; Liu, M.R.; Pei, D.S. Friend or foe, the role of EGR-1 in cancer. Med. Oncol. 2019, 37, 7. [Google Scholar] [CrossRef]
- Liu, J.; Mao, R.; Ren, G.; Liu, X.; Zhang, Y.; Wang, J.; Wang, Y.; Li, M.; Qiu, Q.; Wang, L.; et al. Whole exome sequencing identifies putative predictors of recurrent prostate cancer with high accuracy. OMICS 2019, 23, 380–388. [Google Scholar] [CrossRef]
- Duclot, F.; Kabbaj, M. The role of early growth response 1 (EGR1) in brain plasticity and neuropsychiatric disorders. Front. Behav. Neurosci. 2017, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Riffo-Campos, Á.L.; Castillo, J.; Tur, G.; González-Figueroa, P.; Georgieva, E.I.; Rodríguez, J.L.; López-Rodas, G.; Rodrigo, M.I.; Franco, L. Nucleosome-specific, time- dependent changes in histone modifications during activation of the early growth response 1 (Egr1) gene. J. Biol. Chem. 2015, 290, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Tur, G.; Georgieva, E.I.; Gagete, A.; López-Rodas, G.; Rodríguez, J.L.; Franco, L. Factor binding and chromatin modification in the promoter of murine Egr1 gene upon induction. Cell. Mol. Life Sci. 2010, 67, 4065–4077. [Google Scholar] [CrossRef]
- Gräff, J.; Rei, D.; Guan, J.S.; Wang, W.Y.; Seo, J.; Hennig, K.M.; Nieland, T.J.; Fass, D.M.; Kao, P.F.; Kahn, M.; et al. An epigenetic blockade of cognitive functions in the neurodegenerating brain. Nature 2012, 483, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Korkmaz, K.S.; Braun, K.; Bock, J. Early life stress-induced histone acetylations correlate with activation of the synaptic plasticity genes Arc and Egr1 in the mouse hippocampus. J. Neurochem. 2013, 125, 457–464. [Google Scholar] [CrossRef]
- Rusconi, F.; Grillo, B.; Ponzoni, L.; Bassani, S.; Toffolo, E.; Paganini, L.; Mallei, A.; Braida, D.; Passafaro, M.; Popoli, M.; et al. LSD1 modulates stress- evoked transcription of immediate early genes and emotional behavior. Proc. Natl. Acad. Sci. USA 2016, 113, 3651–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penner, M.R.; Parrish, R.R.; Hoang, L.T.; Roth, T.L.; Lubin, F.D.; Barnes, C.A. Age-related changes in Egr1 transcription and DNA methylation within the hippocampus. Hippocampus 2016, 26, 1008–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massart, R.; Freyburger, M.; Suderman, M.; Paquet, J.; El Helou, J.; Belanger-Nelson, E.; Rachalski, A.; Koumar, O.C.; Carrier, J.; Szyf, M.; et al. The genome-wide landscape of DNA methylation and hydroxymethylation in response to sleep deprivation impacts on synaptic plasticity genes. Transl. Psychiatry 2014, 4, 347. [Google Scholar] [CrossRef] [Green Version]
- Stack, A.; Carrier, N.; Dietz, D.; Hollis, F.; Sorenson, J.; Kabbaj, M. Sex differences in social interaction in rats: Role of the immediate-early gene zif268. Neuropsychopharmacology 2010, 35, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Yagi, S.; Chow, C.; Lieblich, S.E.; Galea, L.A.M. Sex and strategy use matters for pattern separation, adult neurogenesis and immediate early gene expression in the hippocampus. Hippocampus 2016, 26, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Becker, K.A.; Hagen, M.J.; Yan, H.; Roberts, A.L.; Mathews, L.A.; Schneider, S.S.; Siegelmann, H.T.; Macbeth, K.J.; Tirrell, S.M.; et al. Transcriptional responses to estrogen and progesterone in mammary gland identify networks regulating p53 activity. Endocrinology 2008, 149, 4809–4820. [Google Scholar] [CrossRef] [Green Version]
- Slade, J.P.; Carter, D.A. Cyclical expression of egr-1/NGFI-A in the rat anterior pituitary: A molecular signal for ovulation? J. Neuroendocrinol. 2000, 12, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Lee, W.-R.; Safe, S. Egr-1 is activated by 17b-estradiol in MCF-7 cells by mitogen-activated protein kinase-dependent phosphorylation of ELK-1. J. Cell. Biochem. 2004, 93, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Christy, B.A.; Lau, L.F.; Nathans, D. A gene activated in mouse 3T3 cells by serum growth factors encodes a protein with “zinc finger” sequences. Proc. Natl. Acad. Sci. USA 1988, 85, 7857–7861. [Google Scholar] [CrossRef] [Green Version]
- Pavletich, N.P.; Pabo, C.O. Zinc finger-DNA recognition: Crystal structure of a Zif268-DNA complex at 2.1 A. Science 1991, 252, 809–817. [Google Scholar] [CrossRef]
- Swirnoff, A.H.; Milbrandt, J. DNA-binding specificity of NGFI-A and related zinc finger transcription factors. Mol. Cell. Biol. 1995, 15, 2275–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levkovitz, Y.; Baraban, J.M. A dominant negative Egr inhibitor blocks nerve growth factor-induced neurite outgrowth by suppressing c-Jun activation: Role of an Egr/c-Jun complex. J. Neurosci. 2002, 22, 3845–3854. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lin, M.; Abidi, P.; Thiel, G.; Liu, J. Specific interaction of Egr1 and c/EBPb leads to the transcriptional activation of the human low density lipoprotein receptor gene. J. Biol. Chem. 2003, 278, 44246–44254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheval, H.; Chagneau, C.; Levasseur, G.; Veyrac, A.; Faucon-Biguet, N.; Laroche, S.; Davis, S. Distinctive features of Egr transcription factor regulation and DNA binding activity in CA1 of the hippocampus in synaptic plasticity and consolidation and reconsolidation of fear memory. Hippocampus 2012, 22, 631–642. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, D.; Shi, X.; Tang, C.; Xie, X.; Tu, J.; Ge, Q.; Lu, Z. Global Egr1-miRNAs binding analysis in PMA-induced K562 cells using ChIP-Seq. J. Biomed. Biotechnol. 2010, 2010, 867517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zang, J.; Chen, X.; Yang, G.; Zhu, Y.; Wu, Y.; Li, T. Role of miR-124 and miR-141 in the regulation of vascular reactivity and the relationship to RhoA and Rac1 after hemorrhage and hypoxia. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, 206–216. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Warner, L.E.; Mancias, P.; Butler, I.J.; McDonald, C.M.; Keppen, L.; Koob, K.G.; Lupski, J.R. Mutations in the early growth response 2 (EGR2) gene are associated with hereditary myelinopathies. Nat. Genet. 1998, 18, 382–384. [Google Scholar] [CrossRef]
- O’donovan, K.J.; Tourtellotte, W.G.; Milbrandt, J.; Baraban, J.M. The Egr family of transcription-regulatory factors: Progress at the interface of molecular and systems neuroscience. Trends Neurosci. 1999, 22, 167–173. [Google Scholar] [CrossRef]
- Li, S.; Symonds, A.L.; Zhu, B.; Liu, M.; Raymond, M.V.; Miao, T.; Wang, P. Early growth response gene-2 (Egr-2) regulates the development of B and T cells. PLoS ONE 2011, 6, e18498. [Google Scholar] [CrossRef] [Green Version]
- Germain, R.N. T-cell development and the CD4-CD8 lineage decision. Nat. Rev. Immunol. 2002, 2, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Miao, T.; Sebastian, M.; Bhullar, P.; Ghaffari, E.; Liu, M.; Symonds, A.L.; Wang, P. The transcription factors Egr2 and Egr3 are essential for the control of inflammation and antigen-induced proliferation of B and T cells. Immunity 2012, 37, 685–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unoki, M.; Nakamura, Y. Growth-sppressive effects of BPOZ and EGR2, two genes involved in hte PTEN signalling pathway. Oncogene 2001, 20, 4457–4465. [Google Scholar]
- Unoki, M.; Nakamura, Y. EGR2 induces apoptosis in various cancer cell lines by direct transactivation of BNIP3L and BAK. Oncogene 2003, 22, 2172–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salotti, J.; Sakchaisri, K.; Tourtellotte, W.G.; Johnson, P.F. An Arf-Egr-C/EBP beta pathway linked to ras-induced senescence and cancer. Mol. Cell. Biol. 2015, 35, 866–883. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, D.B.; Bhaskaran, A.; Droggiti, A.; Dickinson, S.; D’Antonio, M.; Mirsky, R.; Jessen, K.R. Krox-20 inhibits Jun-NH2-terminal kinase/c-Jun to control Schwann cell proliferation and death. J. Cell. Biol. 2004, 164, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, T.G.; Bernard, V.; Gilardi-Hebenstreit, P.; Raynal, V.; Surdez, D.; Aynaud, M.-M.; Mirabeau, O.; Cidre-Aranaz, F.; Tirode, F.; Zaidi, S.; et al. Chimeric EWSR1-FLI1 regulates the Ewing sarcoma susceptibility gene EGR2 via a GGAA microsatellite. Nat. Genet. 2015, 47, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, R.H. T cell Anergy. Annu. Rev. Immunol. 2003, 21, 305–334. [Google Scholar] [CrossRef]
- Hargreaves, R.G.; Borthwick, N.J.; Montani, M.S.G.; Piccolella, E.; Carmichael, P.; Lechler, R.I.; Akbar, A.N.; Lombardi, G. Dissociation of T cell anergy from apoptosis by blockade of Fas/Apo-1 (CD95) signaling. J. Immunol. 1997, 158, 3099–3107. [Google Scholar]
- Zheng, Y.; Zha, Y.; Driessens, G.; Locke, F.; Gajewski, T.F. Transcriptional regulator early growth response gene 2 (Egr2) is required for T cell anergy in vitro and in vivo. J. Exp. Med. 2012, 209, 2157–2163. [Google Scholar] [CrossRef]
- Zheng, Y.; Zha, Y.; Spaapen, R.M.; Mathew, R.; Barr, K.; Bendelac, A.; Gajewski, T.F. Egr2-dependent gene expression profiling and ChIP-Seq reveal novel biologic targets in T cell anergy. Mol. Immunol. 2013, 55, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.; Lutz, M.A.; Zarek, P.E.; Anders, R.A.; Kersh, G.J.; Powell, J.D. Opposing regulation of T cell function by Egr-1/NAB2 and Egr-2/Egr-3. Eur. J. Immunol. 2008, 38, 528–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.; Wolfraim, L.A.; Drake, C.G.; Horton, M.R.; Powell, J.D. Cutting Edge: TCR-Induced NAB2 enhances T cell function by coactivating IL-2 transcription. J. Immunol. 2006, 177, 8301–8305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safford, M.; Collins, S.; Lutz, M.A.; Allen, A.; Huang, C.-T.; Kowalski, J.; Blackford, A.; Horton, M.R.; Drake, C.; Schwartz, R.H.; et al. Egr-2 and Egr-3 are negative regulators of T cell activation. Nat. Immunol. 2005, 6, 472–480. [Google Scholar] [CrossRef]
- Harris, J.E.; Bishop, K.D.; Phillips, N.E.; Mordes, J.P.; Greiner, D.L.; Rossini, A.A.; Czech, M.P. Early growth response gene-2, a zinc-finger transcription factor, is required for full induction of clonal anergy in CD4+ T cells. J. Immunol. 2004, 173, 7331–7338. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.M.; Park, H.B.; Shin, J.H.; Lee, J.E.; Park, H.Y.; Kho, D.H.; Lee, J.S.; Choi, H.; Okuda, T.; Kokame, K.; et al. Ndrg1 is a T-cell clonal anergy factor negatively regulated by CD28 costimulation and interleukin-2. Nat. Commun. 2015, 6, 8698. [Google Scholar] [CrossRef] [Green Version]
- Topilko, P.; Schneider-Maunoury, S.; Levi, G.; Evercooren, A.B.-V.; Chennoufi, A.B.Y.; Seitanidou, T.; Babinet, C.; Charnay, P. Krox-20 controls myelination in the peripheral nervous system. Nature 1994, 371, 7969. [Google Scholar] [CrossRef]
- Tourtellotte, W.G.; Milbrandt, J. Sensory ataxia and muscle spindle agenesis in mice lacking the transcription factor Egr3. Nat. Genet. 1998, 20, 8791. [Google Scholar] [CrossRef]
- Zhu, B.; Symonds, A.L.; Martin, J.E.; Kioussis, D.; Wraith, D.C.; Li, S.; Wang, P. Early growth response gene 2 (Egr-2) controls the self-tolerance of T cells and prevents the development of lupuslike autoimmune disease. J. Exp. Med. 2008, 205, 2295307. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Ichiyama, K.; Hashimoto, M.; Yoshida, H.; Takimoto, T.; Takaesu, G.; Torisu, T.; Hanada, T.; Yasukawa, H.; Fukuyama, S.; et al. Loss of suppressor of cytokine signaling 1 in helper T cells leads to defective Th17 differentiation by enhancing antagonistic effects of IFN-g on STAT3 and Smads. J. Immunol. 2008, 180, 374656. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Laurence, A.; Kanno, Y.; Pacher-Zavisin, M.; Zhu, B.-M.; Tato, C.; Yoshimura, A.; Hennighausen, L.; O’Shea, J.J. Selective regulatory function of Socs3 in the formation of IL-17-secreting T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 813742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schraml, B.U.; Hildner, K.K.; Ise, W.; Lee, W.-L.; Smith, B.J.; Solomon, B.; Sahota, G.; Sim, J.; Mukasa, R.; Cemerski, S.; et al. The AP-1 transcription factor Batf controls TH17 differentiation. Nature 2009, 460, 4059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yosef, N.; Shalek, A.K.; Gaublomme, J.T.; Jin, H.; Lee, Y.; Awasthi, A.; Wu, C.; Karwacz, K.; Xiao, S.; Jorgolli, M.; et al. Dynamic regulatory network controlling TH17 cell differentiation. Nature 2013, 496, 4618. [Google Scholar] [CrossRef] [PubMed]
- Sumitomo, S.; Fujio, K.; Okamura, T.; Yamamoto, K. Egr2 and Egr3 are the unique regulators for systemic autoimmunity. JAKSTAT 2013, 2, e23952. [Google Scholar] [CrossRef] [Green Version]
- Sumitomo, S.; Fujio, K.; Okamura, T.; Morita, K.; Ishigaki, K.; Suzukawa, K.; Kanaya, K.; Kondo, K.; Yamasoba, T.; Furukawa, A.; et al. Transcription factor early growth response 3 is associated with the TGF-b1 expression and the regulatory activity of CD4-positive T cells in vivo. J. Immunol. 2013, 191, 23519. [Google Scholar] [CrossRef] [Green Version]
- Kehrl, J.H.; Thevenin, C.; Rieckmann, P.; Fauci, A.S. Transforming growth factor-b suppresses human B lymphocyte Ig production by inhibiting synthesis and the switch from the membrane form to the secreted form of Ig mRNA. J. Immunol. 1991, 146, 401623. [Google Scholar]
- Letterio, J.J.; Geiser, A.G.; Kulkarni, A.B.; Dang, H.; Kong, L.; Nakabayashi, T.; Mackall, C.L.; Gress, R.E.; Roberts, A.B. Autoimmunity associated with TGF-b1-deficiency in mice is dependent on MHC class II antigen expression. J. Clin. Investig. 1996, 98, 210919. [Google Scholar] [CrossRef] [Green Version]
- Becker-Merok, A.; Eilertsen, G.O.; Nossent, J.C. Levels of transforming growth factor-b are low in systemic lupus erythematosus patients with active disease. J. Rheumatol. 2010, 37, 203945. [Google Scholar] [CrossRef]
- Ohtsuka, K.; Gray, J.D.; Stimmler, M.M.; Toro, B.; Horwitz, D.A. Decreased production of TGF-b by lymphocytes from patients with systemic lupus erythematosus. J. Immunol. 1998, 160, 253945. [Google Scholar]
- Morita, K.; Okamura, T.; Sumitomo, S.; Iwasaki, Y.; Fujio, K.; Yamamoto, K. Emerging roles of Egr2 and Egr3 in the control of systemic autoimmunity. Rheumatology 2016, 55, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.R.; Li, L.; Yan, M.; Bhaskarabhatla, M.; Mobley, A.B.; Nguyen, C.; Mooney, J.M.; Schatzle, J.D.; Wakeland, E.K.; Mohan, C. Regulation of B cell tolerance by the lupus susceptibility gene Ly108. Science 2006, 312, 16659. [Google Scholar] [CrossRef] [Green Version]
- Myouzen, K.; Kochi, Y.; Shimane, K.; Fujio, K.; Okamura, T.; Okada, Y.; Suzuki, A.; Atsumi, T.; Ito, S.; Takada, K.; et al. Regulatory polymorphisms in EGR2 are associated with susceptibility to systemic lupus erythematosus. Hum. Mol. Genet. 2010, 19, 231320. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, T.; Zetterberg, H.; Wang, Y.C.; Rymo, L. Promoter-Proximal Regulatory Elements Involved in oriP-EBNA1-Independent and -Dependent Activation of the Epstein-Barr Virus C Promoter in B-Lymphoid Cell Lines. J. Virol. 2001, 75, 5796–5811. [Google Scholar] [CrossRef] [Green Version]
- Zalani, S.; Holley-Guthrie, E.; Kenney, S. The Zif268 cellular transcription factor activates expression of the Epstein-Barr virus immediate-early BRLF1 promoter. J. Virol. 1995, 69, 3816–3823. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Lee, H.H.; Chen, Y.T.; Lu, J.; Wu, S.Y.; Chen, C.W.; Takada, K.; Tsai, C.H. Induction of the early growth response 1 gene by Epstein-Barr virus lytic transactivator Zta. J. Virol. 2006, 80, 7748–7755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heather, J.; Flower, K.; Isaac, S.; Sinclair, A.J. The Epstein-Barr virus lytic cycle activator Zta interacts with methylated ZRE in the promoter of host target gene egr1. J. Gen. Virol. 2009, 90, 1450–1454. [Google Scholar] [CrossRef]
- Vockerodt, M.; Wei, W.; Nagy, E.; Prouzova, Z.; Schrader, A.; Kube, D.; Rowe, M.; Woodman, C.B.; Murray, P.G. Suppression of the LMP2A target gene, EGR-1, protects Hodgkin’s lymphoma cells from entry to the EBV lytic cycle. J. Pathol. 2013, 230, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Gradoville, L.; Miller, G. Cellular immediate-early gene expression occurs kinetically upstream of Epstein-Barr virus bzlf1 and brlf1 following cross-linking of the B cell antigen receptor in the Akata Burkitt lymphoma cell line. J. Virol. 2010, 84, 12405–12418. [Google Scholar] [CrossRef] [Green Version]
- Calogero, A.; Cuomo, L.; D’Onofrio, M.; de Grazia, U.; Spinsanti, P.; Mercola, D.; Faggioni, A.; Frati, L.; Adamson, E.D.; Ragona, G. Expression of Egr-1 correlates with the transformed phenotype and the type of viral latency in EBV genome positive lymphoid cell lines. Oncogene 1996, 13, 2105–2112. [Google Scholar]
- Kim, J.H.; Kim, W.S.; Kang, J.H.; Lim, H.Y.; Ko, Y.H.; Park, C. Egr-1, a new downstream molecule of Epstein-Barr virus latent membrane protein 1. FEBS Lett. 2007, 581, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.Y.; Wu, S.J.; Huang, M.H.; Lo, F.Y.; Tsai, M.H.; Tsai, C.H.; Hsu, S.M.; Lin, C.W. EBV-positive Hodgkin lymphoma is associated with suppression of p21cip1/waf1 and a worse prognosis. Mol. Cancer 2010, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Kim, W.S.; Park, C. Epstein-Barr virus latent membrane protein 1 increases genomic instability through Egr-1-mediated up-regulation of activation-induced cytidine deaminase in B-cell lymphoma. Leuk. Lymphoma 2013, 54, 2035–2040. [Google Scholar] [CrossRef] [PubMed]
- Thorley-Lawson, D.A.; Miyashita, E.M.; Khan, G. Epstein-Barr virus and the B cell: That’s all it takes. Trends Microbiol. 1996, 4, 204–208. [Google Scholar] [CrossRef]
- Chaganti, S.; Ma, C.S.; Bell, A.I.; Croom-Carter, D.; Hislop, A.D.; Tangye, S.G.; Rickinson, A.B. Epstein–Barr virus persistence in the absence of conventional memory B cells: IgM+IgD+CD27+ B cells harbor the virus in X-linked lymphoproliferative disease patients. Blood 2008, 112, 672–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conacher, M.; Callard, R.; McAulay, K.; Chapel, H.; Webster, D.; Kumararatne, D.; Chandra, A.; Spickett, G.; Hopwood, P.A.; Crawford, D.H. Epstein-Barr virus can establish infection in the absence of a classical memory B-cell population. J. Virol. 2005, 79, 11128–11134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G. The switch between EBV latency and replication. Yale J. Biol. Med. 1989, 62, 205–213. [Google Scholar]
- Feederle, R.; Kost, M.; Baumann, M.; Janz, A.; Drouet, E.; Hammerschmidt, W.; Delecluse, H.J. The Epstein-Barr virus lytic program is controlled by the co-operative functions of two transactivators. EMBO J. 2000, 19, 3080–3089. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; El-Guindy, A.; Countryman, J.; Ye, J.; Gradoville, L. Lytic cycle switches of oncogenic human gammaherpesviruses. Adv. Cancer Res. 2007, 97, 81–109. [Google Scholar]
- Sinclair, A.J. bZIP proteins of human gammaherpesviruses. J. Gen. Virol. 2003, 84, 1941–1949. [Google Scholar] [CrossRef]
- Sinclair, A.J. Unexpected structure of Epstein-Barr virus lytic cycle activator Zta. Trends Microbiol. 2006, 14, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, A.J.; Farrell, P.J. Epstein-Barr virus transcription factors. Cell Growth Differ. 1992, 3, 557–563. [Google Scholar] [PubMed]
- Speck, S.H.; Chatila, T.; Flemington, E. Reactivation of Epstein-Barr virus: Regulation and function of the BZLF1 gene. Trends Microbiol. 1997, 5, 399–405. [Google Scholar] [CrossRef]
- Gutsch, D.E.; Holley-Guthrie, E.A.; Zhang, Q.; Stein, B.; Blanar, M.A.; Baldwin, A.S.; Kenney, S.C. The bZIP transactivator of Epstein-Barr virus, BZLF1, functionally and physically interacts with the p65 subunit of NF-κB. Mol. Cell. Biol. 1994, 14, 1939–1948. [Google Scholar] [CrossRef] [Green Version]
- Sista, N.D.; Pagano, J.S.; Liao, W.; Kenney, S. Retinoic acid is a negative regulator of the Epstein-Barr virus protein (BZLF1) that mediates disruption of latent infection. Proc. Natl. Acad. Sci. USA 1993, 90, 3894–3898. [Google Scholar] [CrossRef] [Green Version]
- Sista, N.D.; Barry, C.; Sampson, K.; Pagano, J. Physical and functional interaction of the Epstein-Barr virus BZLF1 transactivator with the retinoic acid receptors RARα and RXRα. Nucleic Acids Res. 1995, 23, 1729–1736. [Google Scholar] [CrossRef] [Green Version]
- Swenson, J.J.; Holley-Guthrie, E.; Kenney, S.C. Epstein-Barr virus immediate-early protein BRLF1 interacts with CBP, promoting enhanced BRLF1 transactivation. J. Virol. 2001, 75, 6228–6234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.Y.; Chen, H.; Wang, S.E.; Collete, M.J.a.; Liao, G.; Fujimuro, M.; Farrell, C.J.; Huang, J.; Hayward, S.D.; Hayward, G.S. CCAAT/enhancer binding protein α interacts with ZTA and mediates ZTA-induced p21CIP-1 accumulation and G1 cell cycle arrest during the Epstein-Barr virus lytic cycle. J. Virol. 2003, 77, 1481–1500. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Gutsch, D.; Kenney, S. Functional and physical interaction between p53 and BZLF1: Implications for Epstein-Barr virus latency. Mol. Cell. Biol. 1994, 14, 1929–1938. [Google Scholar] [CrossRef] [Green Version]
- Seyfert, V.L.; McMahon, S.; Glenn, W.; Cao, X.M.; Sukhatme, V.P.; Monroe, J.G. Egr-1 expression in surface Ig-mediated B cell activation. Kinetics and association with protein kinase C activation. J. Immunol. 1990, 145, 3647–3653. [Google Scholar]
- Kieser, A.; Sterz, K.R. The latent membrane protein 1 (LMP1). Curr. Top. Microbiol. Immunol. 2015, 391, 119–149. [Google Scholar]
- Rasa, S.; Nora-Krukle, Z.; Henning, N.; Eliassen, E.; Shikova, E.; Harrer, T.; Scheibenbogen, C.; Murovska, M.; Prusty, B.K. Chronic viral infections in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2018, 16, 268. [Google Scholar] [CrossRef] [Green Version]
- Cliff, J.M.; King, E.C.; Lee, J.S.; Sepúlveda, N.; Wolf, A.S.; Kingdon, C.; Bowman, E.; Dockrell, H.M.; Nacul, L.; Lacerda, E.; et al. Cellular immune function in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). Front. Immunol. 2019, 10, 796. [Google Scholar] [CrossRef] [PubMed]
- Hardcastle, S.L.; Brenu, E.W.; Johnston, S.; Nguyen, T.; Huth, T.; Ramos, S.; Staines, D.; Marshall-Gradisnik, S. Longitudinal analysis of immune abnormalities in varying severities of Chronic Fatigue Syndrome/Myalgic Encephalomyelitis patients. J. Transl. Med. 2015, 13, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blundell, S.; Ray, K.K.; Buckland, M.; White, P.D. Chronic fatigue syndrome and circulating cytokines: A systematic review. Brain Behav. Immun. 2015, 50, 186–195. [Google Scholar] [CrossRef]
- Loebel, M.; Grabowski, P.; Heidecke, H.; Bauer, S.; Hanitsch, L.G.; Wittke, K.; Meisel, C.; Reinke, P.; Volk, H.-D.; Fluge, Ø.; et al. Antibodies to β adrenergic and muscarinic cholinergic receptors in patients with chronic fatigue syndrome. Brain Behav. Immun. 2016, 52, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Scheibenbogen, C.; Loebel, M.; Freitag, H.; Krueger, A.; Bauer, S.; Antelmann, M.; Doehner, W.; Scherbakov, N.; Heidecke, H.; Reinke, P.; et al. Immunoadsorption to remove ß2 adrenergic receptor antibodies in chronic fatigue syndrome CFS/ME. PLoS ONE 2018, 13, e0193672. [Google Scholar] [CrossRef] [Green Version]
- Wirth, K.; Scheibenbogen, C. A unifying hypothesis of the pathophysiology of myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS): Recognitions from the finding of autoantibodies against ß2-adrenergic receptors. Autoimmun. Rev. 2020, 19, 102527. [Google Scholar] [CrossRef]
- Glaser, R.; Pearson, G.R.; Jones, J.F.; Hillhouse, J.; Kennedy, S.; Mao, H.Y.; Kiecolt-Glaser, J.K. Stress-related activation of Epstein-Barr virus. Brain Behav. Immun. 1991, 5, 219–232. [Google Scholar] [CrossRef]
- Cohen, S.; Tyrrell, D.A.; Smith, A.P. Psychological stress and susceptibility to the common cold. N. Engl. J. Med. 1991, 325, 606–612. [Google Scholar] [CrossRef]
- Nguyen, T.; Staines, D.; Johnston, S.; Marshall-Gradisnik, S. Reduced glycolytic reserve in isolated natural killer cells from Myalgic encephalomyelitis/chronic fatigue syndrome patients: A preliminary investigation. Asian Pac. J. Allergy Immunol. 2019, 37, 102–108. [Google Scholar] [PubMed]
- Tomas, C.; Brown, A.; Strassheim, V.; Elson, J.L.; Newton, J.; Manning, P. Cellular bioenergetics is impaired in patients with chronic fatigue syndrome. PLoS ONE 2017, 12, e0186802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandarano, A.H.; Maya, J.; Giloteaux, L.; Peterson, D.L.; Maynard, M.; Gottschalk, C.G.; Hanson, M.R. Myalgic encephalomyelitis/chronic fatigue syndrome patients exhibit altered T cell metabolism and cytokine associations. J. Clin. Investig. 2020, 130, 1491–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| Gene Symbol | Gene Name | GenBank Accession No. | Real-Time PCR Fold-Difference | 2-Tailed P (RT-PCR) |
|---|---|---|---|---|
| EGR1 | Early growth response 1 | NM_001955 | 2.82 | 0.015 † |
| EGR2 | Early growth response 2 | NM_000399 | 2.32 | 0.09 † |
| EGR3 | Early growth response 3 | NM_004421 | 1.92 | 0.017 † |
| SP1 | Sp1 transcription factor | NM_138473 | 2.47 | 0.11 † |
| ETS1 | ETS proto-oncogene 1, transcription factor | NM_005238 | 2.11 | 1 × 10−5 ‡ |
| REPIN1 | Replication initiator 1 | NM_013400 | 3.62 | 6 × 10−6 ‡ |
| GABPA | GA binding protein transcription factor Subunit alpha | NM_002031 | 8.06 | 3 × 10−4 ‡ |
| NHLH1 | Nescient helix loop helix 1 | NM_005589 | 11.51 | 7 × 10−4 ‡ |
| NFKB1 | Nuclear factor kappa B subunit 1 | NM_003998 | 1.59 | 4.04 × 10−5 ‡ |
| Function of EGR Genes in Physiological and Pathological Responses | References |
|---|---|
| Virus Infection | |
| EGR1 is a key mediator in the response to and pathogenesis of many different viruses that infect humans, including those of the herpesviruses, retroviruses, flaviviruses, poxviruses, hepatitis B virus, and Borna disease virus; for Epstein–Barr virus, see Table 3 | See numerous papers in PubMed |
| Immune Response and Inflammation | |
| EGR1 is an important regulator of the immune response and the differentiation pathway of myeloid precursors | [25] |
| Promitotic genes in B lymphocytes include EGR1 and EGR2 | [26] |
| EGR1 functions as a positive regulatory factor in B and T cells via transcription of key cytokines and costimulatory molecules EGR2 and -3 act as negative regulators involved in anergy induction and apoptosis | [27,28] |
| Vascular Homeostasis | |
| EGR1 plays a pivotal role in reperfusion responses to vascular occlusion in mice and possibly other mammals | [29] |
| EGR1 is a critical and potentially therapeutic mediator of revascularization after vascular occlusion and implicates arteriogenesis (collateral vessel formation) as a critical component of this process | [30] |
| Psychological Stress | |
| In animal experiments, acute stress (restraint, immobilization, or forced swim), leads to increased Egr1 mRNA in multiple locations throughout the brain; these include neocortical areas, hippocampus, lateral septum, caudate putamen, nucleus accumbens, amygdala, and paraventricular nucleus (PVN) of the hypothalamus | [31,32,33,34,35,36,37] |
| EGR1 is critical in encoding the chronic behavioural effects of stress, for example, acute exposure to forced swim stress or activation of the glucocorticoid receptor (GR) upregulates EGR1 in the rat and mouse hippocampus, mediating stress-related fear memories | [38,39,40] |
| Connective Tissue Disease | |
| EGR1 is expressed in tendon, cartilage, bone and adipose tissue, and is involved in development, homeostasis, and healing process of these tissues, via regulation of the extracellular matrix EGR1 is often involved in the abnormal production of extracellular matrix in fibrotic conditions such as systemic sclerosis, and EGR1 deletion may be therapeutic for these conditions | 8 |
| Mitochondrial Function | |
| Egr1, Sp1, and SRF are potentially important in mitochondrial biogenesis as their binding sites occur in promoters of important mitochondrial genes; mouse skeletal muscle contraction is associated with marked increase in Egr1 mRNA expression; changes in intracellular Ca(2+) can modify mitochondrial phenotype, in part via the involvement of Egr1 | [41,42,43] |
| Stress-Related Mood Disorders and Schizophrenia | |
| In prefrontal cortex of cadavres of patients with major depression as compared with normal controls, EGR1 is lowered; low EGR1 in PFC was observed in those untreated and also treated but not responding to treatment | [44] |
| In schizophrenia patients, EGR1 mRNA is downregulated in dorsolateral PFC compared with controls | [45,46] |
| EGR1 mRNA in the PFC of schizophrenia patients is positively correlated with glutamic acid decarboxylase 1 (GAD1) mRNA, which is a robust molecular feature of schizophrenia | [46,47] |
| Reduced EGR1 expression is observed in the PVN, mPFC, HPC, or extended amygdala of rats, mice, and prairie voles following social isolation | [48,49,50,51,52,53,54] |
| Drug Reward, Withdrawal and Relapse | |
| Injection of both heroin and morphine upregulates Egr1 mRNA in the nucleus accumbens, dorsal striatum, and cingulate cortex of C57Bl6 mice | [55,56,57,58] |
| In rats and mice, acute ethanol exposure increases EGR1 expression in multiple brain areas including the PFC, amygdala, supraoptic nucleus, hippocampus and nucleus accumbens | [59,60,61,62,63] |
| Cancer | |
| EGR1 regulates angiogenic and osteoclastogenic factors in prostate cancer and promotes metastasis | [64] |
| EGR1 is aberrantly expressed in various cancers, regulating tumour cell proliferation, apoptosis, migration, invasion, and tumour microenvironment. | [65] |
| Description | Reference |
|---|---|
| EBV Transcription | |
| EGR1 is a negative regulator of the EBV C promoter in B lymphoid cell lines EGR1 prevents binding of the positive regulator, SP1 | [123] |
| EBV Lytic Cycle | |
| EGR1 activates expression of BRLF1 via binding to the BRLF1 promoter | [124] |
| EGR1 expression is induced by EBV transactivator Zta | [125] |
| EBV transactivator Zta interacts with methylated ZRE in the EGR1 promoter | [126] |
| Suppression of LMP2A target gene, EGR1, protects Hodgkin’s lymphoma cells from entry into lytic cycle | [127] |
| Cellular immediate early gene (EGR1, EGR2, and EGR3) expression occurs kinetically upstream of EBV BRLF1 and BZLF1 following cross-linking of the B cell receptor in Burkitt lymphoma cells | [128] |
| EBV-Induced B Lymphocyte Transformation | |
| EGR1 expression correlates with the transformed phenotype and type of viral latency in EBV+ lymphoid cell lines | [129] |
| Expression of EGR1 was increased by EBV LMP1 via NFKB, and is required for LMP1-induced cancer cell survival | [130] |
| In EBV+ Hodgkin lymphoma, EBER1 inhibits p21cip1/waf1 transcription and prevents apoptosis through downregulation of p53, EGR1, and STAT1 | [131] |
| In B cell lymphoma, EBV LMP1 increases genomic instability through EGR1-mediated upregulation of activation induced cytidine deaminase | [132] |
| EGR-Associated Pathology | Relevance to ME/CFS | References |
|---|---|---|
| Virus Infection | Numerous viruses have been reported to trigger ME/CFS | [151] |
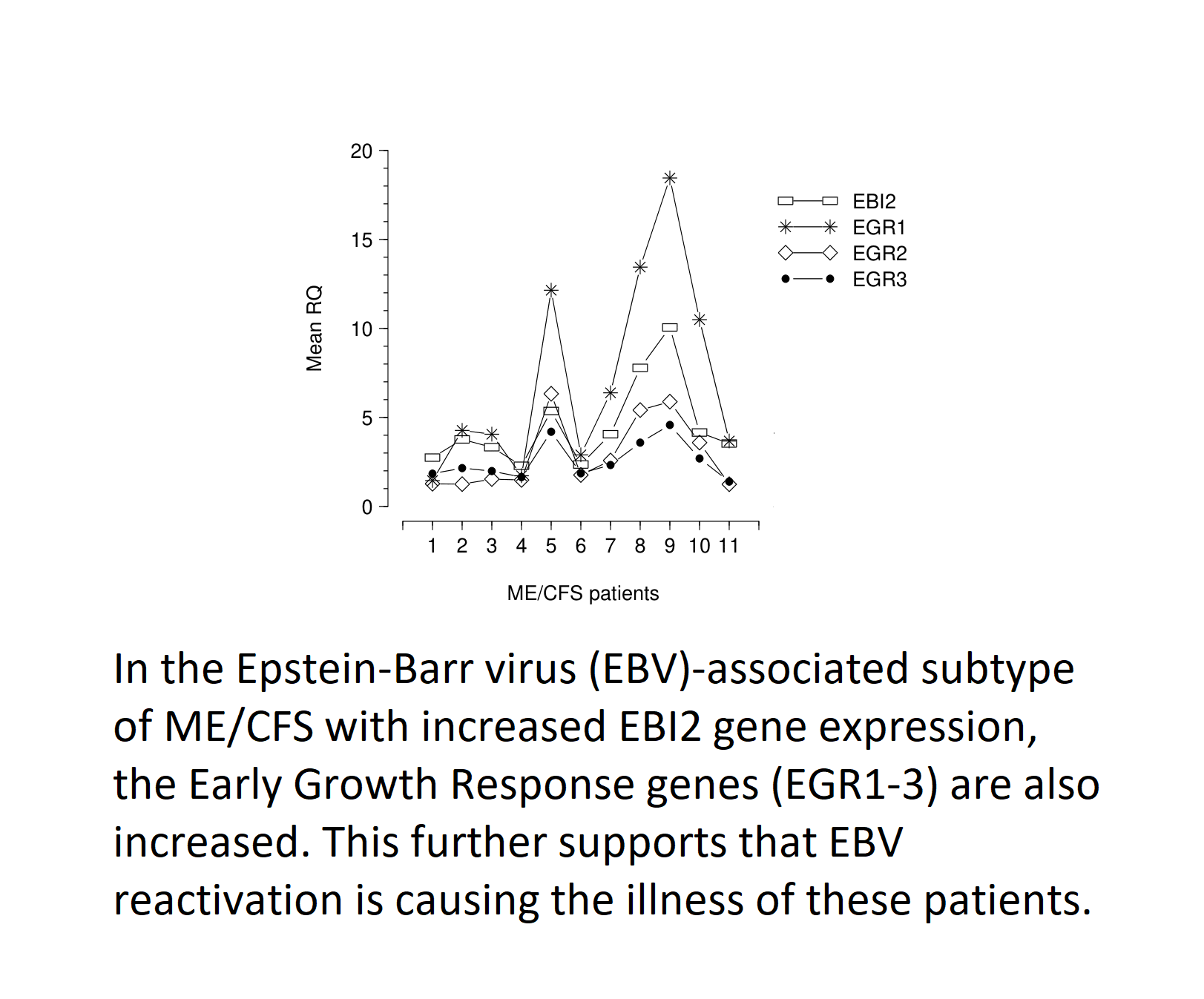
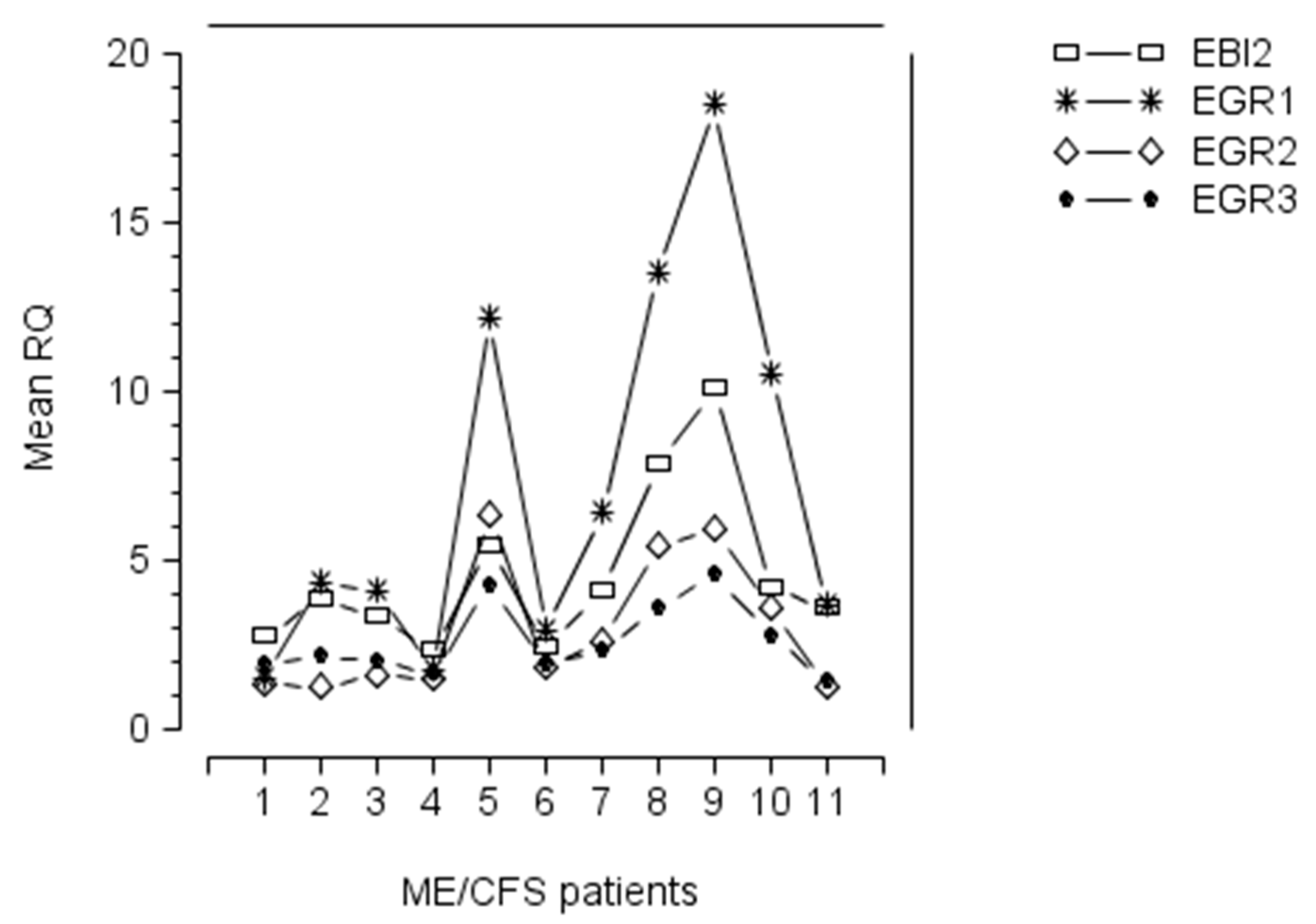
| EBI2 gene upregulation was reported in approximately 50% of ME/CFS patients in one study; EBI2 upregulated patients were severely affected; EBI2 upregulation indicates EBV reactivation | [3] | |
| Immune Response and Inflammation | Increased effector memory CD8+ T cells and decreased terminally differentiated effector CD8+ T cells Significantly increased proportion of mucosal associated invariant T cells (MAIT) cells, especially in severely affected ME/CFS patients | [152] |
| Severe CFS/ME patients differed from controls and moderate CFS/ME patients over time and expressed significant increases in iNKT cell phenotypes, naive CD8+T cells, and γδT cells with significant reduction in NKG2D receptors at 6 months. | [153] | |
| Raised circulating transforming growth factor-β1 has been found in five of eight studies | [154] | |
| ß2 Adrenergic (ß2AdR) and M3 acetylcholine receptor autoantibodies have been found to be elevated in a subset of ME/CFS patients; removal of these autoantibodies by IgG apheresis led to rapid improvement in most patients demonstrating a pathophysiological role of autoantibodies in ME/CFS | [155,156] | |
| Vascular Homeostasis | ME/CFS patients exhibit numerous, well-documented vascular abnormalities, including hypovolaemia, venous pooling, reduced sodium reabsorption, orthostatic dysfunction and postural tachycardia syndrome (POTS), and dominant vagal tone; these abnormalities are likely caused by ß2 adrenergic (ß2AdR) and M3 acetylcholine receptor autoantibodies | [157] |
| Psychological Stress | Psychological stress has been shown to be important in virus transmission, development of symptoms following virus acquisition, as a predisposing factor during the onset of CFS/ME, and in reactivation of EBV | [158,159] |
| Connective Tissue Disease | Pain in muscles and joints are a well-recognised feature of ME/CFS, however, this is likely to be due to systemic effects | [1,2] |
| Mitochondrial Function | Impaired cell metabolism has been documented in ME/CFS | [160,161] |
| ME/CFS CD8+ T cells exhibit reduced mitochondrial membrane potential as compared with those from healthy controls; both CD4+ and CD8+ T cells from patients with ME/CFS exhibit reduced glycolysis at rest, whereas CD8+ T cells also exhibit reduced glycolysis following activation; in addition, proinflammatory cytokines correlated with hypometabolism in T cells | [162] | |
| Stress-Related Mood Disorders and Schizophrenia | None | |
| Drug Reward, Withdrawal and Relapse | None | |
| Cancer | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerr, J. Early Growth Response Gene Upregulation in Epstein–Barr Virus (EBV)-Associated Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Biomolecules 2020, 10, 1484. https://doi.org/10.3390/biom10111484
Kerr J. Early Growth Response Gene Upregulation in Epstein–Barr Virus (EBV)-Associated Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Biomolecules. 2020; 10(11):1484. https://doi.org/10.3390/biom10111484
Chicago/Turabian StyleKerr, Jonathan. 2020. "Early Growth Response Gene Upregulation in Epstein–Barr Virus (EBV)-Associated Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)" Biomolecules 10, no. 11: 1484. https://doi.org/10.3390/biom10111484
APA StyleKerr, J. (2020). Early Growth Response Gene Upregulation in Epstein–Barr Virus (EBV)-Associated Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Biomolecules, 10(11), 1484. https://doi.org/10.3390/biom10111484