Phage Display Selection, Identification, and Characterization of Novel Pancreatic Cancer Targeting Peptides


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Lines
2.3. Phage Display Selection
2.4. Next-Generation Sequencing and Bioinformatic Analysis
2.5. Peptides
2.6. Fluorescent Microscopy
2.7. Modified ELISA
2.8. Statistical Analysis
3. Results
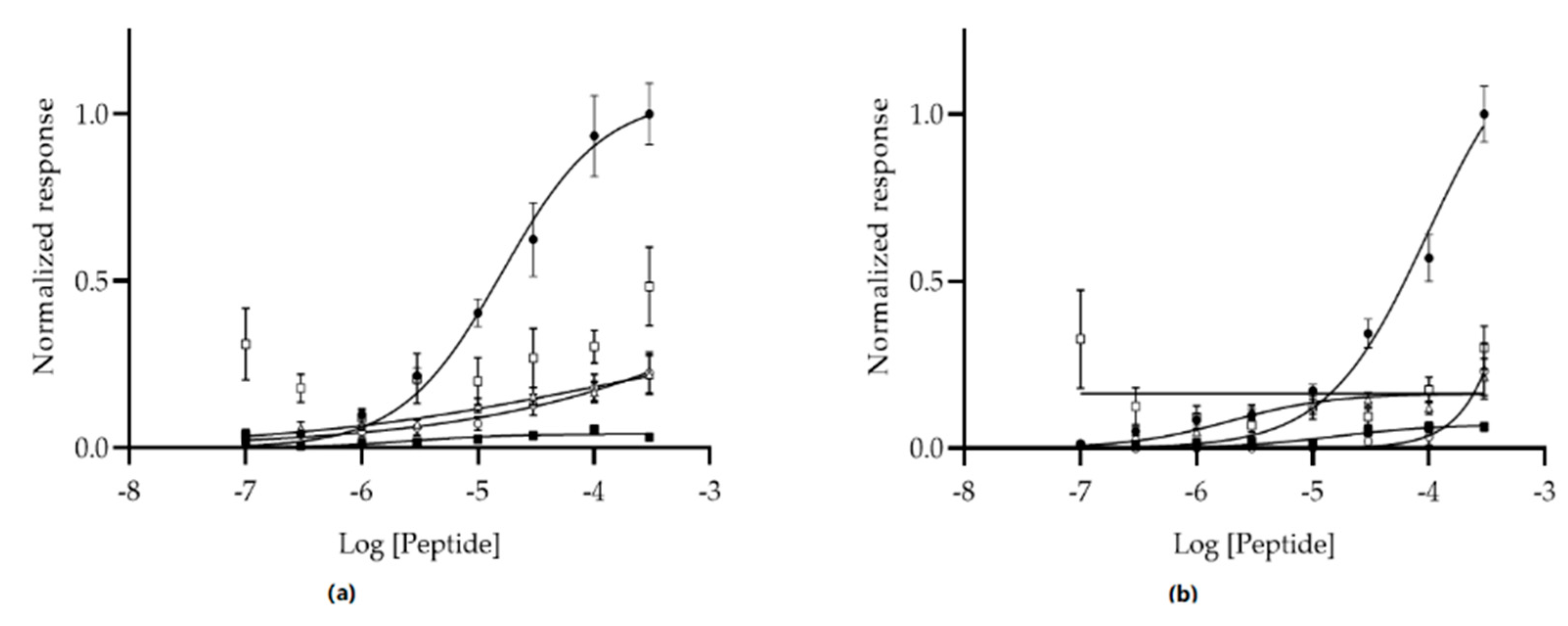
3.1. Phage Display Selection and Next Generation Sequencing
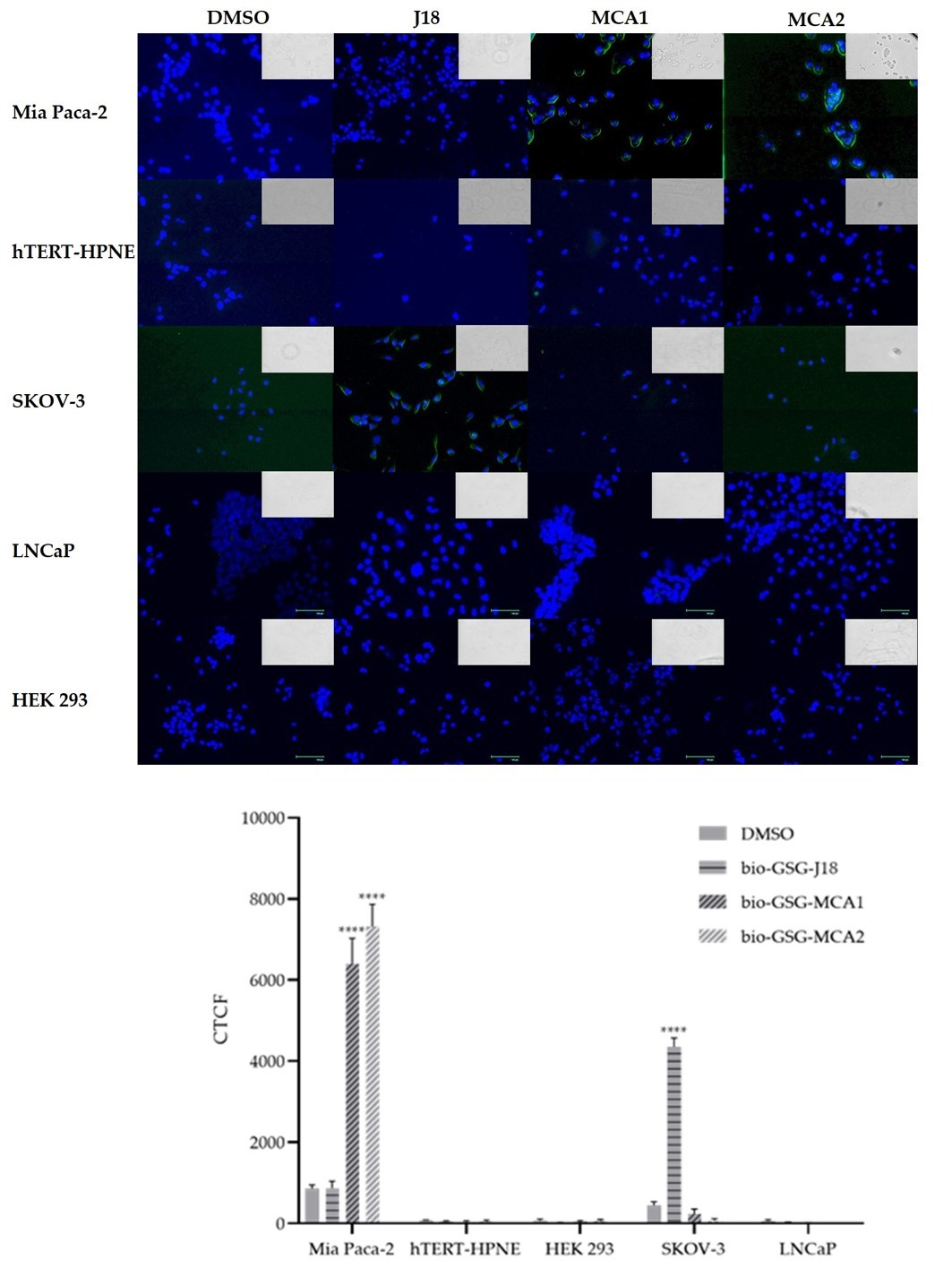
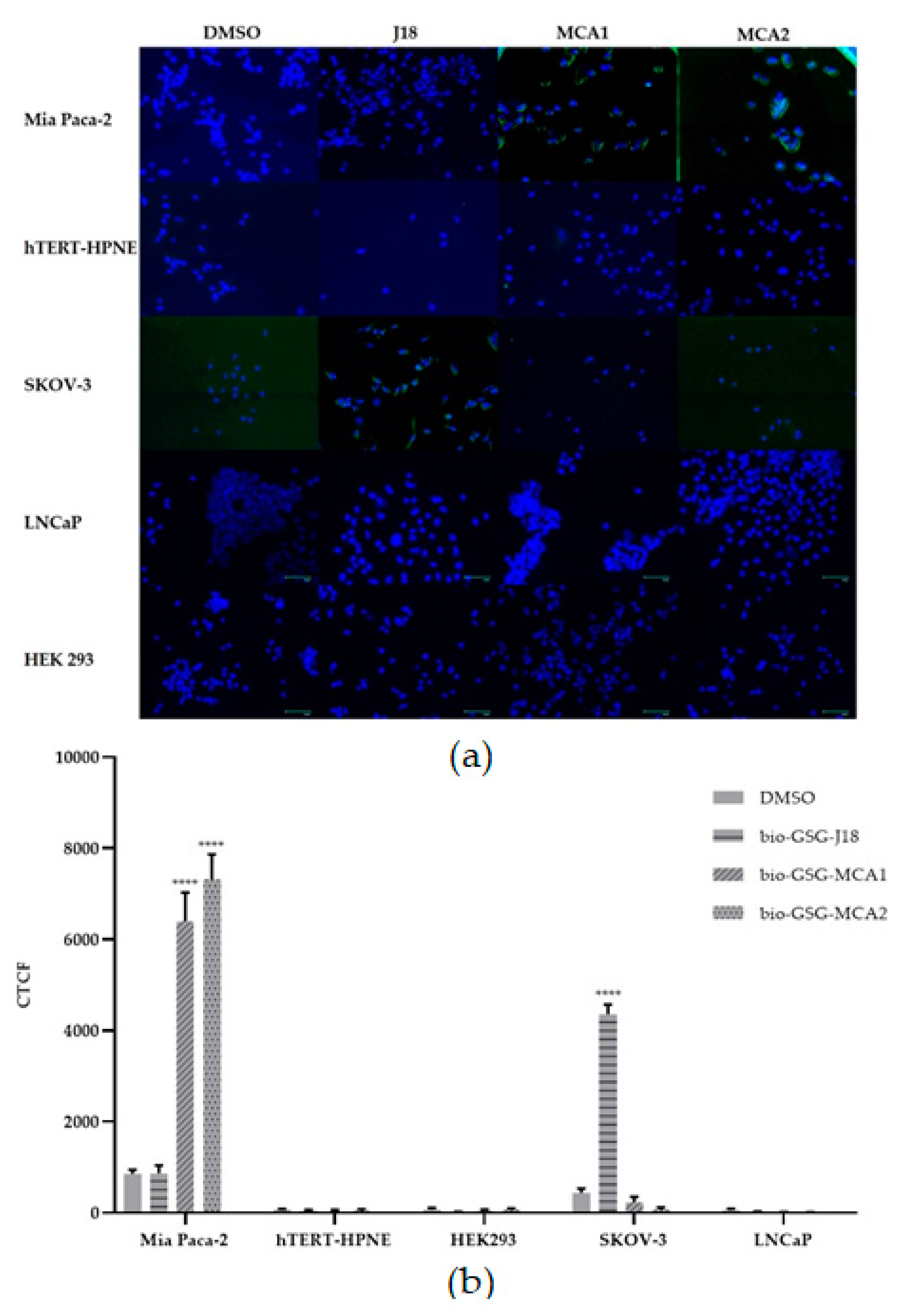
3.2. Fluorescent Microscopy
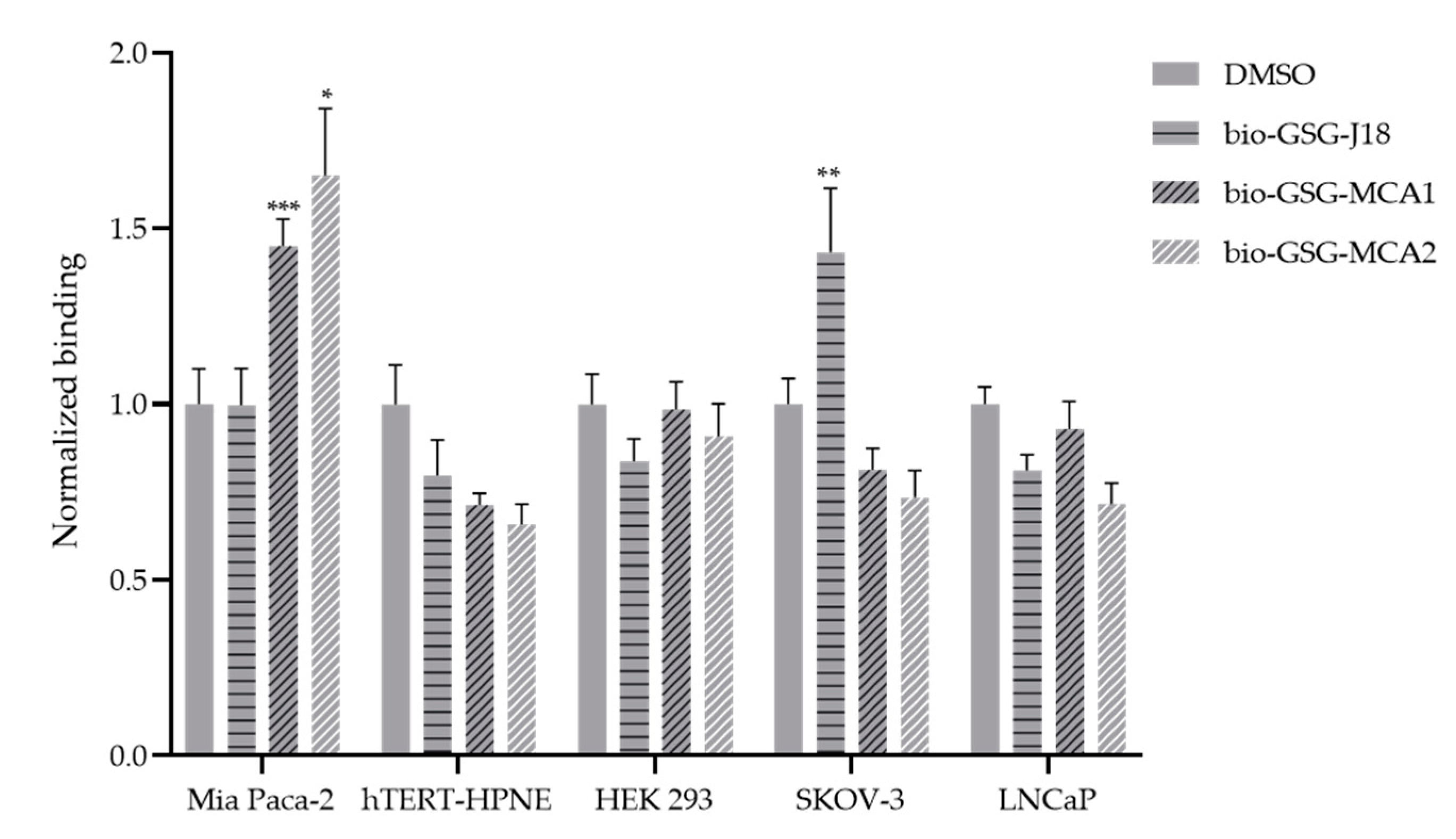
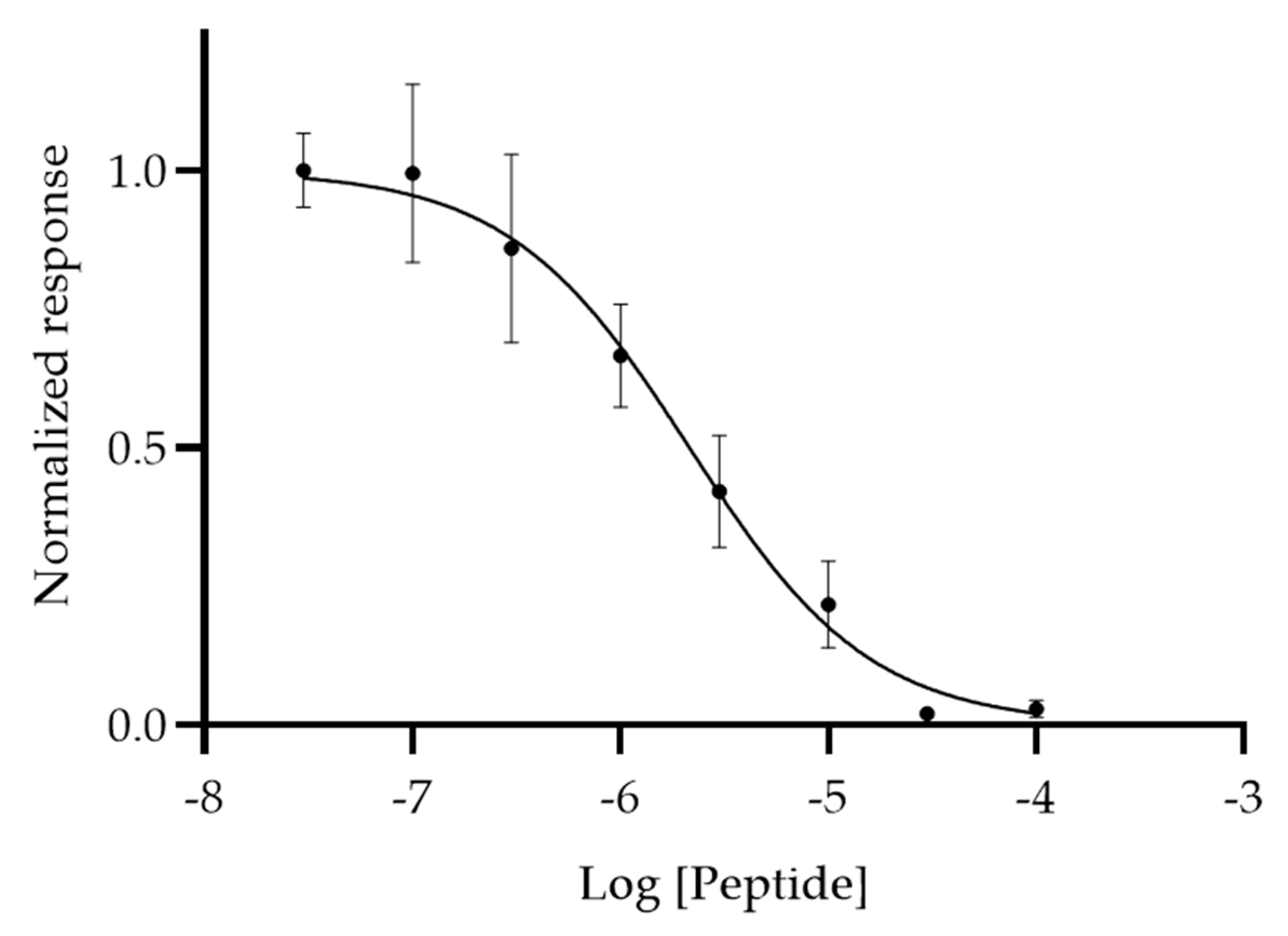
3.3. Modified ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, A.; Herman, J.; Schulick, R.; Hruban, R.H.; Goggins, M. Pancreatic cancer. Lancet 2011, 378, 607–620. [Google Scholar] [CrossRef]
- Chari, S.T. Detecting early pancreatic cancer: Problems and prospects. Semin. Oncol. 2007, 34, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrianifahanana, M.; Moniaux, N.; Schmied, B.M.; Ringel, J.; Friess, H.; Hollingsworth, M.A.; Buchler, M.W.; Aubert, J.P.; Batra, S.K. Mucin (MUC) gene expression in human pancreatic adenocarcinoma and chronic pancreatitis: A potential role of MUC4 as a tumor marker of diagnostic significance. Clin. Cancer Res. 2001, 7, 4033–4040. [Google Scholar]
- Dimastromatteo, J.; Houghton, J.L.; Lewis, J.S.; Kelly, K.A. Challenges of Pancreatic Cancer. Cancer J. 2015, 21, 188–193. [Google Scholar] [CrossRef] [Green Version]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef]
- Staal, B.; Liu, Y.; Barnett, D.; Hsueh, P.; He, Z.; Gao, C.; Partyka, K.; Hurd, M.W.; Singhi, A.D.; Drake, R.R.; et al. The sTRA Plasma Biomarker: Blinded Validation of Improved Accuracy Over CA19-9 in Pancreatic Cancer Diagnosis. Clin. Cancer Res. 2019, 25, 2745–2754. [Google Scholar] [CrossRef] [Green Version]
- van Manen, L.; Groen, J.V.; Putter, H.; Vahrmeijer, A.L.; Swijnenburg, R.-J.; Bonsing, B.A.; Mieog, J.S.D. Elevated CEA and CA19-9 serum levels independently predict advanced pancreatic cancer at diagnosis. Biomarkers 2020, 25, 186–193. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, D.P.; Sandanayake, N.S.; Jenkinson, C.; Gentry-Maharaj, A.; Apostolidou, S.; Fourkala, E.-O.; Camuzeaux, S.; Blyuss, O.; Gunu, R.; Dawnay, A.; et al. Serum CA19-9 Is Significantly Upregulated up to 2 Years before Diagnosis with Pancreatic Cancer: Implications for Early Disease Detection. Clin. Cancer Res. 2015, 21, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Ballehaninna, U.K.; Chamberlain, R.S. The clinical utility of serum CA 19-9 in the diagnosis, prognosis and management of pancreatic adenocarcinoma: An evidence based appraisal. J. Gastrointest. Oncol. 2012, 3, 105–119. [Google Scholar]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: Affinity purification of target genes. Gene 1988, 73, 305–318. [Google Scholar] [CrossRef]
- Scott, J.K.; Smith, G.P. Searching for peptide ligands with an epitope library. Science 1990, 249, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.P.; Scott, J.K. Libraries of peptides and proteins displayed on filamentous phage. Methods Enzymol. 1993, 217, 228–257. [Google Scholar] [PubMed]
- Asar, M.; Newton-Northup, J.; Deutscher, S.; Soendergaard, M. Ovarian Cancer Targeting Phage for In Vivo Near-Infrared Optical Imaging. Diagnostics 2019, 9, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soendergaard, M.; Newton-Northup, J.R.; Deutscher, S.L. In vitro high throughput phage display selection of ovarian cancer avid phage clones for near-infrared optical imaging. Comb. Chem. High Throughput Screen. 2014, 17, 859–867. [Google Scholar] [CrossRef]
- Soendergaard, M.; Newton-Northup, J.R.; Deutscher, S.L. In vivo phage display selection of an ovarian cancer targeting peptide for SPECT/CT imaging. Am. J. Nucl. Med. Mol. Imaging 2014, 4, 561–570. [Google Scholar]
- Deutscher, S.L.; Figueroa, S.D.; Kumar, S.R. In-labeled KCCYSL peptide as an imaging probe for ErbB-2-expressing ovarian carcinomas. J. Labelled Comp. Radiopharm. 2009, 52, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Agris, P.F.; Marchbank, M.T.; Newman, W.; Guenther, R.; Ingram, P.; Swallow, J.; Mucha, P.; Szyk, A.; Rekowski, P.; Peletskaya, E.; et al. Experimental models of protein-RNA interaction: Isolation and analyses of tRNA(Phe) and U1 snRNA-binding peptides from bacteriophage display libraries. J. Protein Chem. 1999, 18, 425–435. [Google Scholar] [CrossRef]
- Kumar, S.R.; Deutscher, S.L. Galectin-3-Targeting Peptides as Novel In Vivo tumor-Imaging and Targeting Agents. In Tumor Microvasculature Environment AACR Meeting; AACR: Florence, Italy, 2006. [Google Scholar]
- Kumar, S.R.; Deutscher, S.L. 111In-labeled galectin-3-targeting peptide as a SPECT agent for imaging breast tumors. J. Nucl. Med. 2008, 49, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Kelly, K.A.; Jones, D.A. Isolation of a colon tumor specific binding peptide using phage display selection. Neoplasia 2003, 5, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Reubi, J.C.; Maecke, H.R. Peptide-based probes for cancer imaging. J. Nucl. Med. 2008, 49, 1735–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behr, T.M.; Gotthardt, M.; Barth, A.; Behe, M. Imaging tumors with peptide-based radioligands. Q. J. Nucl. Med. 2001, 45, 189–200. [Google Scholar] [PubMed]
- Karasseva, N.G.; Glinsky, V.V.; Chen, N.X.; Komatireddy, R.; Quinn, T.P. Identification and characterization of peptides that bind human ErbB-2 selected from a bacteriophage display library. J. Protein Chem. 2002, 21, 287–296. [Google Scholar] [CrossRef]
- Larimer, B.; Thomas, W.; Smith, G.; Deutscher, S. Affinity Maturation of an ERBB2-Targeted SPECT Imaging Peptide by In Vivo Phage Display. Mol. Imaging Biol. 2014, 1–10. [Google Scholar] [CrossRef]
- Peletskaya, E.N.; Glinsky, G.; Deutscher, S.L.; Quinn, T.P. Identification of peptide sequences that bind the Thomsen-Friedenreich cancer-associated glycoantigen from bacteriophage peptide display libraries. Mol. Divers. 1996, 2, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Gallazzi, F.A.; Quinn, T.P.; Deutscher, S.L. (64)Cu-labeled peptide for PET of breast carcinomas expressing the Thomsen-Friedenreich carbohydrate antigen. J. Nucl. Med. 2011, 52, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Arap, W.; Kolonin, M.G.; Trepel, M.; Lahdenranta, J.; Cardo-Vila, M.; Giordano, R.J.; Mintz, P.J.; Ardelt, P.U.; Yao, V.J.; Vidal, C.I.; et al. Steps toward mapping the human vasculature by phage display. Nat. Med. 2002, 8, 121–127. [Google Scholar] [CrossRef]
- Yao, V.J.; Ozawa, M.G.; Trepel, M.; Arap, W.; McDonald, D.M.; Pasqualini, R. Targeting pancreatic islets with phage display assisted by laser pressure catapult microdissection. Am. J. Pathol. 2005, 166, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.P. Filamentous phages as cloning vectors. Biotechnology 1988, 10, 61–83. [Google Scholar]
- G. P. Smith Lab. Homepage. Available online: www.biosci.missouri.edu/smithGP/ (accessed on 6 January 2019).
- Newton, J.R.; Kelly, K.A.; Mahmood, U.; Weissleder, R.; Deutscher, S.L. In vivo selection of phage for the optical imaging of PC-3 human prostate carcinoma in mice. Neoplasia 2006, 8, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Couet, J.; Li, S.; Okamoto, T.; Ikezu, T.; Lisanti, M.P. Identification of peptide and protein ligands for the caveolin-scaffolding domain. Implications for the interaction of caveolin with caveolae-associated proteins. J. Biol. Chem. 1997, 272, 6525–6533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, J. Phage Display to Identify Peptides Binding to or Penetrating the Mouse Zona Pellucida. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 1999. [Google Scholar]
- Landon, L.A.; Harden, W.; Illy, C.; Deutscher, S.L. High-throughput fluorescence spectroscopic analysis of affinity of peptides displayed on bacteriophage. Anal. Biochem. 2004, 331, 60–67. [Google Scholar]
- Landon, L.A.; Peletskaya, E.N.; Glinsky, V.V.; Karasseva, N.; Quinn, T.P.; Deutscher, S.L. Combinatorial evolution of high-affinity peptides that bind to the Thomsen-Friedenreich carcinoma antigen. J. Protein Chem. 2003, 22, 193–204. [Google Scholar] [CrossRef]
- Li, F.; Dluzewski, A.; Coley, A.M.; Thomas, A.; Tilley, L.; Anders, R.F.; Foley, M. Phage-displayed Peptides Bind to the Malarial Protein Apical Membrane Antigen-1 and Inhibit the Merozoite Invasion of Host Erythrocytes. J. Biol. Chem. 2002, 277, 50303–50310. [Google Scholar] [CrossRef] [Green Version]
- Deutscher, S.L. Phage display in molecular imaging and diagnosis of cancer. Chem. Rev. 2010, 110, 3196–3211. [Google Scholar] [CrossRef] [Green Version]
- Glinsky, V.V.; Glinsky, G.V.; Rittenhouse-Olson, K.; Huflejt, M.E.; Glinskii, O.V.; Deutscher, S.L.; Quinn, T.P. The role of Thomsen-Friedenreich antigen in adhesion of human breast and prostate cancer cells to the endothelium. Cancer Res. 2001, 61, 4851–4857. [Google Scholar]
- Glinsky, V.V.; Huflejt, M.E.; Glinsky, G.V.; Deutscher, S.L.; Quinn, T.P. Effects of Thomsen-Friedenreich antigen-specific peptide P-30 on beta-galactoside-mediated homotypic aggregation and adhesion to the endothelium of MDA-MB-435 human breast carcinoma cells. Cancer Res. 2000, 60, 2584–2588. [Google Scholar]
- Matarrese, P.; Fusco, O.; Tinari, N.; Natoli, C.; Liu, F.T.; Semeraro, M.L.; Malorni, W.; Iacobelli, S. Galectin-3 overexpression protects from apoptosis by improving cell adhesion properties. Int. J. Cancer 2000, 85, 545–554. [Google Scholar] [CrossRef]
- Choi, J.H.; Choi, K.C.; Auersperg, N.; Leung, P.C. Gonadotropins upregulate the epidermal growth factor receptor through activation of mitogen-activated protein kinases and phosphatidyl-inositol-3-kinase in human ovarian surface epithelial cells. Endocr. Relat. Cancer 2005, 12, 407–421. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.K.; Kumar, S.; Cannon, A.; Hall, B.; Bhatia, R.; Nasser, M.W.; Mahapatra, S.; Batra, S.K.; Jain, M. MUC4 mucin- a therapeutic target for pancreatic ductal adenocarcinoma. Expert Opin. Ther. Targets 2017, 21, 657–669. [Google Scholar] [CrossRef]
- Ohmine, K.; Kawaguchi, K.; Ohtsuki, S.; Motoi, F.; Ohtsuka, H.; Kamiie, J.; Abe, T.; Unno, M.; Terasaki, T. Quantitative Targeted Proteomics of Pancreatic Cancer: Deoxycytidine Kinase Protein Level Correlates to Progression-Free Survival of Patients Receiving Gemcitabine Treatment. Mol. Pharm. 2015, 12, 3282–3291. [Google Scholar] [CrossRef]
- Giovannetti, E.; Del Tacca, M.; Mey, V.; Funel, N.; Nannizzi, S.; Ricci, S.; Orlandini, C.; Boggi, U.; Campani, D.; Del Chiaro, M.; et al. Transcription analysis of human equilibrative nucleoside transporter-1 predicts survival in pancreas cancer patients treated with gemcitabine. Cancer Res. 2006, 66, 3928–3935. [Google Scholar] [CrossRef] [Green Version]
- Kelly, K.A.; Nahrendorf, M.; Yu, A.M.; Reynolds, F.; Weissleder, R. In vivo phage display selection yields atherosclerotic plaque targeted peptides for imaging. Mol. Imaging Biol. 2006, 8, 201–207. [Google Scholar] [CrossRef]
- Kumar, S.R.; Gallazzi, F.A.; Ferdani, R.; Anderson, C.J.; Quinn, T.P.; Deutscher, S.L. In vitro and in vivo evaluation of Cu-radiolabeled KCCYSL peptides for targeting epidermal growth factor receptor-2 in breast carcinomas. Cancer Biother. Radiopharm. 2010, 25, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Deutscher, S.L.; Figueroa, S.D.; Kumar, S.R. Tumor targeting and SPECT imaging properties of an (111)In-labeled galectin-3 binding peptide in prostate carcinoma. Nucl. Med. Biol. 2009, 36, 137–146. [Google Scholar] [CrossRef]
- Reubi, J.C. Peptide receptors as molecular targets for cancer diagnosis and therapy. Endocr. Rev. 2003, 24, 389–427. [Google Scholar] [CrossRef] [Green Version]
- Saw, P.E.; Song, E.-W. Phage display screening of therapeutic peptide for cancer targeting and therapy. Protein Cell 2019, 10, 787–807. [Google Scholar] [CrossRef] [Green Version]
- Kokoszka, M.E.; Kall, S.L.; Khosla, S.; McGinnis, J.E.; Lavie, A.; Kay, B.K. Identification of two distinct peptide-binding pockets in the SH3 domain of human mixed-lineage kinase 3. J. Biol. Chem. 2018, 293, 13553–13565. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Serohijos, A.W.; Newton, G.; Tassone, G.; Wang, Z.; Sgroi, D.C.; Dokholyan, N.V.; Basilion, J.P. Identification and rational redesign of peptide ligands to CRIP1, a novel biomarker for cancers. PLoS Comput. Biol. 2008, 4. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Log2 (pos/neg) |
|---|---|
| MCA1 | 6.03 |
| MCA2 | 3.73 |
| MCA3 | −1.86 |
| MCA4 | −3.61 |
| MCA5 | −2.49 |
| MCA6 | −3.98 |
| MCA7 | −3.77 |
| MCA8 | −3.90 |
| MCA9 | −3.97 |
| MCA10 | −3.43 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asar, M.C.; Franco, A.; Soendergaard, M. Phage Display Selection, Identification, and Characterization of Novel Pancreatic Cancer Targeting Peptides. Biomolecules 2020, 10, 714. https://doi.org/10.3390/biom10050714
Asar MC, Franco A, Soendergaard M. Phage Display Selection, Identification, and Characterization of Novel Pancreatic Cancer Targeting Peptides. Biomolecules. 2020; 10(5):714. https://doi.org/10.3390/biom10050714
Chicago/Turabian StyleAsar, Mallika C., April Franco, and Mette Soendergaard. 2020. "Phage Display Selection, Identification, and Characterization of Novel Pancreatic Cancer Targeting Peptides" Biomolecules 10, no. 5: 714. https://doi.org/10.3390/biom10050714