Treatment of Diabetic Neuropathy with A Novel PAR1-Targeting Molecule

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
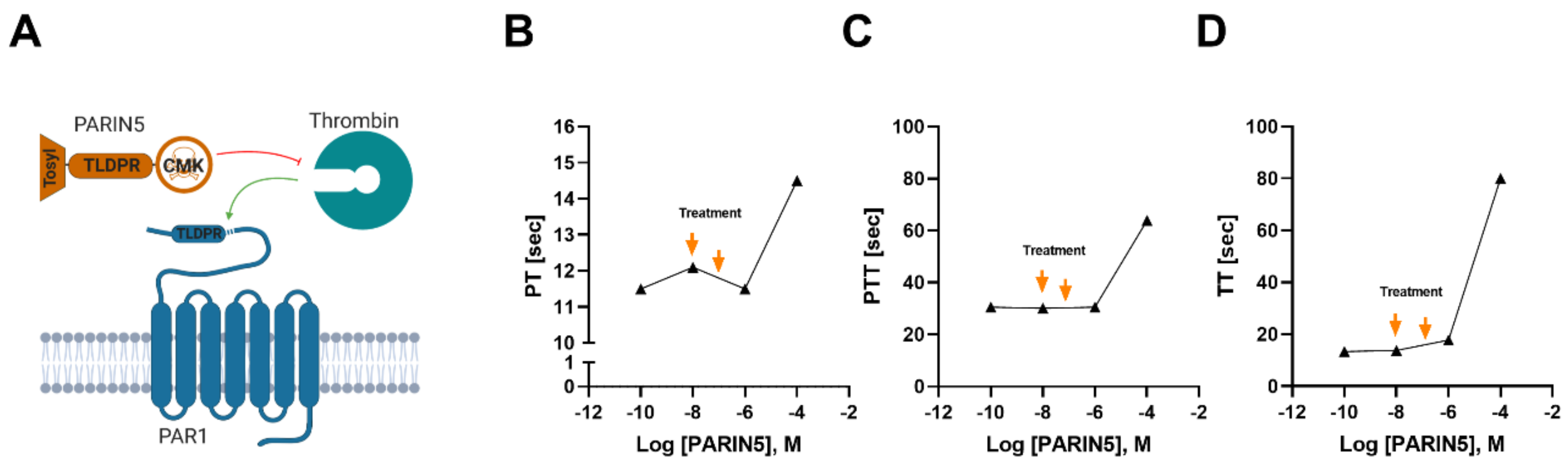
2.1. Molecule Design
2.2. Ethical Considerations
2.3. Induction of Diabetes
2.4. Treatment Protocols
2.5. Coagulation in Plasma
2.6. Motor Function Evaluation
2.7. Sensory Function Evaluation
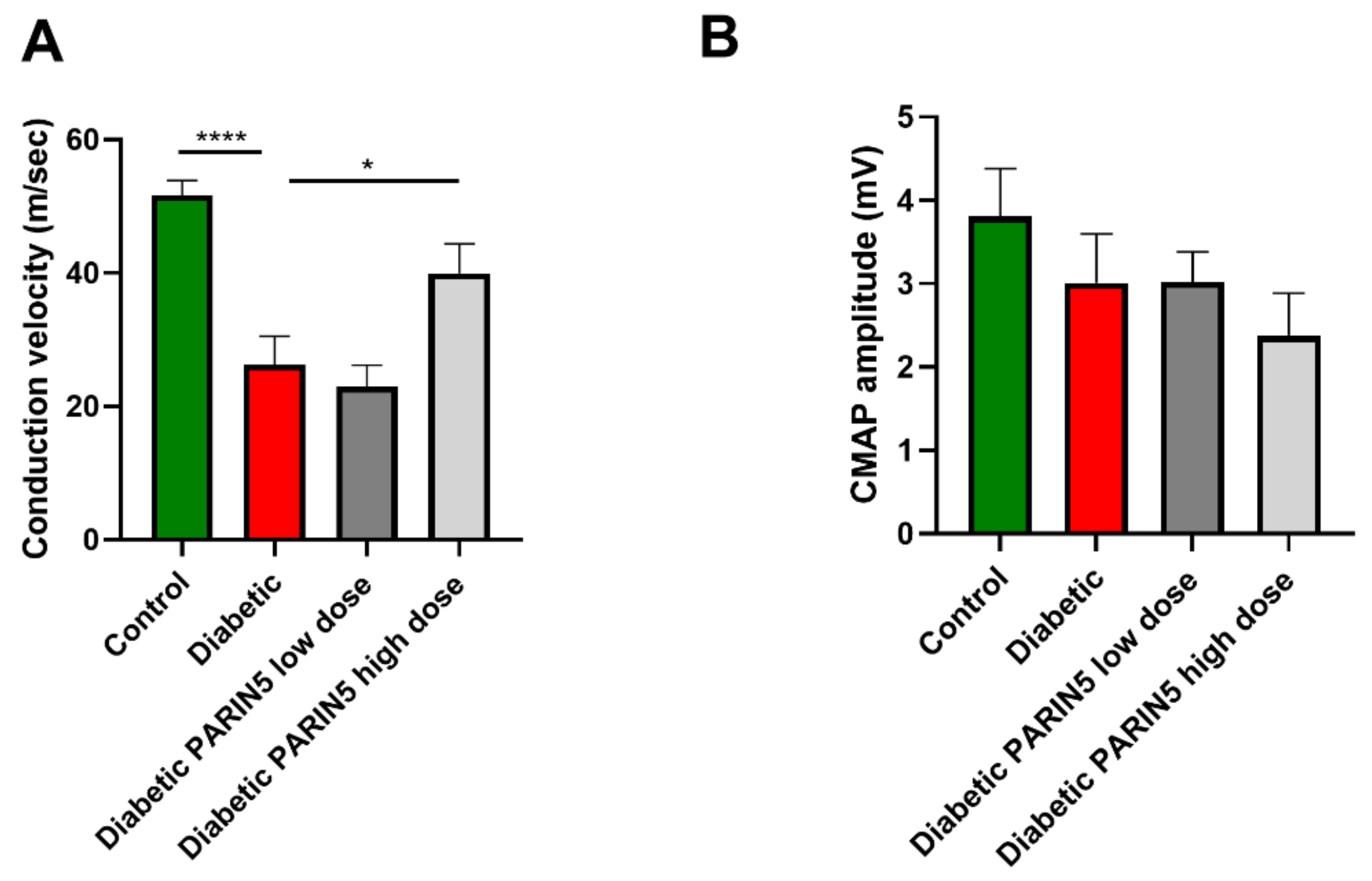
2.8. Nerve Conduction Measurements
2.9. Intraepidermal Nerve Fiber Density (IENFD) Measurement in Skin
2.10. Statistics
3. Results
3.1. Coagulation Studies
3.2. Weight and Blood Glucose Levels
3.3. Motor Function
3.4. Sensory Function
3.5. Nerve Conduction Studies
3.6. Intradermal Nerve Fiber Density
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The worldwide epidemiology of type 2 diabetes mellitus—Present and future perspectives. Nat. Rev. Endocrinol. 2012, 8, 228–236. [Google Scholar] [CrossRef]
- Feldman, E.L.; Russell, J.W.; Sullivan, K.A.; Golovoy, D. New insights into the pathogenesis of diabetic neuropathy. Curr. Opin. Neurol. 1999, 12, 553–563. [Google Scholar] [CrossRef]
- Sumner, C.J.; Sheth, S.; Griffin, J.W.; Cornblath, D.R.; Polydefkis, M. The spectrum of neuropathy in diabetes and impaired glucose tolerance. Neurology 2003, 60, 108–111. [Google Scholar] [CrossRef]
- Said, G. Diabetic neuropathy—A review. Nat. Clin. Pract. Neurol. 2007, 3, 331–340. [Google Scholar] [CrossRef]
- Dugina, T.N.; Kiseleva, E.V.; Lange, M.A.; Vasil’eva, T.V.; Grandfils, C.; Markvicheva, E.A.; Bespalova, Z.D.; Pal’keeva, M.E.; Strukova, S.M. Effect of synthetic peptide thrombin receptor agonist encapsulated in microparticles based on lactic and glycolic acid copolymer on healing of experimental skin wounds in mice. Bull. Exp. Biol. Med. 2004, 138, 463–466. [Google Scholar] [CrossRef]
- Davalos, D.; Baeten, K.M.; Whitney, M.A.; Mullins, E.S.; Friedman, B.; Olson, E.S.; Ryu, J.K.; Smirnoff, D.S.; Petersen, M.A.; Bedard, C.; et al. Early detection of thrombin activity in neuroinflammatory disease. Ann. Neurol. 2014, 75, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Festoff, B.W.; Nelson, P.G.; Brenneman, D.E. Prevention of activity-dependent neuronal death: Vasoactive intestinal polypeptide stimulates astrocytes to secrete the thrombin-inhibiting neurotrophic serpin, protease nexin I. J. Neurobiol. 1996, 30, 255–266. [Google Scholar] [CrossRef]
- Shavit, E.; Beilin, O.; Korczyn, A.D.; Sylantiev, C.; Aronovich, R.; Drory, V.E.; Gurwitz, D.; Horresh, I.; Bar-Shavit, R.; Peles, E.; et al. Thrombin receptor PAR-1 on myelin at the node of Ranvier: A new anatomy and physiology of conduction block. Brain 2008, 131, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Shavit-Stein, E.; Aronovich, R.; Sylantiev, C.; Gofrit, S.G.; Chapman, J.; Dori, A. The role of thrombin in the pathogenesis of diabetic neuropathy. PLoS ONE 2019, 14, e0219453. [Google Scholar] [CrossRef]
- Aoki, I.; Shimoyama, K.; Aoki, N.; Homori, M.; Yanagisawa, A.; Nakahara, K.; Kawai, Y.; Kitamura, S.I.; Ishikawa, K. Platelet-dependent thrombin generation in patients with diabetes mellitus: Effects of glycemic control on coagulability in diabetes. J. Am. Coll. Cardiol. 1996, 27, 560–566. [Google Scholar] [CrossRef] [Green Version]
- Lauria, G.; Hsieh, S.T.; Johansson, O.; Kennedy, W.R.; Leger, J.M.; Mellgren, S.I.; Nolano, M.; Merkies, I.S.J.; Polydefkis, M.; Smith, A.G.; et al. European Federation of Neurological Societies/Peripheral Nerve Society guideline on the use of skin biopsy in the diagnosis of small fiber neuropathy. Report of a joint task force of the European Federation of Neurological Societies and the Peripheral Ner. J. Peripher. Nerv. Syst. 2010, 15, 79–92. [Google Scholar] [CrossRef]
- Gofrit, S.; Shavit-Stein, E. The neuro-glial coagulonome: The thrombin receptor and coagulation pathways as major players in neurological diseases. Neural Regen. Res. 2019, 14, 2043. [Google Scholar] [CrossRef]
- Shavit-Stein, E.; Aronovich, R.; Sylantiev, C.; Gera, O.; Gofrit, S.G.; Chapman, J.; Dori, A. Blocking Thrombin Significantly Ameliorates Experimental Autoimmune Neuritis. Front. Neurol. 2018, 9, 1139. [Google Scholar] [CrossRef] [Green Version]
- Pompili, E.; Ciraci, V.; Leone, S.; De Franchis, V.; Familiari, P.; Matassa, R.; Familiari, G.; Tata, A.M.; Fumagalli, L.; Fabrizi, C. Thrombin regulates the ability of schwann cells to support neuritogenesis and to maintain the integrity of the nodes of ranvier. Eur. J. Histochem. 2020, 64, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Festoff, B.W.; D’Andrea, M.R.; Citron, B.A.; Salcedo, R.M.; Smirnova, I.V.; Andrade-Gordon, P. Motor neuron cell death in wobbler mutant mice follows overexpression of the G-protein-coupled, protease-activated receptor for thrombin. Mol. Med. 2000, 6, 410–429. [Google Scholar] [CrossRef]
- Guo, H.; Zhao, Z.; Yang, Q.; Wang, M.; Bell, R.D.; Wang, S.; Chow, N.; Davis, T.P.; Griffin, J.H.; Goldman, S.A.; et al. An activated protein C analog stimulates neuronal production by human neural progenitor cells via a PAR1-PAR3-S1PR1-Akt pathway. J. Neurosci. 2013, 33, 6181–6190. [Google Scholar] [CrossRef] [Green Version]
- Maggio, N.; Itsekson, Z.; Ikenberg, B.; Strehl, A.; Vlachos, A.; Blatt, I.; Tanne, D.; Chapman, J. The anticoagulant activated protein C (aPC) promotes metaplasticity in the hippocampus through an EPCR-PAR1-S1P1 receptors dependent mechanism. Hippocampus 2014, 24, 1030–1038. [Google Scholar] [CrossRef]
- Pompili, E.; Fabrizi, C.; Somma, F.; Correani, V.; Maras, B.; Schininà, M.E.; Ciraci, V.; Artico, M.; Fornai, F.; Fumagalli, L. PAR1 activation affects the neurotrophic properties of Schwann cells. Mol. Cell. Neurosci. 2017, 79, 23–33. [Google Scholar] [CrossRef]
- Doppler, K.; Frank, F.; Koschker, A.-C.; Reiners, K.; Sommer, C. Nodes of Ranvier in skin biopsies of patients with diabetes mellitus. J. Peripher. Nerv. Syst. 2017, 22, 182–190. [Google Scholar] [CrossRef]
- Walter-Höliner, I.; Barbarini, D.S.; Lütschg, J.; Blassnig-Ezeh, A.; Zanier, U.; Saely, C.H.; Simma, B. High Prevalence and Incidence of Diabetic Peripheral Neuropathy in Children and Adolescents With Type 1 Diabetes Mellitus: Results From a Five-Year Prospective Cohort Study. Pediatr. Neurol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- Shavit-Stein, E.; Sheinberg, E.; Golderman, V.; Sharabi, S.; Wohl, A.; Gofrit, S.G.; Zivli, Z.; Shelestovich, N.; Last, D.; Guez, D.; et al. A Novel Compound Targeting Protease Receptor 1 Activators for the Treatment of Glioblastoma. Front. Neurol. 2018, 9, 1087. [Google Scholar] [CrossRef]
- Hughes, R.A.C.; Cornblath, D.R. Guillain-Barré syndrome. Lancet 2005, 366, 1653–1666. [Google Scholar] [CrossRef]
- Weiner, H.L. Multiple sclerosis is an inflammatory T-cell-mediated autoimmune disease. Arch. Neurol. 2004, 61, 1613–1615. [Google Scholar] [CrossRef]
- Ghanem, K.G. Neurosyphilis: A historical perspective and reviewa. CNS Neurosci. Ther. 2010, 16, e157–e168. [Google Scholar] [CrossRef]
- Ørstavik, K.; Norheim, I.; Jørum, E. Pain and small-fiber neuropathy in patients with hypothyroidism. Neurology 2006, 67, 786–791. [Google Scholar] [CrossRef]
- Kanat, O.; Ertas, H.; Caner, B. Platinum-induced neurotoxicity: A review of possible mechanisms. World J. Clin. Oncol. 2017, 8, 329–333. [Google Scholar] [CrossRef]
- Obrosova, I.G. Diabetic Painful and Insensate Neuropathy: Pathogenesis and Potential Treatments. Neurotherapeutics 2009, 6, 638–647. [Google Scholar] [CrossRef]
- Holland, N.R.; Stocks, A.; Hauer, P.; Cornblath, D.R.; Griffin, J.W.; McArthur, J.C. Intraepidermal nerve fiber density in patients with painful sensory neuropathy. Neurology 1997, 48, 708–711. [Google Scholar] [CrossRef]
- Rubinstein, R.; Genaro, A.M.; Motta, A.; Cremaschi, G.; Wald, M.R. Impaired immune responses in streptozotocin-induced type I diabetes in mice. Involvement of high glucose. Clin. Exp. Immunol. 2008, 154, 235–246. [Google Scholar] [CrossRef]
- Carr, M.E. Diabetes mellitus: A hypercoagulable state. J. Diabetes Complicat. 2001, 15, 44–54. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shavit-Stein, E.; Gofrit, S.G.; Gayster, A.; Teldan, Y.; Ron, A.; Abu Bandora, E.; Golderman, V.; Gera, O.; Harnof, S.; Chapman, J.; et al. Treatment of Diabetic Neuropathy with A Novel PAR1-Targeting Molecule. Biomolecules 2020, 10, 1552. https://doi.org/10.3390/biom10111552
Shavit-Stein E, Gofrit SG, Gayster A, Teldan Y, Ron A, Abu Bandora E, Golderman V, Gera O, Harnof S, Chapman J, et al. Treatment of Diabetic Neuropathy with A Novel PAR1-Targeting Molecule. Biomolecules. 2020; 10(11):1552. https://doi.org/10.3390/biom10111552
Chicago/Turabian StyleShavit-Stein, Efrat, Shany Guly Gofrit, Alexandra Gayster, Yotam Teldan, Ariel Ron, Eiman Abu Bandora, Valery Golderman, Orna Gera, Sagi Harnof, Joab Chapman, and et al. 2020. "Treatment of Diabetic Neuropathy with A Novel PAR1-Targeting Molecule" Biomolecules 10, no. 11: 1552. https://doi.org/10.3390/biom10111552