From Cheese Whey Permeate to Sakacin A: A Circular Economy Approach for the Food-Grade Biotechnological Production of an Anti-Listeria Bacteriocin

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Maintenance
2.2. Quantification of Sakacin a Activity
2.3. Lactobacillus Sakei Growth Evaluation
2.4. Sakacin-A Production and Process Optimization
2.5. Lactic Acid and Amino Acids Determinations
3. Results and Discussion
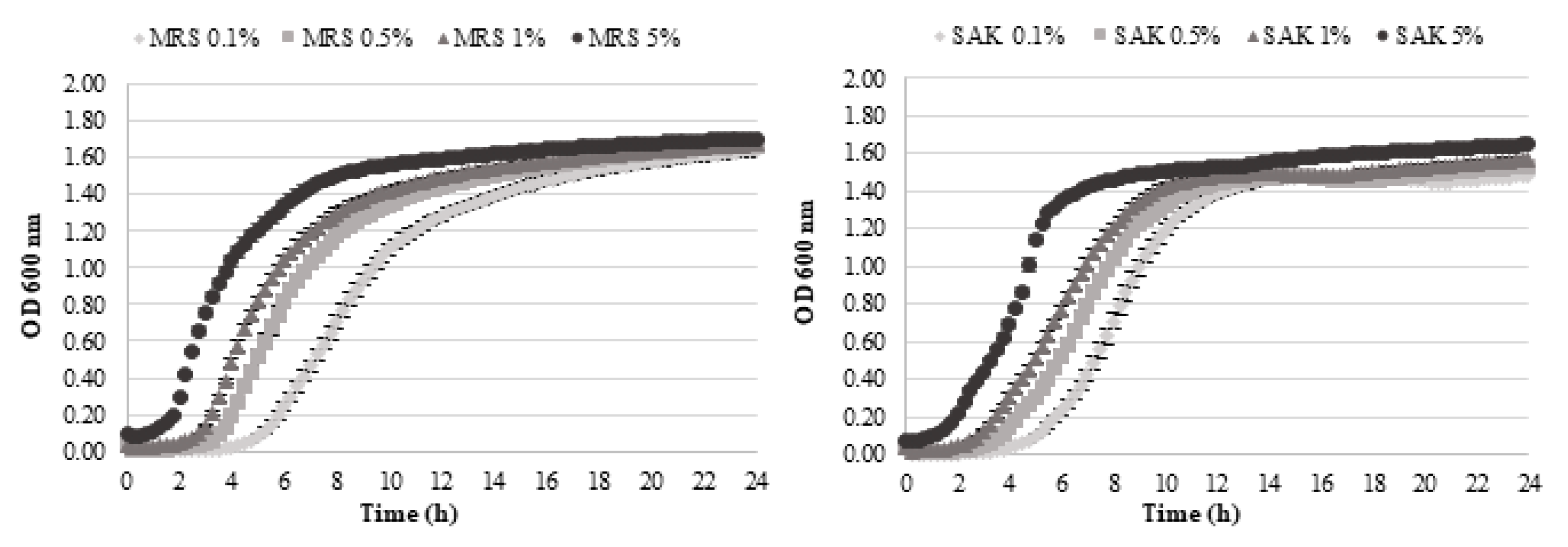
3.1. L. Sakei Growth and Sakacin a Production in MRS and SAK Media
3.2. Optimization of SAK Medium Formulation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gálvez, A.; Abriouel, H.; Lucas López, R.; Ben Omar, N. Bacteriocin-based strategies for food biopreservation. Int. J. Food Microbiol. 2007, 120, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Corsetti, A. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol. 2008, 121, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Garsa, A.K.; Kumariya, S.; Sood, S.K.; Kumar, A.; Kapila, S. Bacteriocin Production and Different Strategies for Their Recovery and Purification. Probiot. Antimicrob. Prot. 2014, 6, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Sidooski, T.; Brandelli, A.; Bertoli, S.L.; Souza, C.K.; Carvalho, L.F. Physical and nutritional conditions for optimized production of bacteriocins by lactic acid bacteria—A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2839–2849. [Google Scholar] [CrossRef]
- Kaškonienė, V.; Stankevičius, M.; Bimbiraitė-Survilienė, K.; Naujokaitytė, G.; Šernienė, L.; Mulkytė, K.; Malakauskas, M.; Maruška, A. Current state of purification, isolation and analysis of bacteriocins produced by lactic acid bacteria. Appl. Microbiol. Biotechnol. 2017, 101, 1323–1335. [Google Scholar] [CrossRef]
- Mapelli, C.; Barbiroli, A.; De Benedetti, S.; Musatti, A.; Rollini, M. Antilisterial bacteriocins for food security: The case of sakacin A. In Encyclopaedia of Food Security and Sustainability; Ferranti, P., Berry, E.M., Anderson, J.R., Eds.; Elsevier: New York, NY, USA, 2018; Volume 2, pp. 385–392. [Google Scholar]
- Ganju, S.; Gogate, P.R. A review on approaches for efficient recovery of whey proteins from dairy industry effluents. J. Food Eng. 2017, 215, 84–96. [Google Scholar] [CrossRef]
- Prazeres, A.R.; Carvalho, F.; Rivas, J. Cheese whey management: A review. J. Environ. Manag. 2012, 110, 48–68. [Google Scholar] [CrossRef]
- Cassano, A.; Conidi, C.; Castro-Muñoz, R. Current and Future Applications of Nanofiltration in Food Processing. In Separation of Functional Molecules in Food by Membrane Technology; Galanakis, C.M., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2019; pp. 305–348. [Google Scholar]
- Zotta, T.; Solieri, L.; Iacumin, L.; Picozzi, C.; Gullo, M. Valorization of cheese whey using microbial fermentations. Appl. Microbiol. Biotechnol. 2020, 104, 2749–2764. [Google Scholar] [CrossRef]
- Lagrange, V.; Whitsett, D.; Burris, C. Global market for dairy proteins. J. Food Sci. 2015, 80, A16–A22. [Google Scholar] [CrossRef]
- European Commission. Prospects for EU Agricultural Markets and Income 2016–2026; European Commission: Brussels, Belgium, 2016; Available online: http://www.euroconsulting.be/wp-content/uploads/2016/12/mp-mto-2016-fullrep_en.pdf (accessed on 15 January 2020).
- OECD/FAO Agricultural Outlook 2019–2028. Available online: https://doi.org/10.1787/agr_outlook-2019-en (accessed on 12 April 2020).
- Pescuma, M.; de Valdez, G.F.; Mozzi, F. Whey-derived valuable products obtained by microbial fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 6183–6196. [Google Scholar] [CrossRef]
- Permeate Market: Global Industry Analysis 2012–2016 and Opportunity Assessment 2017–2027; Future Market Insights: London, UK, 2019.
- Korhonen, H.; Pihlanto-Leppälä, A.; Rantamäki, P.; Tupasela, T. The functional and biological properties of whey proteins: Prospects for the development of functional foods. Agric. Food Sci. 1998, 7, 283–296. [Google Scholar] [CrossRef]
- Božanić, R.; Barukčić, I.; Lisak, K. Possibilities of whey utilisation. Austin J. Nutr. Food Sci. 2014, 2, 1036–1043. [Google Scholar]
- Gonzalez, M.I.; Alvarez, S.; Riera, F.; Alvarez, R. Economic evaluation of an integrated process for lactic acid production from ultrafiltered whey. J. Food Eng. 2007, 80, 553–561. [Google Scholar] [CrossRef]
- Wolf, O.; Crank, M.; Patel, M.; Marscheider-Weidemann, F.; Schleich, J.; Hüsing, B.; Angerer, G. Techno-economic feasibility of large-scale production of bio-based polymers in Europe. Eur. Communities 2005, 50–64. [Google Scholar]
- Espinosa-Gonzalez, I.; Parashar, A.; Bressler, D.C. Heterotrophic growth and lipid accumulation of Chlorella protothecoides in whey permeate, a dairy by-product stream, for biofuel production. Bioresour. Technol. 2014, 155, 170–176. [Google Scholar] [CrossRef]
- Colombo, B.; Calvo, M.V.; Sciarria, T.P.; Scaglia, B.; Kizito, S.S.; D’Imporzano, G.; Adani, F. Biohydrogen and polyhydroxyalkanoates (PHA) as products of a two-steps bioprocess from deproteinized dairy wastes. Waste Manag. 2019, 95, 22–31. [Google Scholar] [CrossRef]
- Ende, S.S.; Noke, A. Heterotrophic microalgae production on food waste and by-products. J. Appl. Phycol. 2019, 31, 1565–1571. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations/World Health Organization (FAO/WHO). Risk assessment of Listeria Monocytogenes in Ready-To-Eat Foods, Technical Report. Microbiological Risk Assessment. 2004. Available online: https://apps.who.int/iris/handle/10665/42874 (accessed on 20 January 2020).
- Delves-Broughton, J. Natural antimicrobials as additives and ingredients for the preservation of foods and beverages. In Natural Food Additives, Ingredients and Flavourings, 1st ed.; Baines, D., Seal, R., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition: Cambridge, UK, 2012; pp. 127–161. [Google Scholar]
- Axelsson, L.; Holck, A. The genes involved in production of and immunity to sakacin A, a bacteriocin from Lactobacillus sake Lb706. J. Bacteriol. 1995, 177, 2125–2137. [Google Scholar] [CrossRef] [Green Version]
- Barbiroli, A.; Musatti, A.; Capretti, G.; Iametti, S.; Rollini, M. Sakacin-A antimicrobial packaging to reduce Listeria contamination in thin-cut meat. J. Sci. Food Agric. 2017, 97, 1042–1047. [Google Scholar] [CrossRef]
- Mapelli, C.; Musatti, A.; Barbiroli, A.; Seini, S.; Bras, J.; Cavicchioli, D.; Rollini, M. Cellulose nano-fibers (CNF)—Sakacin-A active material: Production, characterization and application in storage trials of smoked salmon. J. Sci. Food Agric. 2019, 99, 4731–4738. [Google Scholar] [CrossRef] [Green Version]
- Future Market Insights. Protective Packaging Market: Global Industry Analysis and Opportunity Assessment 2015–2025. Available online: http://www.futuremarketinsights.com/reports/protective-packaging-market (accessed on 28 January 2020).
- de la Caba, K.; Guerrero, P.; Trung, T.S.; Cruz-Romero, M.; Kerry, J.P.; Fluhr, J.; Maurer, M.; Kruijssen, F.; Albalat, A.; Bunting, S.; et al. From seafood waste to active seafood packaging: An emerging opportunity of the circular economy. J. Clean. Prod. 2019, 208, 86–98. [Google Scholar] [CrossRef]
- Trinetta, V.; Rollini, M.; Manzoni, M. Development of a low cost culture medium for sakacin A production by L. Sakei. Proc. Biochem. 2008, 43, 1275–1280. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.S. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Hogenboom, J.A.; D’Incecco, P.; Fuselli, F.; Pellegrino, L. Ion-exchange chromatographic method for the determination of the free amino acid composition of cheese and other dairy products: An inter-laboratory validation study. Food Anal. Meth. 2017, 10, 3137–3148. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.K.; Kjems, J.; Mygind, T.; Snabe, T.; Meyer, R.L. Effects of Tween 80 on Growth and Biofilm Formation in Laboratory Media. Front. Microbiol. 2016, 7, 1878. [Google Scholar] [CrossRef]
- D’Incecco, P.; Gatti, M.; Hogenboom, J.A.; Bottari, B.; Rosi, V.; Neviani, E.; Pellegrino, L. Lysozyme affects the microbial catabolism of free arginine in raw-milk hard cheeses. Food Microbiol. 2016, 57, 16–22. [Google Scholar] [CrossRef]
- Campos, C.A.; Castro, M.P.; Rivas, F.P.; Schelegueda, L.I. Bacteriocins in food: Evaluation of the factors affecting their effectiveness. In Microbial Pathogens and Strategies for Combating them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 994–1004. [Google Scholar]
- Irmler, S.; Bavan, T.; Oberli, A.; Roetschi, A.; Badertscher, R.; Guggenbühl, B.; Berthoud, H. Catabolism of Serine by Pediococcus acidilactici and Pediococcus pentosaceus. Appl. Environ. Microbiol. 2013, 79, 1309–1315. [Google Scholar] [CrossRef] [Green Version]
- McLeod, A.; Mosleth, E.F.; Rud, I.; Branco dos Santos, F.; Snipen, L.; Liland, K.H. Effects of glucose availability in Lactobacillus sakei: Metabolic change and regulation of the proteome and transcriptome. PLoS ONE 2017, 12, e0187542. [Google Scholar] [CrossRef]
- Montanari, C.; Barbieri, F.; Magnani, M.; Grazia, L.; Gardini, F.; Tabanelli, G. Phenotypic diversity of Lactobacillus sakei strains. Front. Microbiol. 2018, 9, 2003. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Medium | Ingredient | Cost €/Kg | g/L | €/L | Share of SAK Cost (%) |
|---|---|---|---|---|---|
| MRS | As purchased | 104 | 52.2 | 5.40 | - |
| SAK | Yeast Extract | 60 | 8 | 0.48 | 21 |
| Meat Extract | 149 | 8 | 1.19 | 52 | |
| Arginine | 415 | 0.5 | 0.20 | 9 | |
| Tween-80 | 83.7 | 0.5 | 0.04 | 2 | |
| Vitamin mix | 0.37 €/mL | 1 mL | 0.37 | 16 | |
| CWP | 0.015 €/L | 1 L | 0.015 | 1 | |
| TOTAL | 2.30 | 100 |
| Inoculum Medium | 0.1% (v/v) MRS SAK | 0.5% (v/v) MRS SAK | 1% (v/v) MRS SAK | 5% (v/v) MRS SAK | |||||
|---|---|---|---|---|---|---|---|---|---|
| Lag phase (h) | mean | 3.267 A | 5.266 B | 2.581 A | 3.794 B | 1.854 A | 2.895 B | 0.010 A | 1.839 B |
| std. dev | 0.009 | 0.109 | 0.063 | 0.111 | 0.083 | 0.061 | 0.010 | 0.067 | |
| Growth rate (OD/h) | mean | 0.200 A | 0.255 B | 0.219 A | 0.239 B | 0.237 A | 0.250 B | 0.241 A | 0.306 B |
| std. dev | 0.009 | 0.004 | 0.007 | 0.002 | 0.009 | 0.002 | 0.014 | 0.009 | |
| Final Value (OD) | mean | 1.517 B | 1.459 A | 1.550 B | 1.452 A | 1.558 B | 1.496 A | 1.613 B | 1.534 A |
| std. dev | 0.031 | 0.013 | 0.005 | 0.063 | 0.016 | 0.018 | 0.019 | 0.005 | |
| Sakacin A | AU/mL | 267 | 233 | 267 | 233 | 267 | 267 | 300 | 333 |
| Time (h) | 16 | 16 | 16 | 16 | 12 | 12 | 8 | 8 | |
| Trial | A-ME (g/L) | B-YE (g/L) | C-Arg (g/L) | Culture pH | Lactic Acid (g/L) | Sakacin A Prod. (mm) | |||
|---|---|---|---|---|---|---|---|---|---|
| Mean Std. dev | Mean Std. dev. | Mean Std. dev | |||||||
| 1 | 0 | 0 | 0.5 | 6.32 | 0.04 | 0.01 | 0.00 | 0.0 | 0.0 |
| 2 | 0 | 4 | 0.5 | 5.15 | 0.07 | 1.35 | 0.00 | 7.0 | 0.0 |
| 3 | 0 | 8 | 0.5 | 5.02 | 0.09 | 1.84 | 0.21 | 9.5 | 0.7 |
| 4 | 4 | 0 | 0.5 | 6.18 | 0.02 | 0.68 | 0.00 | 8.0 | 0.0 |
| 5 | 4 | 4 | 0.5 | 5.64 | 0.00 | 1.56 | 0.01 | 9.5 | 0.7 |
| 6 | 4 | 8 | 0.5 | 5.14 | 0.16 | 2.35 | 0.13 | 11.5 | 0.7 |
| 7 | 8 | 0 | 0.5 | 6.18 | 0.04 | 0.98 | 0.01 | 10.0 | 0.0 |
| 8 | 8 | 4 | 0.5 | 5.72 | 0.00 | 1.81 | 0.05 | 11.5 | 0.7 |
| 9 | 8 | 8 | 0.5 | 5.35 | 0.03 | 2.59 | 0.02 | 13.0 | 0.0 |
| 10 | 0 | 0 | 1 | 6.49 | 0.02 | 0.01 | 0.00 | 0.0 | 0.0 |
| 11 | 0 | 4 | 1 | 5.23 | 0.19 | 1.20 | 0.18 | 3.0 | 0.0 |
| 12 | 0 | 8 | 1 | 5.16 | 0.11 | 1.70 | 0.33 | 8.0 | 0.0 |
| 13 | 4 | 0 | 1 | 6.45 | 0.05 | 0.80 | 0.01 | 7.5 | 0.7 |
| 14 | 4 | 4 | 1 | 5.97 | 0.10 | 1.57 | 0.05 | 10.0 | 0.0 |
| 15 | 4 | 8 | 1 | 5.44 | 0.18 | 2.47 | 0.04 | 11.5 | 0.7 |
| 16 | 8 | 0 | 1 | 6.49 | 0.04 | 1.03 | 0.06 | 10.0 | 0.0 |
| 17 | 8 | 4 | 1 | 6.06 | 0.01 | 1.88 | 0.04 | 11.0 | 0.0 |
| 18 | 8 | 8 | 1 | 5.63 | 0.15 | 2.66 | 0.13 | 12.0 | 0.0 |
| 19 | 0 | 0 | 0 | 6.10 | 0.01 | 0.03 | 0.01 | 0.0 | 0.0 |
| 20 | 0 | 4 | 0 | 4.99 | 0.06 | 1.33 | 0.10 | 7.0 | 0.0 |
| 21 | 0 | 8 | 0 | 4.85 | 0.05 | 1.93 | 0.00 | 9.5 | 0.7 |
| 22 | 4 | 0 | 0 | 5.54 | 0.07 | 0.69 | 0.06 | 7.5 | 0.7 |
| 23 | 4 | 4 | 0 | 5.13 | 0.11 | 1.45 | 0.07 | 9.0 | 0.0 |
| 24 | 4 | 8 | 0 | 4.90 | 0.07 | 2.11 | 0.26 | 10.5 | 0.7 |
| 25 | 8 | 0 | 0 | 5.63 | 0.04 | 0.90 | 0.06 | 8.5 | 0.7 |
| 26 | 8 | 4 | 0 | 5.24 | 0.04 | 1.74 | 0.06 | 10.5 | 0.7 |
| 27 | 8 | 8 | 0 | 5.05 | 0.04 | 2.44 | 0.07 | 11.5 | 0.7 |
| MRS | - | - | - | 4.45 | 0.10 | 5.62 | 0.18 | 13.0 | 0.7 |
| Parameter | pH | Lactic Acid | Sakacin A Production |
|---|---|---|---|
| Model | p < 0.0001 Linear | p < 0.0001 Square root | p < 0.0001 Linear |
| A-ME | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| B-YE | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| C-Arg | p < 0.0001 | ns * | p < 0.0001 |
| AB | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| AC | p < 0.0001 | ns | p < 0.0001 |
| BC | p 0.0093 | ns | p 0.0040 |
| ABC | ns | ns | p 0.0008 |
| Lack of fit | 0.457 ns | 0.1066 ns | - (Saturated) |
| Model std. dev | 0.086 | 0.048 | 0.047 |
| mean | 5.59 | 1.13 | 0.85 |
| C.V.% | 1.53 | 4.25 | 5.56 |
| R2 | 0.9828 | 0.9894 | 0.9916 |
| Adj R2 | 0.9740 | 0.9875 | 0.9834 |
| Pred R2 | 0.9591 | 0.9847 | 0.9663 |
| Adeq precision | 32.449 | 75.839 | 39.000 |
| Amino Acid | YE 8-ME 8-ARG 0 | YE 8-ME 8-ARG 0.5 | ||||
|---|---|---|---|---|---|---|
| (mg/L) | t0 | 16 h | (%) | t0 | 16 h | (%) |
| Alanine | 171.0 | 183.8 | 7 | 169.3 | 183.4 | 8 |
| Arginine | 194.0 | 0.0 | −100 | 526.0 | 0.0 | −100 |
| Asparagine | 78.0 | 63.6 | −18 | 76.5 | 56.7 | −26 |
| Aspartate | 138.4 | 127.8 | −8 | 137.5 | 131.9 | −4 |
| Citrulline | 26.5 | 24.1 | −9 | 25.8 | 24.9 | −3 |
| Cysteine | 0.0 | 0.0 | 0 | 0.0 | 0.0 | 0 |
| GABA | 10.2 | 10.4 | 2 | 10.1 | 10.5 | 4 |
| Glutamine | 0.0 | 0.0 | 0 | 0.0 | 0.0 | 0 |
| Glutamate | 402.6 | 429.9 | 7 | 392.3 | 423.6 | 8 |
| Glycine | 70.0 | 89.0 | 27 | 69.1 | 89.9 | 30 |
| Histidine | 44.2 | 49.3 | 12 | 45.8 | 50.6 | 10 |
| Isoleucine | 131.7 | 175.4 | 33 | 129.9 | 175.9 | 35 |
| Leucine | 254.5 | 296.0 | 16 | 251.2 | 293.5 | 17 |
| Lysine | 189.5 | 216.8 | 14 | 185.7 | 214.9 | 16 |
| Methionine | 50.2 | 51.9 | 3 | 48.0 | 53.0 | 10 |
| Ornithine | 15.8 | 142.4 | 802 | 15.1 | 233.8 | 1449 |
| Phenilalanine | 180.3 | 201.1 | 12 | 180.6 | 202.4 | 12 |
| Proline | 52.8 | 70.2 | 33 | 52.0 | 71.3 | 37 |
| Serine | 98.4 | 11.4 | −88 | 94.5 | 5.2 | −95 |
| Threonine | 89.4 | 124.9 | 40 | 88.7 | 117.9 | 33 |
| Tryptophan | 40.1 | 54.1 | 35 | 39.6 | 54.4 | 37 |
| Tyrosine | 95.7 | 141.4 | 48 | 138.8 | 180.7 | 30 |
| Valine | 176.6 | 221.1 | 25 | 177.2 | 220.7 | 25 |
| Medium | ME (g/L) | YE (g/L | Arg (g/L) | pH | Lactic Acid (g/L) | Desirability | Medium Cost (€/L) | Sak. A Yield (106 AU/L) | Sak. A Cost (€/106 AU) |
|---|---|---|---|---|---|---|---|---|---|
| 1 (SAK) | 8 | 8 | 0.5 | 5.35 | 2.56 | 0.907 | 2.30 | 0.289 | 7.96 |
| 2 | 8 | 8 | 0 | 5.09 | 2.56 | 0.907 | 2.13 | 0.242 | 8.80 |
| 3 | 4 | 8 | 0 | 4.84 | 2.30 | 0.878 | 1.53 | 0.270 | 5.67 |
| 4 | 4 | 8 | 0.5 | 5.16 | 2.30 | 0.870 | 1.73 | 0.278 | 6.22 |
| 5 | 0 | 8 | 0.5 | 4.98 | 1.82 | 0.825 | 1.14 | 0.190 | 7.37 |
| 6 | 4 | 8 | 1 | 5.47 | 2.30 | 0.823 | 1.93 | 0.233 | 8.28 |
| 7 | 8 | 8 | 1 | 5.65 | 2.56 | 0.819 | 2.53 | 0.233 | 10.86 |
| MRS | - | - | - | 4.45 | 5.62 | - | 5.40 | 0.300 | 18.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musatti, A.; Cavicchioli, D.; Mapelli, C.; Bertoni, D.; Hogenboom, J.A.; Pellegrino, L.; Rollini, M. From Cheese Whey Permeate to Sakacin A: A Circular Economy Approach for the Food-Grade Biotechnological Production of an Anti-Listeria Bacteriocin. Biomolecules 2020, 10, 597. https://doi.org/10.3390/biom10040597
Musatti A, Cavicchioli D, Mapelli C, Bertoni D, Hogenboom JA, Pellegrino L, Rollini M. From Cheese Whey Permeate to Sakacin A: A Circular Economy Approach for the Food-Grade Biotechnological Production of an Anti-Listeria Bacteriocin. Biomolecules. 2020; 10(4):597. https://doi.org/10.3390/biom10040597
Chicago/Turabian StyleMusatti, Alida, Daniele Cavicchioli, Chiara Mapelli, Danilo Bertoni, Johannes A. Hogenboom, Luisa Pellegrino, and Manuela Rollini. 2020. "From Cheese Whey Permeate to Sakacin A: A Circular Economy Approach for the Food-Grade Biotechnological Production of an Anti-Listeria Bacteriocin" Biomolecules 10, no. 4: 597. https://doi.org/10.3390/biom10040597