Assessment of Fatty Acids Profile and Omega-3 Polyunsaturated Fatty Acid Production by the Oleaginous Marine Thraustochytrid Aurantiochytrium sp. T66 Cultivated on Volatile Fatty Acids





Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Cultivation
2.2. Batch Cultivation on Different Volatile Fatty Acids
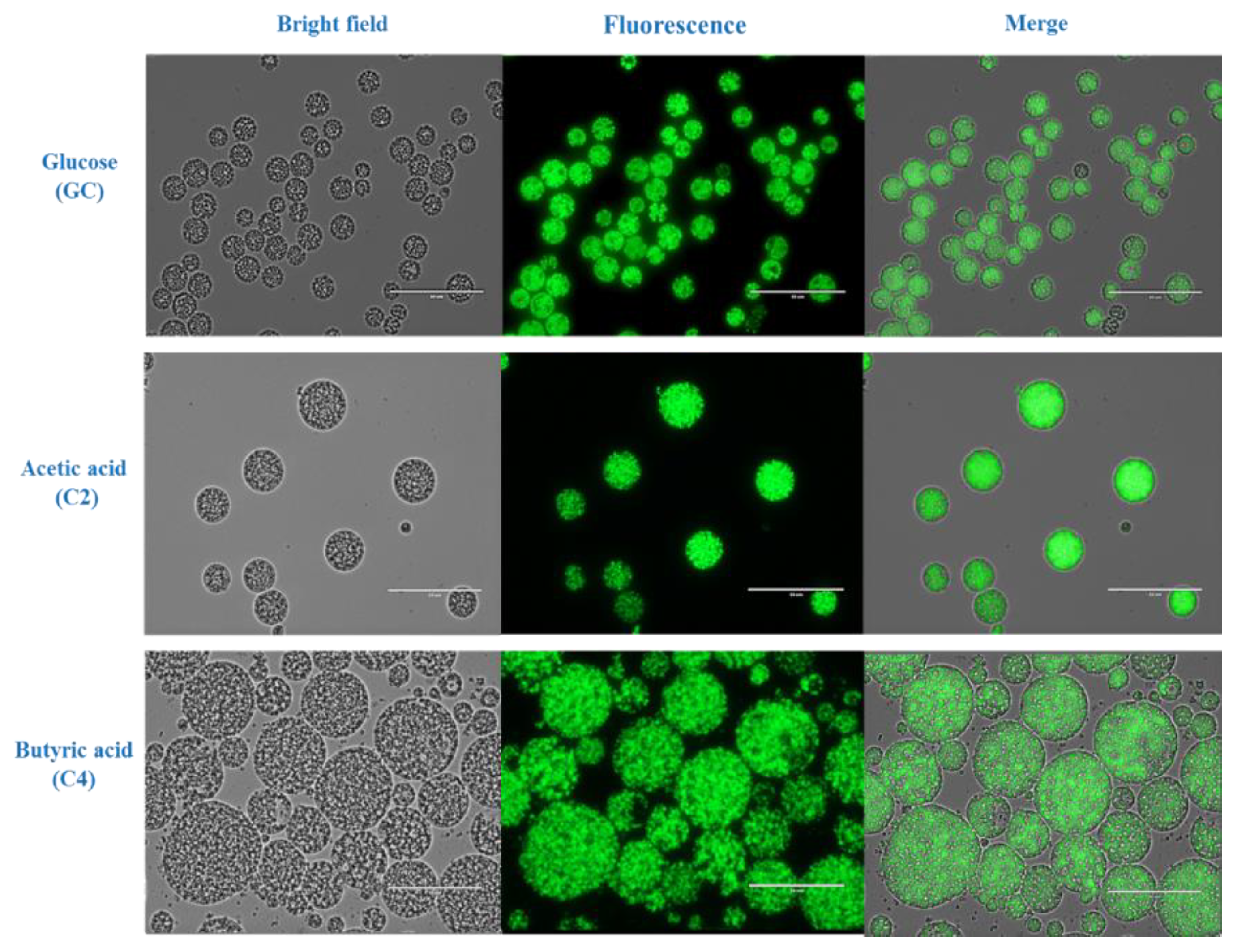
2.3. Morphologic Analysis and Estimation of Lipid Accumulation by Fluorescence Microscopy
2.4. Cell Dry Weight (CDW) Determination
2.5. Total Lipid Determination
2.6. Residual Carbon Estimation by High-Performance Liquid Chromatography (HPLC)
2.7. Analysis of Fatty Acid Profile by Gas Chromatography-Mass Spectrometry (GC-MS)
2.8. Statistical Analysis
3. Results and Discussion
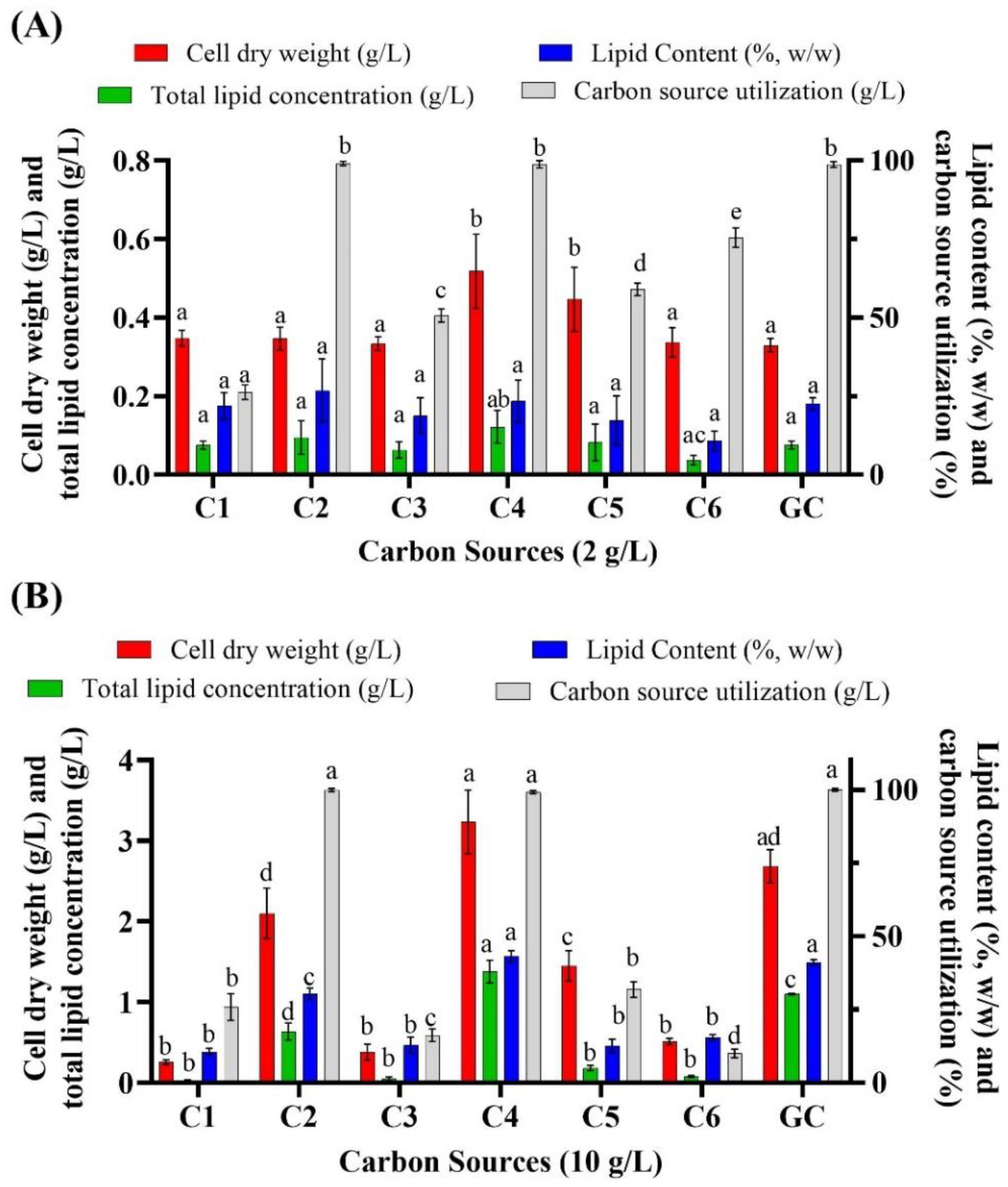
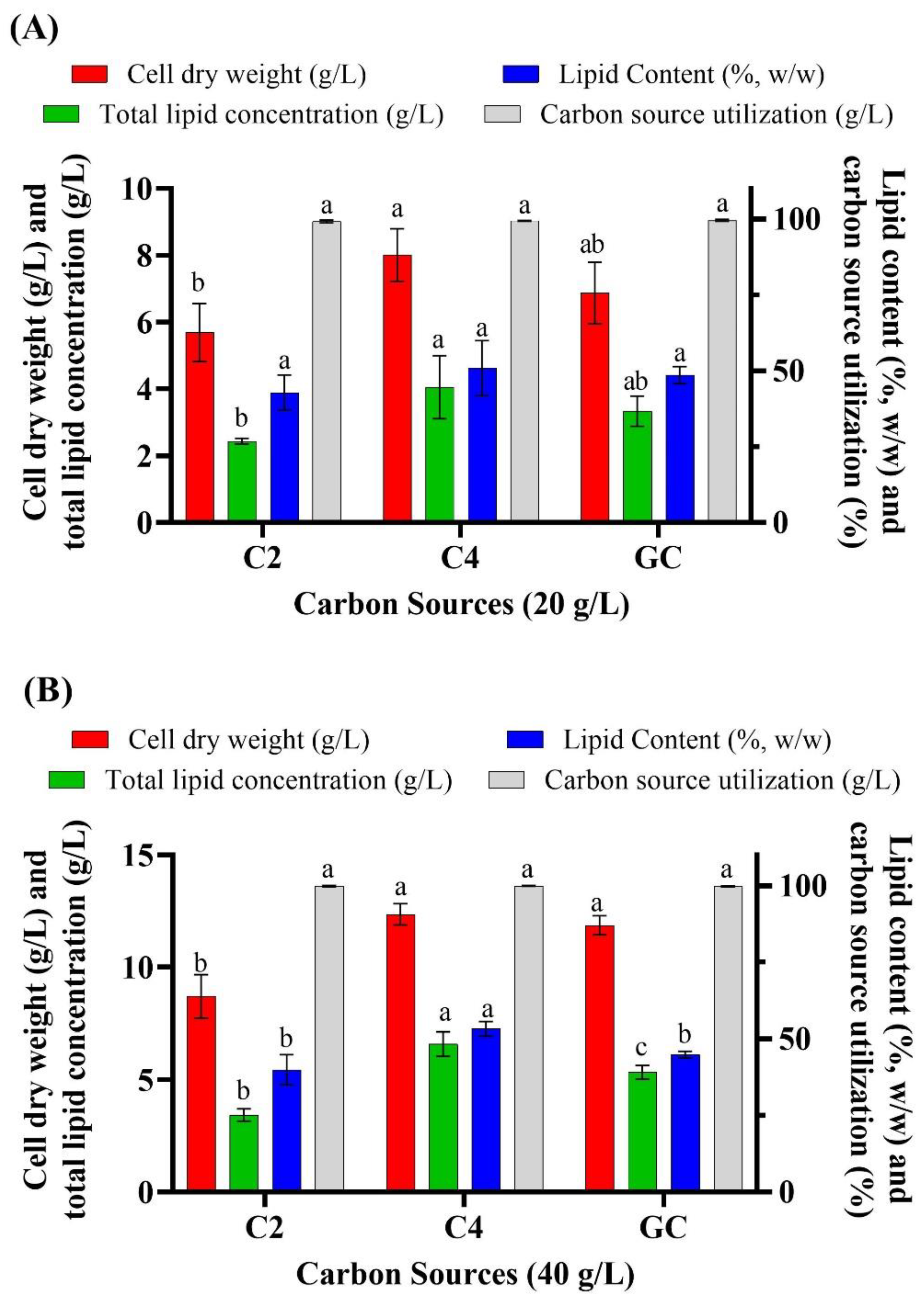
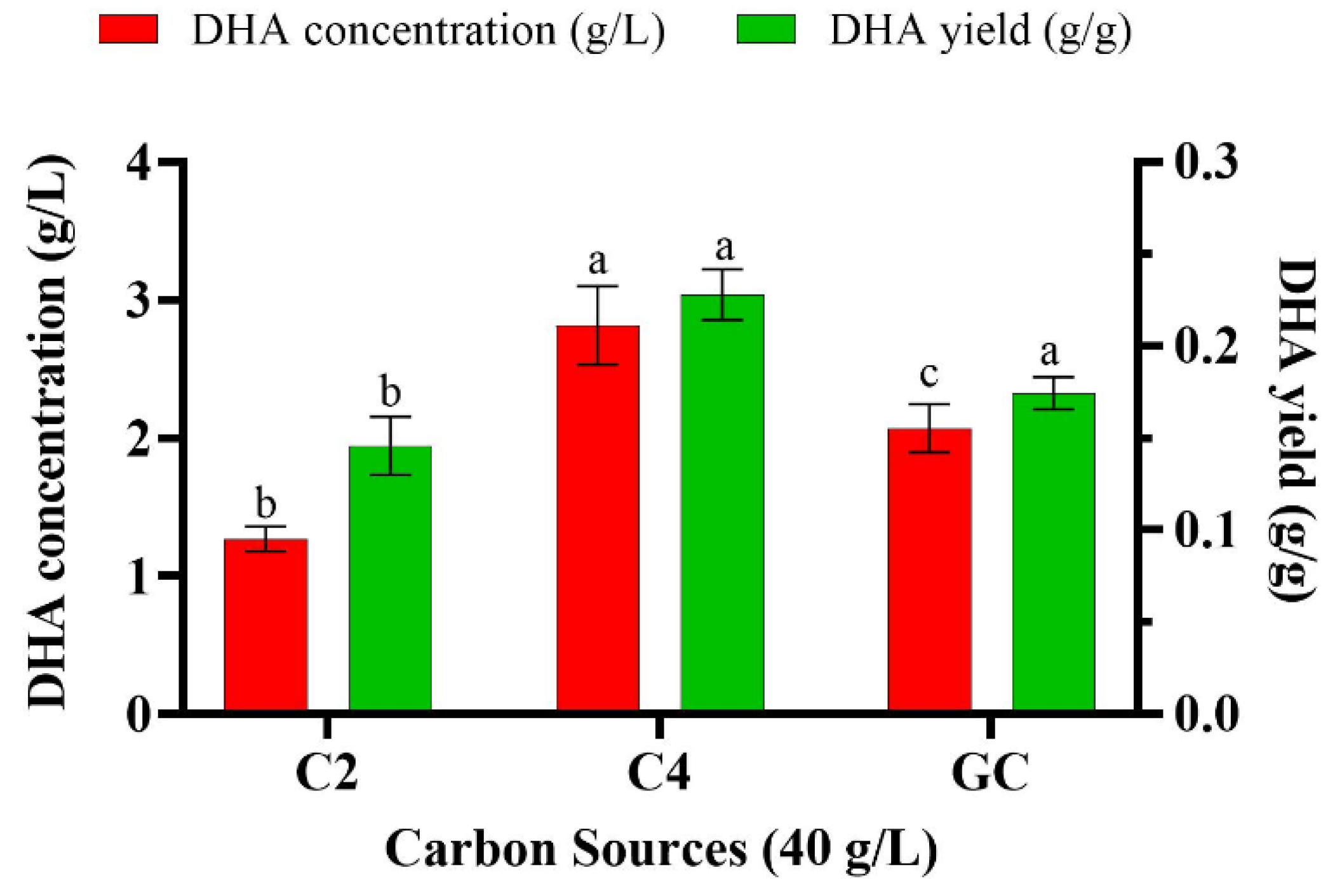
3.1. Batch Cultivation of Aurantiochytrium sp. T66 on Different Volatile Fatty Acids
3.2. Effect of Various Volatile Fatty Acids on the Fatty Acid Profile of Aurantiochytrium sp. T66
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kaur, G.; Cameron-Smith, D.; Garg, M.; Sinclair, A.J. Docosapentaenoic acid (22:5n-3): A review of its biological effects. Prog. Lipid Res. 2011, 50, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, C.C.; Su, K.P.; Cheng, T.C.; Liu, H.C.; Chang, C.J.; Dewey, M.E.; Stewart, R.; Huang, S.Y. The effects of omega-3 fatty acids monotherapy in Alzheimer’s disease and mild cognitive impairment: A preliminary randomized double-blind placebo-controlled study. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2008, 32, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Lazzarin, N.; Vaquero, E.; Exacoustos, C.; Bertonotti, E.; Romanini, M.E.; Arduini, D. Low-dose aspirin and omega-3 fatty acids improve uterine artery blood flow velocity in women with recurrent miscarriage due to impaired uterine perfusion. Fertil. Steril. 2009, 92, 296–300. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K.; Belury, M.A.; Andridge, R.; Malarkey, W.B.; Hwang, B.S.; Glaser, R. Omega-3 supplementation lowers inflammation in healthy middle-aged and older adults: A randomized controlled trial. Brain. Behav. Immun. 2012, 26, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA effects in brain development and function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M. Dietary omega 3 fatty acids and the developing brain. Brain Res. 2008, 1237, 35–43. [Google Scholar] [CrossRef]
- Guesnet, P.; Alessandri, J. Biochimie Docosahexaenoic acid (DHA) and the developing central nervous system (CNS) e implications for dietary recommendations. Biochimie 2011, 93, 7–12. [Google Scholar] [CrossRef]
- Elagizi, A.; Lavie, C.J.; Marshall, K.; DiNicolantonio, J.J.; O’Keefe, J.H.; Milani, R.V. Omega-3 polyunsaturated fatty acids and cardiovascular health: A comprehensive review. Prog. Cardiovasc. Dis. 2018, 61, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Harris, W.S.; Mozaffarian, D.; Rimm, E.; Kris-Etherton, P.; Rudel, L.L.; Appel, L.J.; Engler, M.M.; Engler, M.B.; Sacks, F. Omega-6 fatty acids and risk for cardiovascular disease: A science advisory from the American heart association nutrition subcommittee of the council on nutrition, physical activity, and metabolism; council on cardiovascular nursing; and council on epidem. Circulation 2009, 119, 902–907. [Google Scholar] [CrossRef] [Green Version]
- Kris-Etherton, P.; Fleming, J.; Harris, W.S. The debate about n-6 polyunsaturated fatty acid recommendations for cardiovascular health. J. Am. Diet. Assoc. 2010, 110, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Wanasundara, U.N. Omega-3 fatty acid concentrates: Nutritional aspects and production technologies. Trends Food Sci. Technol. 1998, 9, 230–240. [Google Scholar] [CrossRef]
- Pejin, B.; Bianco, A.; Newmaster, S.; Sabovljevic, M.; Vujisic, L.; Tesevic, V.; Vajs, V.; De Rosa, S. Fatty acids of Rhodobryum ontariense (Bryaceae). Nat. Prod. Res. 2012, 26, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Vujisic, L.; Sabovljevic, M.; Tesevic, V.; Vajs, V. The moss mnium hornum, a promising source of arachidonic acid. Chem. Nat. Compd. 2012, 48, 120–121. [Google Scholar] [CrossRef]
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Process Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Alonso, D.L.; Maroto, F.G. Plants as “chemical factories” for the production of polyunsaturated fatty acids. Biotechnol. Adv. 2000, 18, 481–497. [Google Scholar] [CrossRef]
- Domingo, J.L.; Bocio, A.; Falcó, G.; Llobet, J.M. Benefits and risks of fish consumption. Part I. A quantitative analysis of the intake of omega-3 fatty acids and chemical contaminants. Toxicology 2007, 30, 219–226. [Google Scholar] [CrossRef]
- Patel, A.; Liefeldt, S.; Rova, U.; Christakopoulos, P.; Matsakas, L. Co-production of DHA and squalene by thraustochytrid from forest biomass. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Raghukumar, S. Thraustochytrid marine protists: Production of PUFAs and other emerging technologies. Mar. Biotechnol. 2008, 631–640. [Google Scholar] [CrossRef]
- Vigani, M.; Parisi, C.; Rodríguez-Cerezo, E.; Barbosa, M.J.; Sijtsma, L.; Ploeg, M.; Enzing, C. Food and feed products from micro-algae: Market opportunities and challenges for the EU. Trends Food Sci. Technol. 2015, 42, 81–92. [Google Scholar] [CrossRef]
- Bannenberg, G.; Mallon, C.; Edwards, H.; Yeadon, D.; Yan, K.; Johnson, H.; Ismail, A. Omega-3 long-chain polyunsaturated fatty acid content and oxidation state of fish oil supplements in New Zealand. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subhadra, B.G.; Edwards, M. Coproduct market analysis and water footprint of simulated commercial algal biorefineries. Appl. Energy 2011, 88, 3515–3523. [Google Scholar] [CrossRef]
- Li, X.; Xu, H.; Wu, Q. Large-scale biodiesel production from microalga Chlorella protothecoides through heterotrophic cultivation in bioreactors. Biotechnol. Bioeng. 2007, 98, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Park, G.W.; Fei, Q.; Jung, K.; Chang, H.N.; Kim, Y.C.; Kim, N.J.; Choi, J.d.R.; Kim, S.; Cho, J. Volatile fatty acids derived from waste organics provide an economical carbon source for microbial lipids/biodiesel production. Biotechnol. J. 2014, 9, 1536–1546. [Google Scholar] [CrossRef] [PubMed]
- Leong, H.Y.; Su, C.A.; Lee, B.S.; Lan, J.C.W.; Law, C.L.; Chang, J.S.; Show, P.L. Development of Aurantiochytrium limacinum SR21 cultivation using salt-rich waste feedstock for docosahexaenoic acid production and application of natural colourant in food product. Bioresour. Technol. 2019, 271, 30–36. [Google Scholar] [CrossRef]
- Ferreira, G.F.; Ríos Pinto, L.F.; Maciel Filho, R.; Fregolente, L.V. A review on lipid production from microalgae: Association between cultivation using waste streams and fatty acid profiles. Renew. Sustain. Energy Rev. 2019, 109, 448–466. [Google Scholar] [CrossRef]
- Choi, J.D.R.; Chang, H.N.; Han, J.I. Performance of microbial fuel cell with volatile fatty acids from food wastes. Biotechnol. Lett. 2011, 33, 705–714. [Google Scholar] [CrossRef]
- Mengmeng, C.; Hong, C.; Qingliang, Z.; Shirley, S.N.; Jie, R. Optimal production of polyhydroxyalkanoates (PHA) in activated sludge fed by volatile fatty acids (VFAs) generated from alkaline excess sludge fermentation. Bioresour. Technol. 2009, 100, 1399–1405. [Google Scholar] [CrossRef]
- Huang, Y.L.; Wu, Z.; Zhang, L.; Ming Cheung, C.; Yang, S.T. Production of carboxylic acids from hydrolyzed corn meal by immobilized cell fermentation in a fibrous-bed bioreactor. Bioresour. Technol. 2002, 82, 51–59. [Google Scholar] [CrossRef]
- Akaraonye, E.; Keshavarz, T.; Roy, I. Production of polyhydroxyalkanoates: The future green materials of choice. J. Chem. Technol. Biotechnol. 2010, 85, 732–743. [Google Scholar] [CrossRef]
- Kondo, T.; Kondo, M. Efficient production of acetic acid from glucose in a mixed culture of Zymomonas mobilis and Acetobacter sp. J. Ferment. Bioeng. 1996, 81, 42–46. [Google Scholar] [CrossRef]
- Fei, Q.; Fu, R.; Shang, L.; Brigham, C.J.; Chang, H.N. Lipid production by microalgae Chlorella protothecoides with volatile fatty acids (VFAs) as carbon sources in heterotrophic cultivation and its economic assessment. Bioprocess Biosyst. Eng. 2015, 38, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, Q.; Chang, H.N.; Shang, L.; Choi, J.d.R.; Kim, N.J.; Kang, J.W. The effect of volatile fatty acids as a sole carbon source on lipid accumulation by Cryptococcus albidus for biodiesel production. Bioresour. Technol. 2011, 102, 2695–2701. [Google Scholar] [CrossRef] [PubMed]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioconversion of volatile fatty acids into lipids by the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Chalima, A.; Hatzidaki, A.; Karnaouri, A.; Topakas, E. Integration of a dark fermentation effluent in a microalgal-based biorefinery for the production of high-added value omega-3 fatty acids. Appl. Energy 2019, 241, 130–138. [Google Scholar] [CrossRef]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Simultaneous production of DHA and squalene from Aurantiochytrium sp. grown on forest biomass hydrolysates. Biotechnol. Biofuels 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Heterotrophic cultivation of Auxenochlorella protothecoides using forest biomass as a feedstock for sustainable biodiesel production. Biotechnol. Biofuels 2018, 11, 169. [Google Scholar] [CrossRef]
- Van Wychen, S.; Ramirez, K.; Laurens, L.M. Determination of total lipids as fatty acid methyl esters (FAME) by in situ transesterification. Contract 2013, 303, 275–3000. [Google Scholar]
- Ratledge, C.; Kanagachandran, K.; Anderson, A.J.; Grantham, D.J.; Stephenson, J.C. Production of docosahexaenoic acid by Crypthecodinium cohnii grown in a pH-auxostat culture with acetic acid as principal carbon source. Lipids 2001, 36, 1241–1246. [Google Scholar] [CrossRef]
- Wen, Q.; Chen, Z.; Li, P.; Duan, R.; Ren, N. Lipid production for biofuels from hydrolyzate of waste activated sludge by heterotrophic Chlorella protothecoides. Bioresour. Technol. 2013, 143, 695–698. [Google Scholar] [CrossRef]
- Turon, V.; Trably, E.; Fouilland, E.; Steyer, J.P. Growth of Chlorella sorokiniana on a mixture of volatile fatty acids: The effects of light and temperature. Bioresour. Technol. 2015, 198, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.F.; Young, K.D.; Swaffer, M.; Wood, E.; Nurse, P.; Kimura, A.; Frankel, J.; Wallingford, J.; Walbot, V.; Qu, X.; et al. What determines cell size? BMC Biol. 2012, 10, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morabito, C.; Bournaud, C.; Maës, C.; Schuler, M.; Aiese Cigliano, R.; Dellero, Y.; Maréchal, E.; Amato, A.; Rébeillé, F. The lipid metabolism in thraustochytrids. Prog. Lipid Res. 2019, 76, 101007. [Google Scholar] [CrossRef] [PubMed]
- Fossier, M.L.; Lee, C.K.J.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef]
- Qiu, X. Biosynthesis of docosahexaenoic acid (DHA, 22:6-4, 7,10,13,16,19): Two distinct pathways. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 181–186. [Google Scholar] [CrossRef]
- Monroig, Ó.; Wang, S.; Zhang, L.; You, C.; Tocher, D.R.; Li, Y. Elongation of long-chain fatty acids in rabbitfish Siganus canaliculatus: Cloning, functional characterisation and tissue distribution of Elovl5- and Elovl4-like elongases. Aquaculture 2012, 350–353, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Sijtsma, L.; De Swaaf, M.E. Biotechnological production and applications of the -3 polyunsaturated fatty acid docosahexaenoic acid. Appl. Microbiol. Biotechnol. 2004, 64, 146–153. [Google Scholar] [CrossRef]
- Ye, C.; Qiao, W.; Yu, X.; Ji, X.; Huang, H.; Collier, J.L.; Liu, L. Reconstruction and analysis of the genome-scale metabolic model of Schizochytrium limacinum SR21 for docosahexaenoic acid production. BMC Genom. 2015, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wallis, J.G.; Watts, J.L.; Browse, J. Polyunsaturated fatty acid synthesis: What will they think of next? Trends Biochem. Sci. 2002, 27, 467–473. [Google Scholar] [CrossRef]
- Metz, J.G.; Roessler, P.; Facciotti, D.; Levering, C.; Dittrich, F.; Lassner, M.; Valentine, R.; Lardizabal, K.; Domergue, F.; Yamada, A.; et al. Production of polyunsaturated fatty acids by potyketide synthases in both prokaryotes and eukaryotes. Science 2001, 293, 290–293. [Google Scholar] [CrossRef]
- Qiu, X.; Hong, H.; Mackenzie, S.L. Identification of a Δ4 Fatty Acid Desaturase from Thraustochytrium sp. Involved in the Biosynthesis of Docosahexanoic Acid by Heterologous Expression in Saccharomyces cerevisiae and Brassica juncea *. J. Biol. Chem. 2001, 276, 31561–31566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weete, J.D.; Kim, H.; Gandhi, S.R.; Wang, Y.; Dute, R. Lipids and Ultrastructure of Thraustochytrium sp. ATCC 26185. Lipids 1997, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.E.; Nichols, P.D.; Mcmeekin, T.A. The biotechnological potential of Thraustochytrids. Mar. Biotechnol. 1999, 580–587. [Google Scholar] [CrossRef] [PubMed]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable biodiesel production from oleaginous yeasts utilizing hydrolysates of various non-edible lignocellulosic biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef]
- Boyle, N.R.; Morgan, J.A. Flux balance analysis of primary metabolism in Chlamydomonas reinhardtii. BMC Syst. Biol. 2009, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Harrop, L.C.; Kornberg, H.L. The role of isocitrate lyase in the metabolism of algae. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1966, 166, 11–29. [Google Scholar]
- Combres, C.; Laliberté, G.; Reyssac, J.S.; de la Noüe, J. Effect of acetate on growth and ammonium uptake in the microalga Scenedesmus obliquus. Physiol. Plant. 1994, 91, 729–734. [Google Scholar] [CrossRef]
- Lopes Da Silva, T.; Reis, A. The use of multi-parameter flow cytometry to study the impact of n-dodecane additions to marine dinoflagellate microalga Crypthecodinium cohnii batch fermentations and DHA production. J. Ind. Microbiol. Biotechnol. 2008, 35, 875–887. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids (%) in Total Lipid | C1 | C2 | C3 | C4 | C5 | C6 | GC |
|---|---|---|---|---|---|---|---|
| C14:0 | 6.04 ± 0.32 b | 7.50 ± 0.28 c | 7.26 ± 0.12 c,d | 6.46 ± 0.57 b,c | 5.44 ± 0.16 b | 6.90 ± 0.52 b,c | 21.59 ± 0.30 a |
| C14:1 | 12.12 ± 0.58 b | 14.89 ± 0.63 c | 17.60 ± 0.81 d | 17.30 ± 0.41 d | 12.64 ± 0.42 b | 25.93 ± 1.14 a | ND |
| C15:0 | ND | ND | ND | ND | ND | ND | 18.60 ± 0.73 |
| C15:1 | 3.67 ± 0.61 b | 3.42 ± 0.42 b | 4.14 ± 0.13 b | 4.44 ± 0.65 b | 3.09 ± 0.23 b | 8.38 ± 0.34 a | ND |
| C 16:0 | 35.44 ± 0.82 b | 37.72 ± 0.94 c | 32.90 ± 1.23 f | 38.46 ± 0.60 c,d | 36.44 ± 0.73 b,c | 32.63 ± 0.42 e,f | 44.31 ± 0.63 a |
| C16:1 | 8.54 ± 1.17 a | 8.17 ± 0.64 a | 8.18 ± 0.82 a | 7.97± 0.35 a | 8.68 ± 1.00 a | 7.73 ± 0.92 a | ND |
| C 18:0 | 18.66 ± 0.92 b | 22.45 ± 0.58 c | 25.58 ± 1.08 d | 23.23 ±1.19 c,e | 27.79 ± 0.24 a,d | 17.736 ± 0.89 b | 9.64 ± 0.35 f |
| C18:1 | 14.88 ± 1.40 d | 5.75 ± 0.48 a,b | 4.37 ± 0.73 a,b | 2.16 ± 0.41 b,c | 5.81 ± 0.89 a | ND | ND |
| DPA | ND | ND | ND | ND | ND | ND | 2.65 ± 0.37 |
| DHA | ND | ND | ND | ND | ND | ND | 2.48 ± 0.52 |
| Fatty Acids (%) in Total Lipid | C1 | C2 | C3 | C4 | C5 | C6 | GC |
|---|---|---|---|---|---|---|---|
| C14:0 | 6.92 ± 0.42 b | 4.79 ± 0.32 c | 9.00 ± 0.40 d | 15.08 ± 0.37 a | 7.74 ± 0.35 b,d | 5.09 ± 0.30 c | 14.99 ± 1.13 a |
| C14:1 | ND | ND | ND | ND | ND | 17.37 ± 0.72 | ND |
| C15:0 | 18.54 ± 0.49 d | 3.49 ± 0.03 c | 19.59 ± 0.90 a | 10.25 ± 0.28 b | 22.56 ± 1.28 a | 8.39 ± 0.82 b | 21.37 ± 0.85 a |
| C 16:0 | 28.43 ± 0.85 b | 23.22 ± 0.32 c | 29.58 ± 0.82 b | 34.19 ± 1.25 d | 30.19 ± 1.05 b | 31.29 ± 1.01 b,e | 42.79 ± 0.47 a |
| C16:1 | ND | ND | ND | ND | ND | 5.10 ± 0.49 | ND |
| C17:0 | 9.22 ± 0.69 a | ND | 9.27 ± 0.45 a | 5.77 ± 0.55 b | 8.92 ± 0.38 a | ND | 8.56 ± 0.93 a |
| C 18:0 | 14.44 ± 0.75 a | 8.90 ± 0.52 b | 14.89 ± 1.37 a | 6.74 ± 0.49 c | 14.32 ± 0.55 a | 8.63 ± 0.78 b | 5.79 ± 0.33 c |
| C18:1 | ND | 1.84 ± 0.49 | ND | ND | ND | 9.46 ± 0.89 | ND |
| C20:0 | ND | ND | ND | ND | ND | 5.58 ± 0.81 | ND |
| C20:4 | 2.31 ± 0.27 b | 5.13 ± 0.57 a | 1.03 ± 0.14 c | ND | 0.96 ± 0.05 c,d | ND | ND |
| EPA | 2.32 ± 0.16 b | 5.24 ± 0.05 a | 1.60 ± 0.27 c | ND | 1.94 ± 0.14 b,c | ND | ND |
| DPA | 7.52 ± 0.60 b | 18.29 ± 0.78 a | 7.27 ± 0.78 b | 8.88 ± 0.41 b | 4.87 ± 0.41 d | 2.41± 0.12 c | 2.20 ± 0.13 c |
| DHA | 9.57± 0.68 b | 28.73 ± 1.28 a | 7.28 ± 0.21 c | 18.64 ± 0.59 d | 7.88 ± 0.54 b,c | 6.01 ± 0.53 c | 4.14 ± 0.28 c |
| Fatty Acids (%) in Total Lipid | C2 | C4 | GC |
|---|---|---|---|
| C14:0 | 2.85 ± 0.35 b | 6.67 ± 0.31 a | 8.33 ± 0.58 a |
| C15:0 | 10.32 ± 0.40 a | 2.60 ± 0.18 b | 8.03 ± 0.10 c |
| C 16:0 | 25.59 ± 0.43 a | 19.79 ± 0.77 b | 18.54 ± 1.04 b |
| C16:1 | 5.48 ± 0.68 | 2.37 ± 0.35 | ND |
| C 18:0 | 3.67 ± 0.17 b | 3.00 ± 0.53 b | 7.99 ± 0.40 a |
| C18:1 | ND | 1.33 ± 0.16 | ND |
| C20:4 | 1.74 ± 0.17 | 5.23 ± 0.53 | ND |
| EPA | 0.72 ± 0.13 b | 1.89 ± 0.48 a | 1.59 ± 0.43 a |
| DPA | 12.16 ± 0.14 c | 13.93 ± 0.48 b | 17.00 ± 0.62 a |
| DHA | 36.93 ± 0.69 a | 42.63 ± 1.08 b | 38.72 ± 1.04 a |
| Fatty Acids (%) in Total Lipid | C2 | C4 | GC |
|---|---|---|---|
| C14:0 | 7.65 ± 0.23 a | 2.45 ± 0.17 b | 5.31 ± 0.04 b |
| C15:0 | 8.93 ± 0.28 b | 3.43 ± 0.18 c | 18.3 ± 0.76 a |
| C 16:0 | 22.78 ± 0.46 a | 20.34 ± 0.48 b | 15.58 ± 1.02 c |
| C17:0 | 7.11 ± 0.26 b | 1.02 ± 0.13 c | 11.74 ± 0.37 a |
| C 18:0 | 2.45 ± 0.91 b | 4.47 ± 0.73 a | 4.043 ± 0.40 a,b |
| C20:4 | 1.17 ± 0.14 | 5.28 ± 0.37 | ND |
| EPA | 1.65 ± 0.37 b | 3.12 ± 0.16 a | 0.82 ± 0.03 c |
| DPA | 17.94 ± 0.29 b | 23.35 ± 0.81 a | 14.54 ± 0.72 c |
| DHA | 30.40 ± 0.64 b | 36.25 ± 1.17 a | 29.58 ± 0.95 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Assessment of Fatty Acids Profile and Omega-3 Polyunsaturated Fatty Acid Production by the Oleaginous Marine Thraustochytrid Aurantiochytrium sp. T66 Cultivated on Volatile Fatty Acids. Biomolecules 2020, 10, 694. https://doi.org/10.3390/biom10050694
Patel A, Rova U, Christakopoulos P, Matsakas L. Assessment of Fatty Acids Profile and Omega-3 Polyunsaturated Fatty Acid Production by the Oleaginous Marine Thraustochytrid Aurantiochytrium sp. T66 Cultivated on Volatile Fatty Acids. Biomolecules. 2020; 10(5):694. https://doi.org/10.3390/biom10050694
Chicago/Turabian StylePatel, Alok, Ulrika Rova, Paul Christakopoulos, and Leonidas Matsakas. 2020. "Assessment of Fatty Acids Profile and Omega-3 Polyunsaturated Fatty Acid Production by the Oleaginous Marine Thraustochytrid Aurantiochytrium sp. T66 Cultivated on Volatile Fatty Acids" Biomolecules 10, no. 5: 694. https://doi.org/10.3390/biom10050694
APA StylePatel, A., Rova, U., Christakopoulos, P., & Matsakas, L. (2020). Assessment of Fatty Acids Profile and Omega-3 Polyunsaturated Fatty Acid Production by the Oleaginous Marine Thraustochytrid Aurantiochytrium sp. T66 Cultivated on Volatile Fatty Acids. Biomolecules, 10(5), 694. https://doi.org/10.3390/biom10050694