Ameliorative Effect of Gum Acacia on Hookah Smoke-Induced Testicular Impairment in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Exposure to HS
2.3. Blood Collection and Tissue Preparation
2.4. Biochemical Estimations in Plasma and Urine
2.5. Biochemical Measurements in Testicular Homogenates
2.6. Histopathology of Testicular Sections
2.7. Superoxide Dismutase (SOD) Immunohistochemistry
2.8. StAR Western Blotting
2.9. Drugs, Chemicals, and Kits
2.10. Statistical Analysis
3. Results
3.1. Effect of HS on Body and Testicular Weights
3.2. Effect of HS on the Levels of Some Plasma Hormones, Inhibin B, and Androgen-Binding Protein
3.3. Effect of HS on the Levels of Urine Cotinine
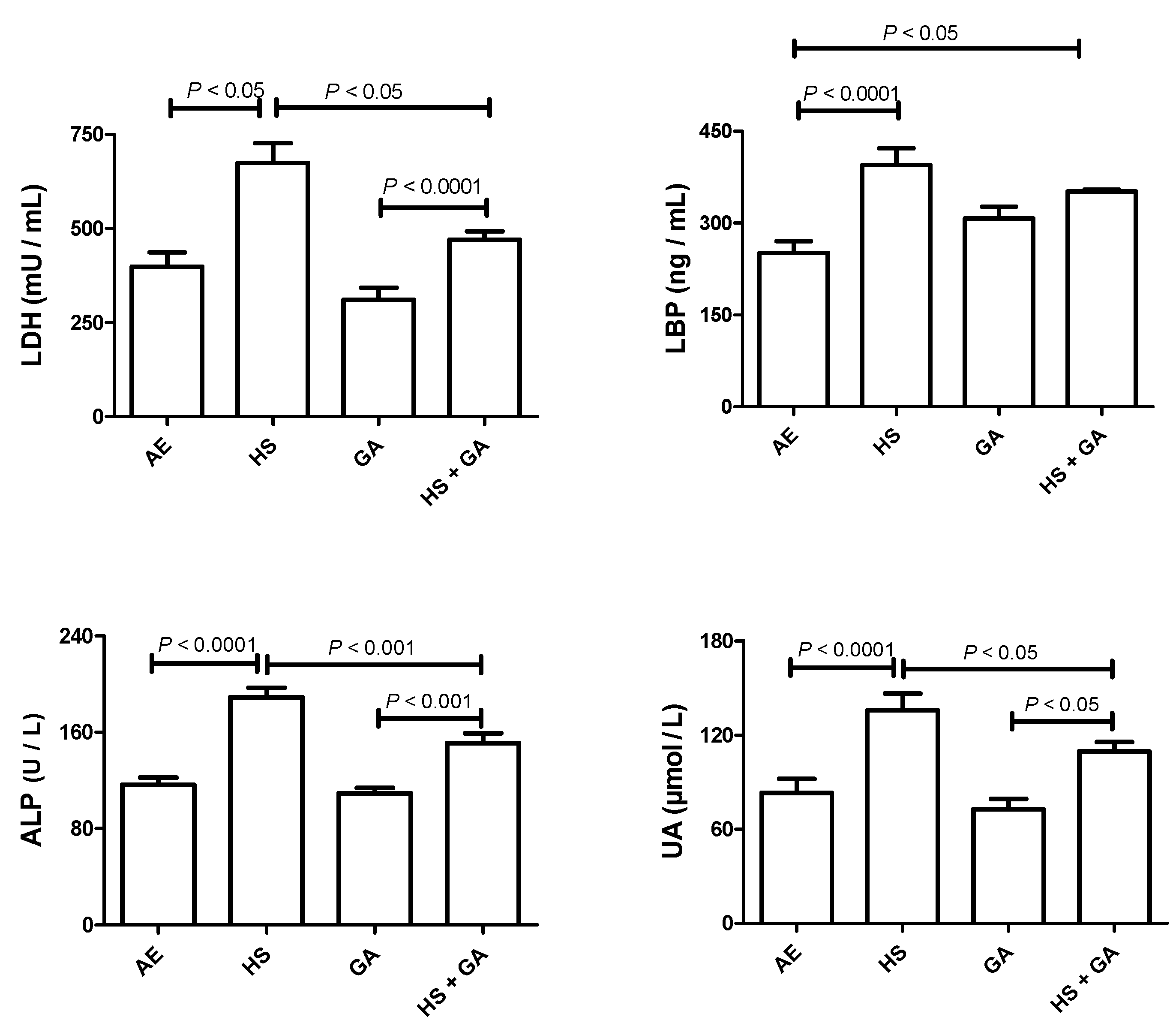
3.4. Effect of HS on the Concentrations of Some Plasma Analytes
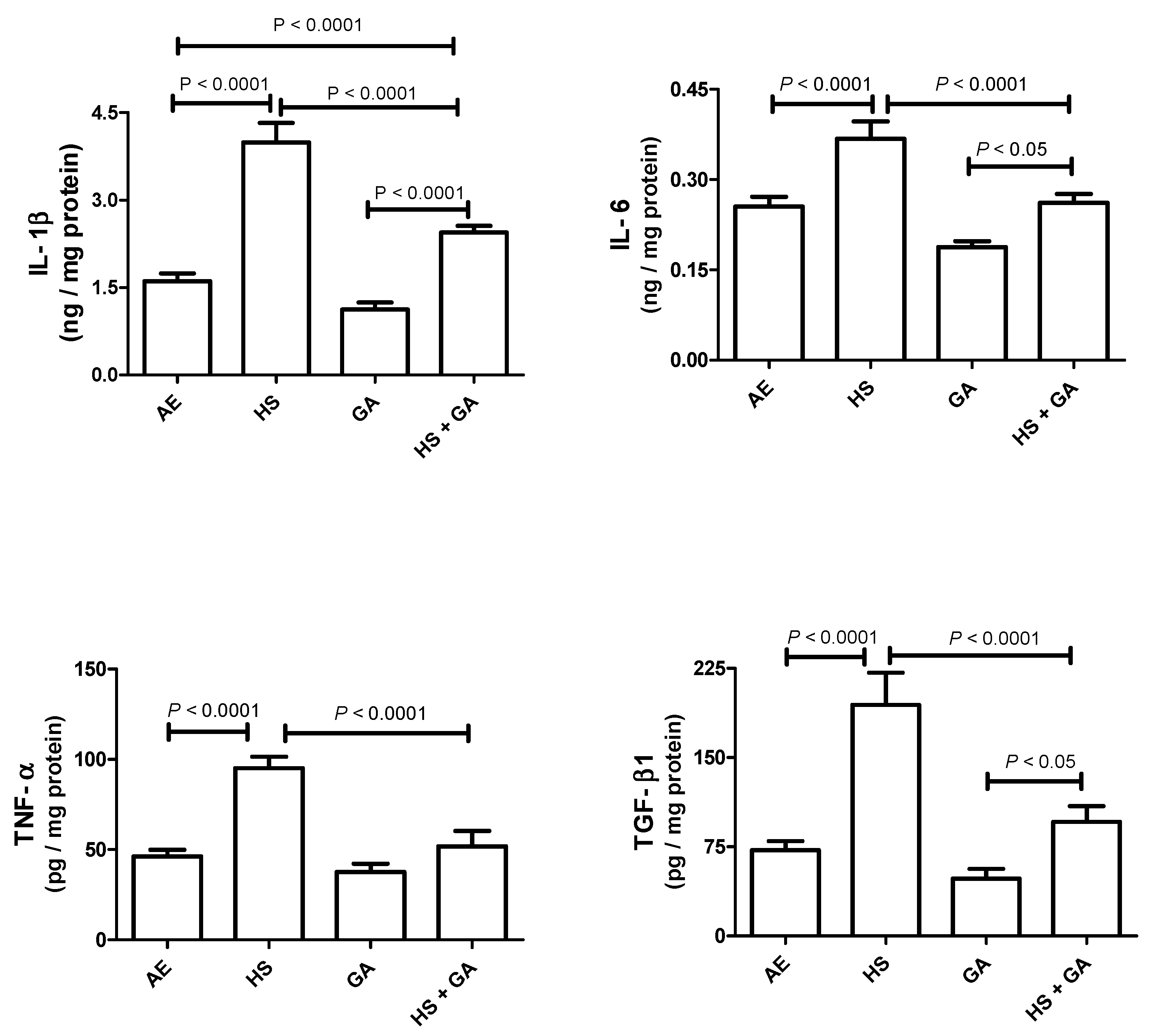
3.5. Effect of HS on Some Cytokines in the Testis Homogenate
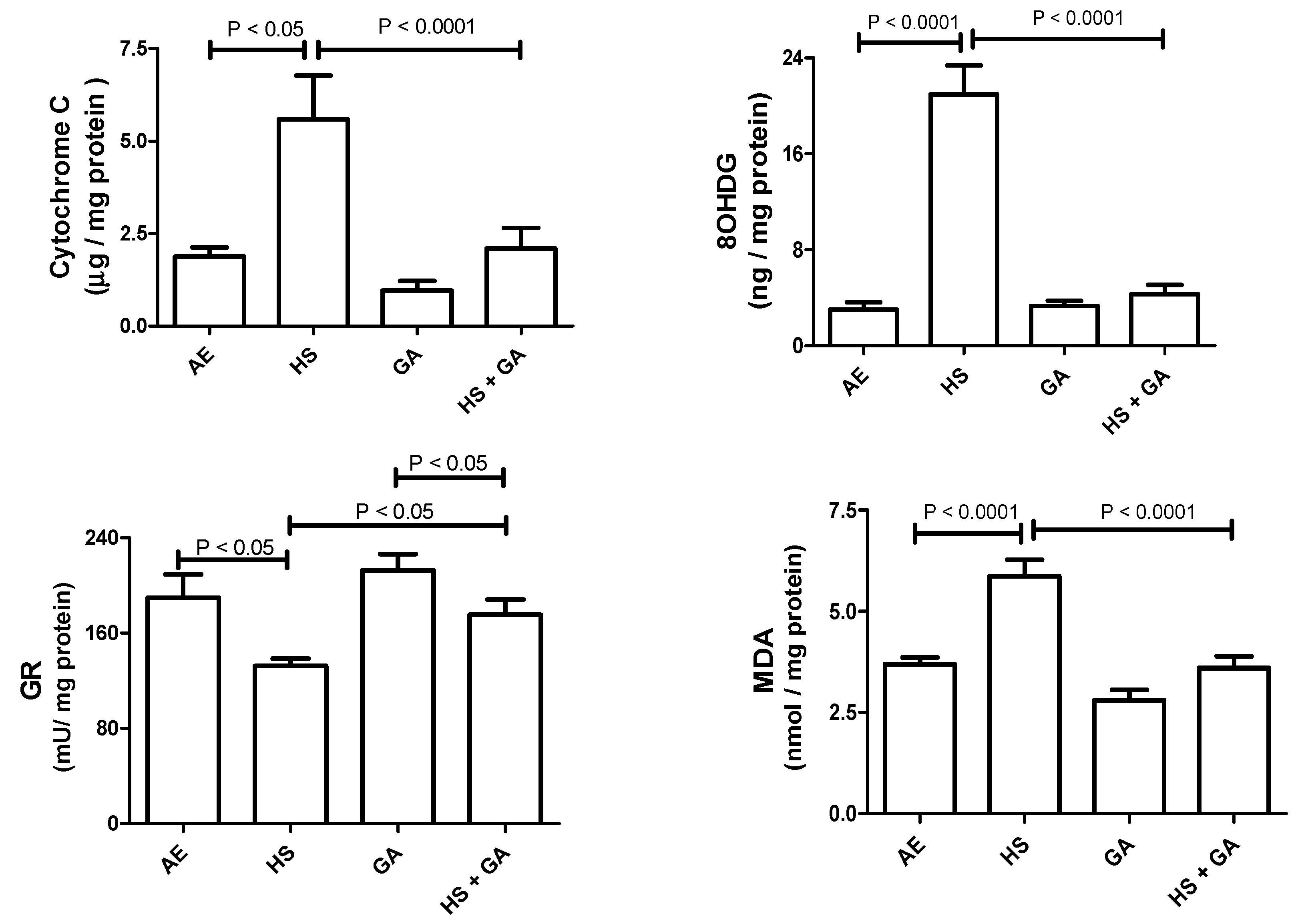
3.6. Effect of HS on Some Antioxidants in the Testis Homogenate
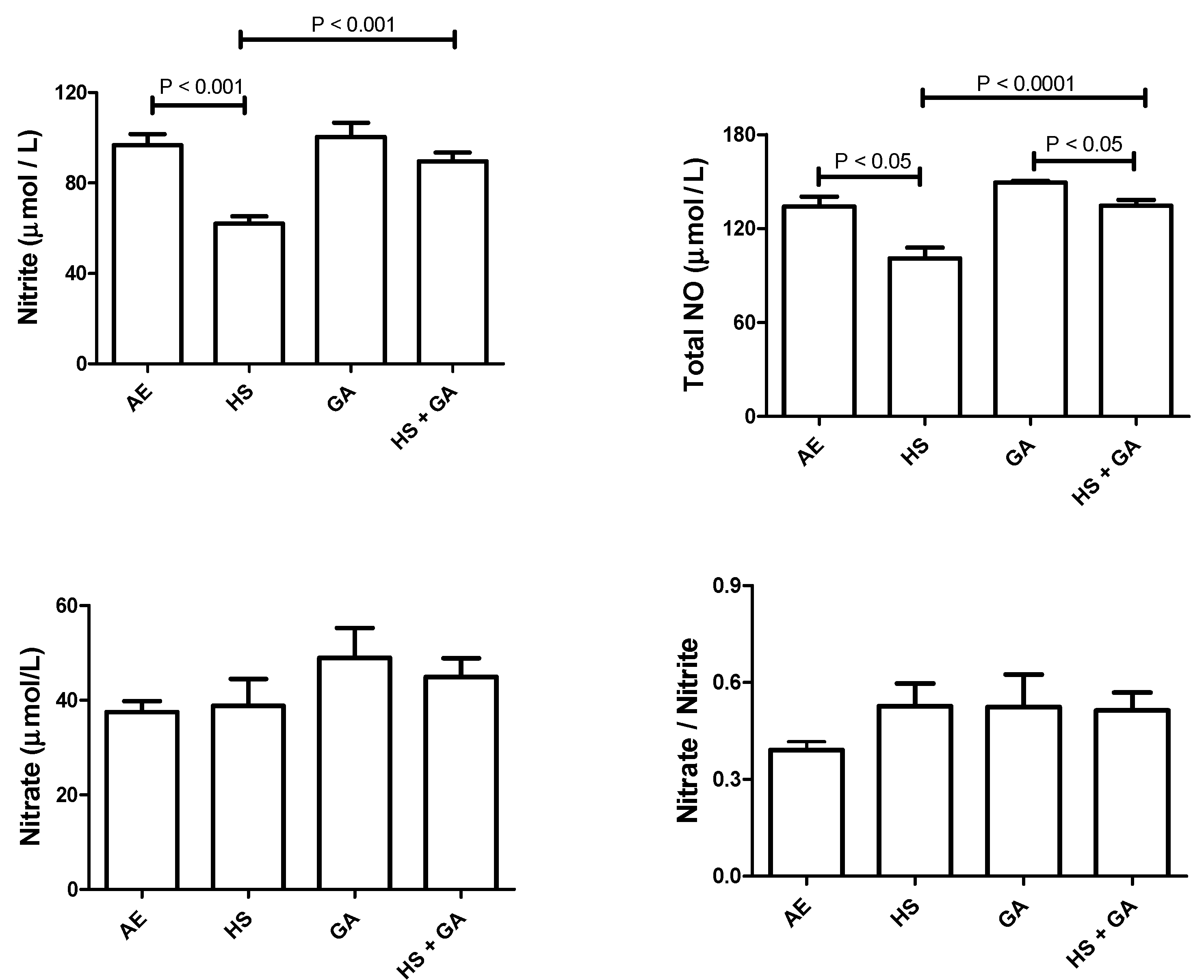
3.7. Effect of HS on Nitrosative Stress in the Testis of HS-Exposed Mice
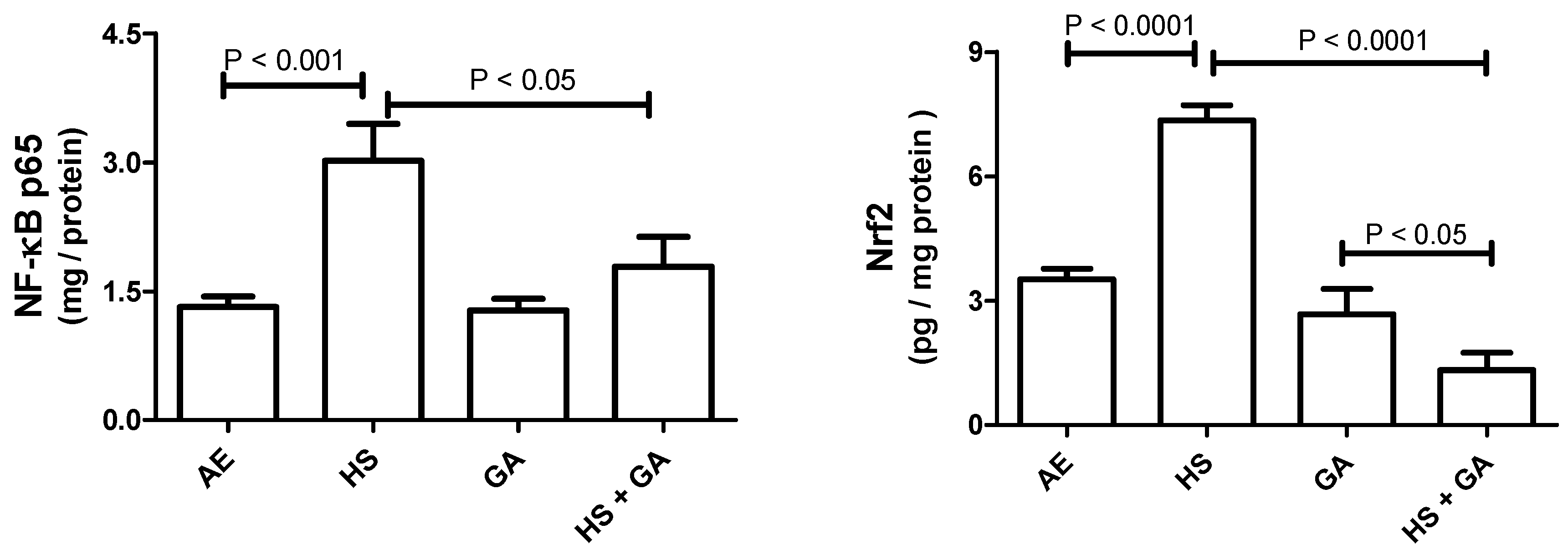
3.8. Effect of HS on NF-ĸB p65 and Nrf2 in Testicular Homogenate
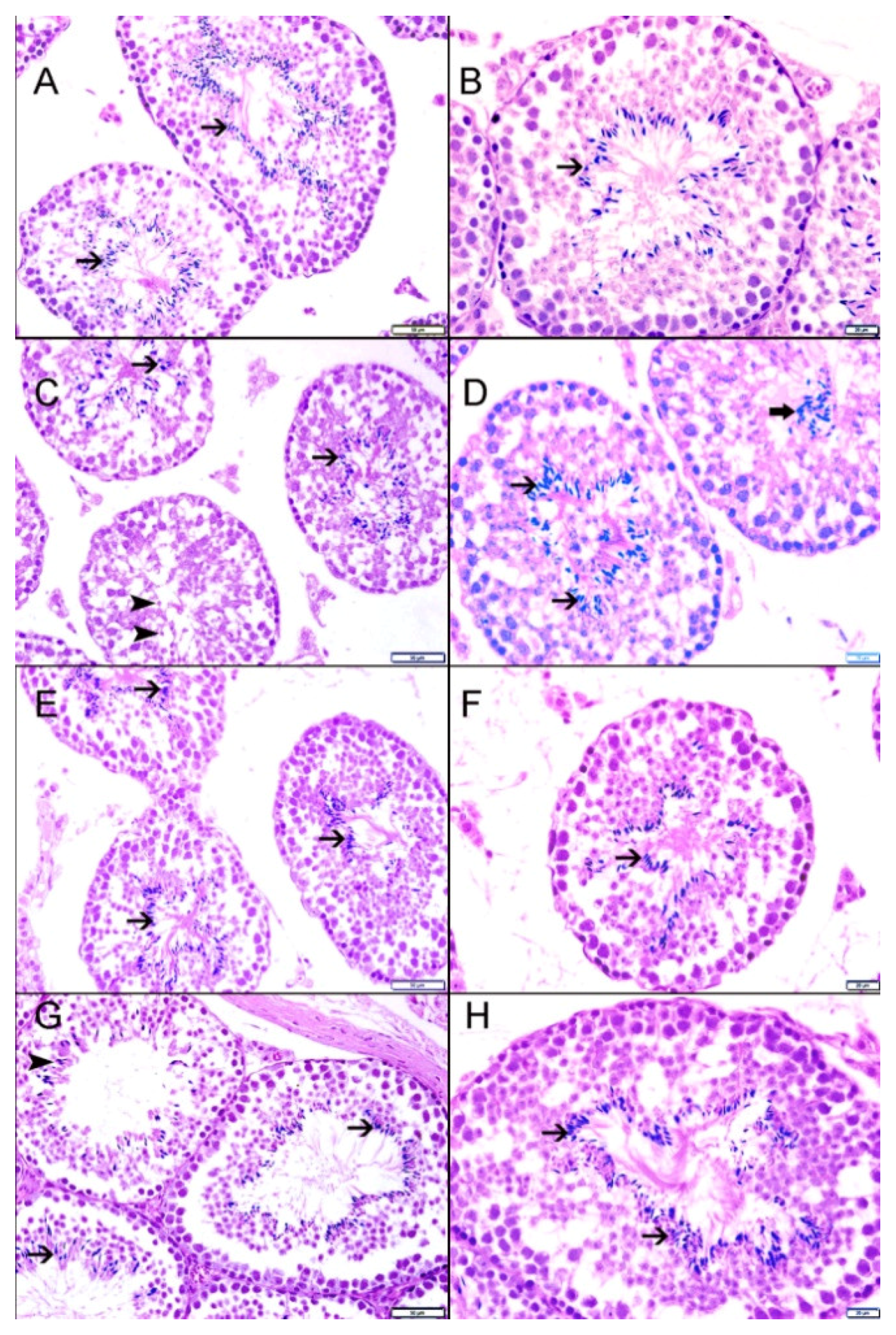
3.9. Light Microscopic Histopathology
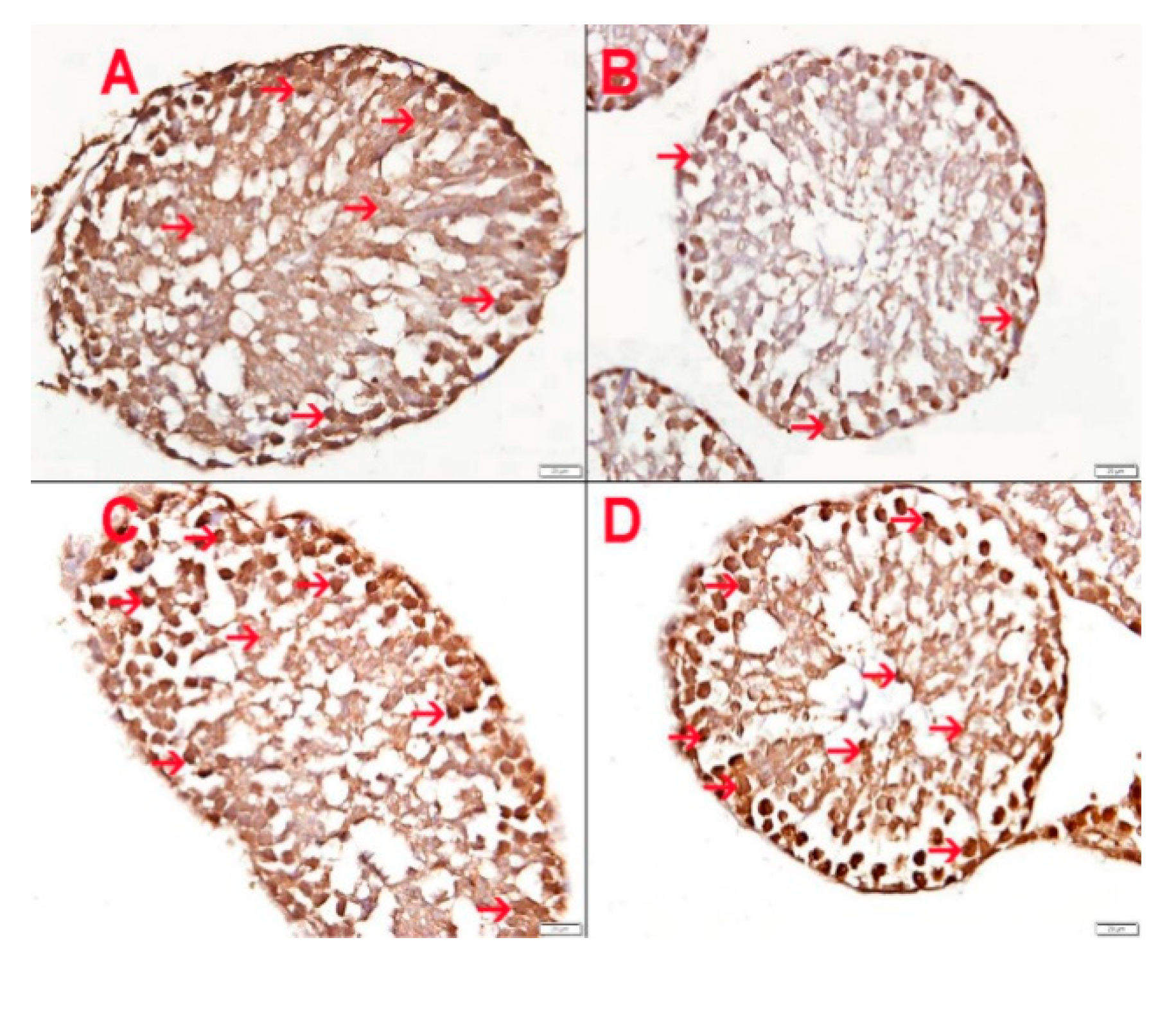
3.10. Effect of HS Exposure on Testicular SOD Immunohistochemistry
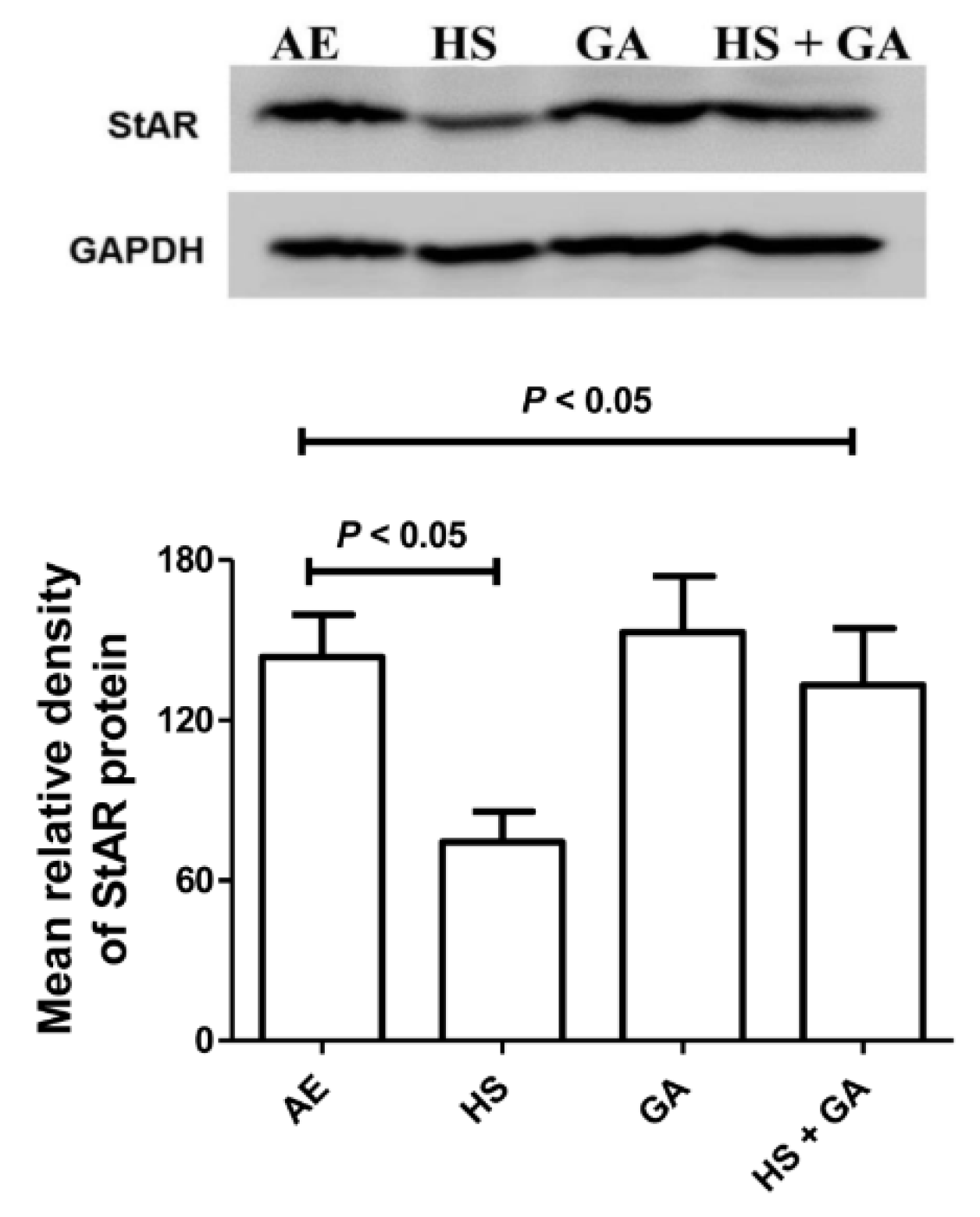
3.11. Effect of HS Exposure on Testicular StAR Protein
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Maziak, W.; Taleb, Z.B.; Bahelah, R.; Islam, F.; Jaber, R.; Auf, R.; Salloum, R.G. The global epidemiology of waterpipe smoking. Tob. Control 2015, 24 (Suppl. 1), i3–i12. [Google Scholar] [CrossRef]
- Salloum, R.G.; Haider, M.R.; Barnett, T.E.; Guo, Y.; Getz, K.R.; Thrasher, J.; Maziak, W. Water- pipe tobacco smoking and susceptibility to cigarette smoking among young Adults in the United States, 2012–2013. Prev. Chronic. Dis. 2016, 13, E24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, T.; Arrazola, R.A.; Corey, C.G.; Husten, C.G.; Neff, L.J.; Homa, D.M. Tobacco use among Middle and High School students -United States, 2011–2015. MMWR Morb. Mortal Wkly. Rep. 2016, 65, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Kulak, J.A.; Saddleson, M.L.; Travers, M.J.; Bansal-Travers, M.; Homish, G.G.; Mahoney, M.C.; Giovino, G.A. Examining hookah as an introduction to nicotine products among college students. Subst. Use Misuse 2018, 53, 1869–1877. [Google Scholar] [CrossRef] [PubMed]
- Primack, B.A.; Carroll, M.V.; Weiss, P.M.; Shihadeh, A.L.; Shensa, A.; Farley, S.T.; Fine, M.J.; Eissenberg, T.; Nayak, S. Systematic review and meta-analysis of inhaled toxicants from waterpipe and cigarette smoking. Public Health Rep. 2016, 131, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahelah, R.; DiFranza, J.R.; Ward, K.D.; Eissenberg, T.; Fouad, F.M.; Taleb, Z.B.; Jaber, R.; Maziak, W. Waterpipe smoking patterns and symptoms of nicotine dependence: The waterpipe dependence in Lebanese Youth Study. Addict. Behav. 2017, 74, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Onor, I.O.; Stirling, D.L.; Williams, S.R.; Bediako, D.; Borghol, A.; Harris, M.B.; Darensburg, T.B.; Clay, S.D.; Okpechi, S.C.; Sarpong, D.F. Clinical effects of cigarette smoking: Epidemiologic impact and review of pharmacotherapy options. Int. J. Environ. Res. Public Health 2017, 14, 1147. [Google Scholar] [CrossRef] [Green Version]
- Waziry, R.; Jawad, M.; Ballout, R.A.; Al Akel, M.; Akl, E.A. The effects of water pipe tobacco smoking on health outcomes: An updated systematic review and meta-analysis. Int. J. Epidemiol. 2017, 46, 32–43. [Google Scholar]
- Muzammil, D.S.; Al Rethaiaa, A.S.; Al Mutairi, A.S.; Al Rashidi, T.H.; Al Rasheedi, H.A.; Al Rasheedi, S.A. Prevalence and perception of shisha smoking among university students: A cross-sectional study. J. Int. Soc. Prev. Commun. Dent. 2019, 9, 275–281. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Ali, B.H. Chronic exposure to water-pipe smoke induces alveolar enlargement, DNA damage and impairment of lung function. Cell Physiol. Biochem. 2016, 38, 982–992. [Google Scholar] [CrossRef]
- Nemmar, A.; Al Salam, S.; Beegam, S.; Yuvaraju, P.; Oulhaj, A.; Ali, B.H. Water pipe smoke exposure—Induced circulatory disturbances in mice, and the influence of betaine supplementation thereon. Cell Physiol. Biochem. 2017, 41, 1098–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, B.H.; Al Balushi, K.A.; Ashique, M.; Shalaby, A.; Al Kindi, M.A.; Adham, S.A.; Karaca, T.; Beegam, S.; Yuvaraju, P.; Nemmar, A. Chronic water-pipe smoke exposure induces injurious effects to reproductive system in male mice. Front Physiol. 2017, 8, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Ali, B.H. Gum arabic ameliorates impaired coagulation and cardiotoxicity induced by water-pipe smoke exposure in mice. Front Physiol. 2019, 10, 53. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Al Salam, S.; Adham, S.A.; Al Balushi, K.; Al Za’abi, M.; Beegam, S. Testicular toxicity of water pipe smoke exposure in mice and the effect of treatment with nootkatone thereon. Oxid. Med. Cell Longev. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calame, W.; Weseler, A.R.; Viebke, C.; Flynn, C.; Siemensma, A.D. Gum arabic establishes prebiotic functionality in healthy human volunteers in a dose-dependent manner. Br. J. Nutr. 2008, 100, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Ziada, A.; Blunden, G. Biological effects of gum arabic: A review of some recent research. Food Chem. Toxicol. 2009, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Al Husseni, I.; Beegam, S.; Al Shukaili, A.; Nemmar, A.; Schierling, S. Effect of gum arabic on oxidative stress and inflammation in adenine-induced chronic renal failure in rats. PLoS ONE 2013, 8, e55242. [Google Scholar] [CrossRef] [Green Version]
- Kamal, E.; Kaddam, L.A.; Dahawi, M.; Osman, M.; Salih, M.A.; Alagib, A.; Saeed, A. Gum arabic fibers decreased inflammatory markers and disease severity score among rheumatoid arthritis patients, phase II trial. Int. J. Rheumatol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Babiker, R.; Merghani, T.H.; Elmusharaf, K.; Badi, R.M.; Lang, F.; Saeed, A.M. Effects of gum arabic ingestion on body mass index and body fat percentage in healthy adult females: Two-arm randomized, placebo controlled, double-blind trial. Nutr. J. 2012, 11, 111. [Google Scholar] [CrossRef]
- Kaddam, L.; Fadl-Elmula, I.; Eisawi, O.A.; Abdelrazig, H.A.; Saeed, A.M. Acacia senegal (Gum Arabic) supplementation modulate lipid profile and ameliorated dyslipidemia among sickle cell anemia patients. J. Lipids. 2019, 3129461. [Google Scholar] [CrossRef] [Green Version]
- Nemmar, A.; Raza, H.; Yuvaraju, P.; Beegam, S.; John, A.; Yasin, J.; Hameed, S.R.; Adeghate, E.; Ali, B.H. Nose- only water- pipe smoking effects on airway resistance, inflammation, and oxidative stress in mice. J. Appl. Physiol. 2013, 115, 1316–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Ali, B.H. Chronic exposure to water-pipe smoke induces cardiovascular dysfunction in mice. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H329–H339. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Adham, S.A.; Al Balushi, K.A.; Shalaby, A.; Waly, M.I.; Manoj, P.; Beegam, S.; Yuvaraju, P.; Nemmar, A. Reproductive toxicity to male mice of nose only exposure to water- pipe smoke. Cell Physiol. Biochem 2015, 35, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Ma, Q.; Yuan, J.; Hu, X.; Tan, Q.; Zhang, Z.; Wang, L.; Xu, X. The effects of di-2-ethylhexyl phthalate on testicular ultrastructure and hormone-regulated gene expression in male rats. Toxicol. Res. 2018, 7, 408–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemmar, A.; Yuvaraju, P.; Beegam, S.; John, A.; Raza, H.; Ali, B.H. Cardiovascular effects of nose-only water-pipe smoking exposure in mice. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H740–H746. [Google Scholar] [CrossRef]
- Marom-Haham, L.; Shulman, A. Cigarette smoking and hormones. Curr Opin Obstet. Gynecol. 2016, 28, 230–235. [Google Scholar] [CrossRef]
- Chan, Y.L.; Saad, S.; Pollock, C.; Oliver, B.; Al Odat, I.; Zaky, A.A.; Jones, N.; Chen, H. Impact of maternal cigarette smoke exposure on brain inflammation and oxidative stress in male mice offspring. Sci. Rep. 2016, 6, 25881. [Google Scholar] [CrossRef] [Green Version]
- Jandíková, H.; Dušková, M.; Stárka, L. The influence of smoking and cessation on the human reproductive hormonal balance. Physiol. Res. 2017, 66 (Suppl. 3), S323–S331. [Google Scholar]
- Csordas, A.; Wick, G.; Laufer, G.; Bernhard, D. An evaluation of the clinical evidence on the role of inflammation and oxidative stress in smoking-mediated cardiovascular disease. Biomark Insights 2008, 3, 127–139. [Google Scholar] [CrossRef]
- Gentner, N.J.; Weber, L.P. Secondhand tobacco smoke, arterial stiffness, and altered circadian blood pressure patterns are associated with lung inflammation and oxidative stress in rats. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H818–H825. [Google Scholar] [CrossRef] [Green Version]
- Anitha, T.S.; Srikanth, K.; Suganya, S.; Muthukumar, S.A. Comparative clinical study on the generation of nitrosative stress in cataractous lenses of smokers and non-smoker tobacco patients. Eur. J. Ophthalmol. 2019, 29, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Talhout, R.; Schulz, T.; Florek, E.; van Benthem, J.; Wester, P.; Opperhuizen, A. Hazardous compounds in tobacco smoke. Int. J. Environ. Res. Public Health. 2011, 8, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Hom, S.; Chen, L.; Wang, T.; Ghebrehiwet, B.; Yin, W.; Rubenstein, D.A. Platelet activation, adhesion, inflammation, and aggregation potential are altered in the presence of electronic cigarette extracts of variable nicotine concentrations. Platelets 2016, 27, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Oyeyipo, I.P.; Raji, Y.; Bolarinwa, A.F. Antioxidant profile changes in reproductive tissues of rats treated with nicotine. J. Hum. Reprod. Sci. 2014, 7, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Diemer, T.; Allen, J.A.; Hales, K.H.; Hales, D.B. Reactive oxygen disrupts mitochondria in MA-10 tumor Leydig cells and inhibits steroidogenic acute regulatory (StAR) protein and steroidogenesis. Endocrinology. 2003, 144, 2882–2891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedail, S.J.; Ahmed, A.A.; Musa, H.H.; Ismail, E.; Sifaldin, Z.A.; Musa, H.T. Gum arabic improves semen quality and oxidative stress capacity in alloxan induced diabetes rats. Asian Pac. J. Reprod. 2016, 5, 434–441. [Google Scholar] [CrossRef]
- Al Yahya, A.A.; Al Majed, A.A.; Gado, A.M.; Daba, M.H.; Al Shabanah, O.A.; Abd-Allah, A.R. Acacia senegal gum exudate offers protection against cyclophosphamide-induced urinary bladder cytotoxicity. Oxid. Med. Cell Longev. 2009, 2, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Valcarce, D.G.; Riesco, M.F.; Martínez-Vázquez, J.M.; Robles, V. Long exposure to a diet supplemented with antioxidant and anti-inflammatory probiotics improves sperm quality and progeny survival in the Zebrafish model. Biomolecules 2019, 9, 338. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters/Treatment | AE | HS | GA | HS + GA |
|---|---|---|---|---|
| Initial Body weight (g) | 21.4 ± 0.6 | 21.3 ± 0.62 | 20.8 ± 0.56 | 25.4 ± 0.65 |
| Final Body weight (g) | 22.9 ± 0.38 | 20.6 ± 0.56 | 20.6 ± 0.63 | 22.5 ± 0.42 |
| Body weight change (%) | 3.3 ± 2.5 | −2.8 ± 1.9 | −0.6 ± 1.2 | 1.0 ± 2.4 |
| Parameters/Treatment | AE | HS | GA | HS + GA |
|---|---|---|---|---|
| Testosterone (ng/mL) | 5.14 ± 0.3 | 2.0 ± 0.2 a | 5.2 ± 0.5 | 4.1 ± 0.2 b |
| Estradiol (pg/mL) | 28.57 ± 5.9 | 10.83 ± 0.6 a | 34.3 ± 2.58 | 17.8 ± 0.8 c |
| LH (mIU/mL) | 37.4 ± 1.2 | 28.2 ± 0.9 a | 41.0 ± 1.5 | 30.9 ± 1.0 b,c |
| Inhibin B (pg/mL) | 9.24 ± 0.71 | 66.45 ± 6.26 a | 6.44 ± 1.68 | 37.51 ± 6.22 b,c |
| ABP (ng/mL) | 9.67 ± 0.51 | 5.39 ± 0.51 a | 9.73 ± 0.96 | 8.32 ± 0.56 b |
| Group | Cotinine (ng/mL) |
|---|---|
| AE | 1.19 ± 0.13 |
| HS | 3.05 ± 0.6 a |
| GA | 1.11 ± 0.07 |
| GA + HS | 1.29 ± 0.09 b |
| Johnsen Score % | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 10 | 9 | 8 | 7 | 6 | 5 | 4 | 3 | 2 | 1 | |
| AE | 51 | 17 | 20 | 7 | 3 | 2 | 0 | 0 | 0 | 0 |
| HS | 39 | 17 | 21 | 4 | 4 | 15 | 0 | 0 | 0 | 0 |
| GA | 64 | 13 | 10 | 7 | 4 | 2 | 0 | 0 | 0 | 0 |
| HS + GA | 59 | 12 | 17 | 7 | 4 | 1 | 0 | 0 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, B.H.; Al-Salam, S.; Al Balushi, K.A.; Al Za’abi, M.; Adham, S.A.; Beegam, S.; Yuvaraju, P.; Manoj, P.; Nemmar, A. Ameliorative Effect of Gum Acacia on Hookah Smoke-Induced Testicular Impairment in Mice. Biomolecules 2020, 10, 762. https://doi.org/10.3390/biom10050762
Ali BH, Al-Salam S, Al Balushi KA, Al Za’abi M, Adham SA, Beegam S, Yuvaraju P, Manoj P, Nemmar A. Ameliorative Effect of Gum Acacia on Hookah Smoke-Induced Testicular Impairment in Mice. Biomolecules. 2020; 10(5):762. https://doi.org/10.3390/biom10050762
Chicago/Turabian StyleAli, Badreldin H., Suhail Al-Salam, Khalid A. Al Balushi, Mohammed Al Za’abi, Sirin A. Adham, Sumaya Beegam, Priya Yuvaraju, Priyadarsini Manoj, and Abderrahim Nemmar. 2020. "Ameliorative Effect of Gum Acacia on Hookah Smoke-Induced Testicular Impairment in Mice" Biomolecules 10, no. 5: 762. https://doi.org/10.3390/biom10050762
APA StyleAli, B. H., Al-Salam, S., Al Balushi, K. A., Al Za’abi, M., Adham, S. A., Beegam, S., Yuvaraju, P., Manoj, P., & Nemmar, A. (2020). Ameliorative Effect of Gum Acacia on Hookah Smoke-Induced Testicular Impairment in Mice. Biomolecules, 10(5), 762. https://doi.org/10.3390/biom10050762