Reduced Reverse Cholesterol Transport Efficacy in Healthy Men with Undesirable Postprandial Triglyceride Response

 ,
, 
Abstract
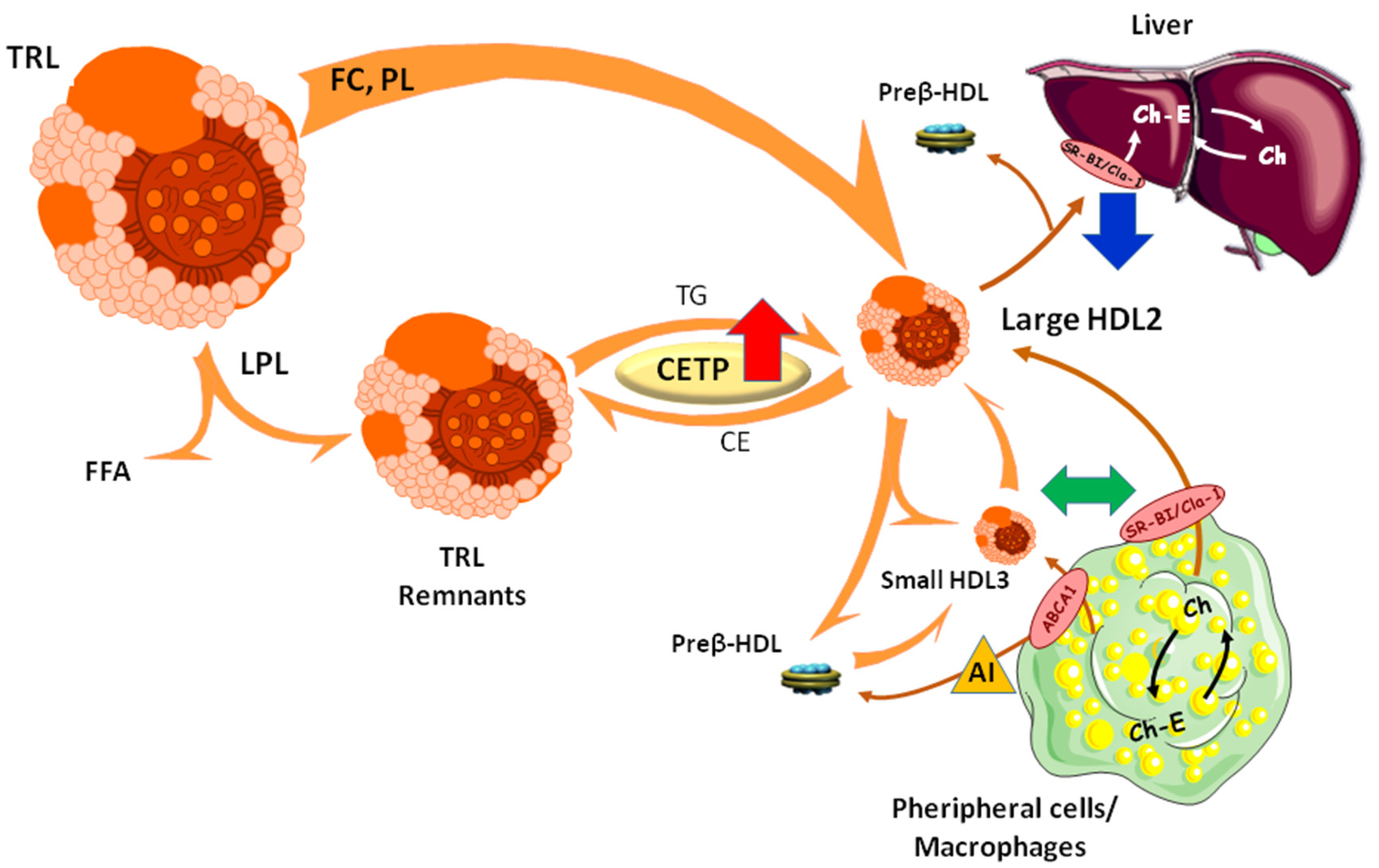
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Ethics
2.3. Study Design
2.4. Biochemical Analyses
2.5. Lipoprotein Fractionation
2.6. Determination of CETP Activity
2.7. Cholesterol Efflux Measurements
2.8. In Vitro Selective Hepatic Uptake of HDL-CE
2.9. Statistical Analyses
3. Results and Discussion
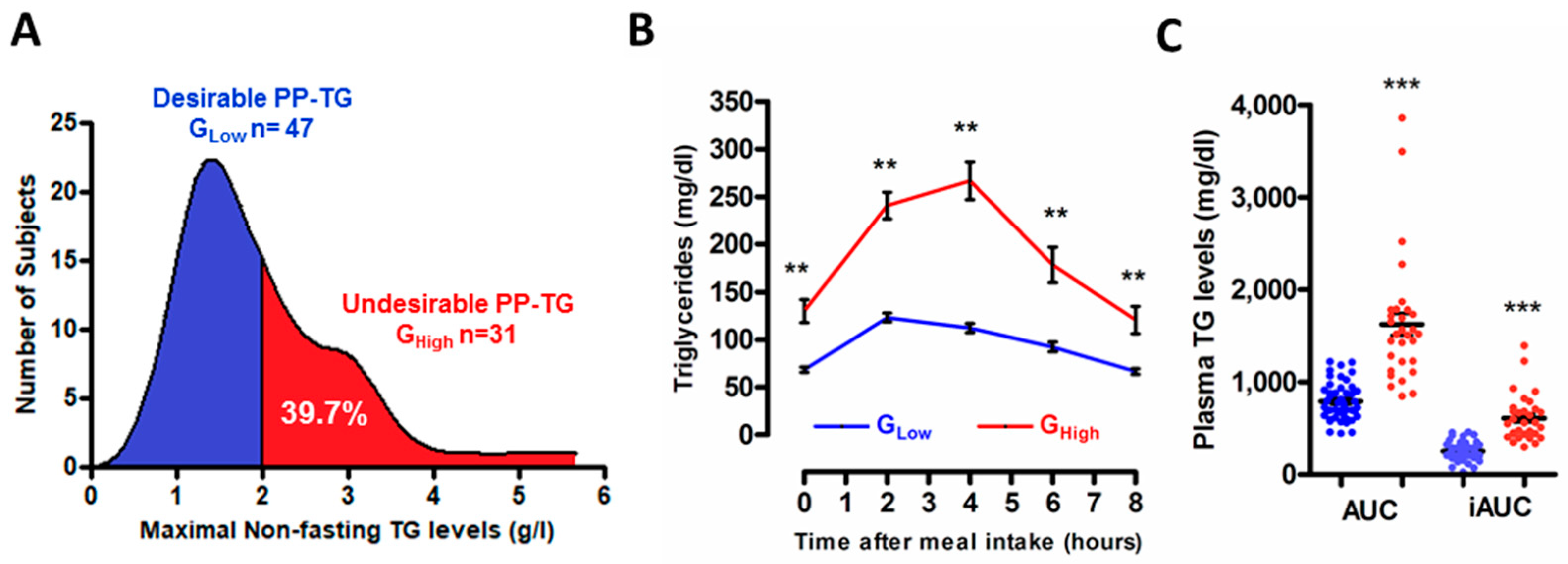
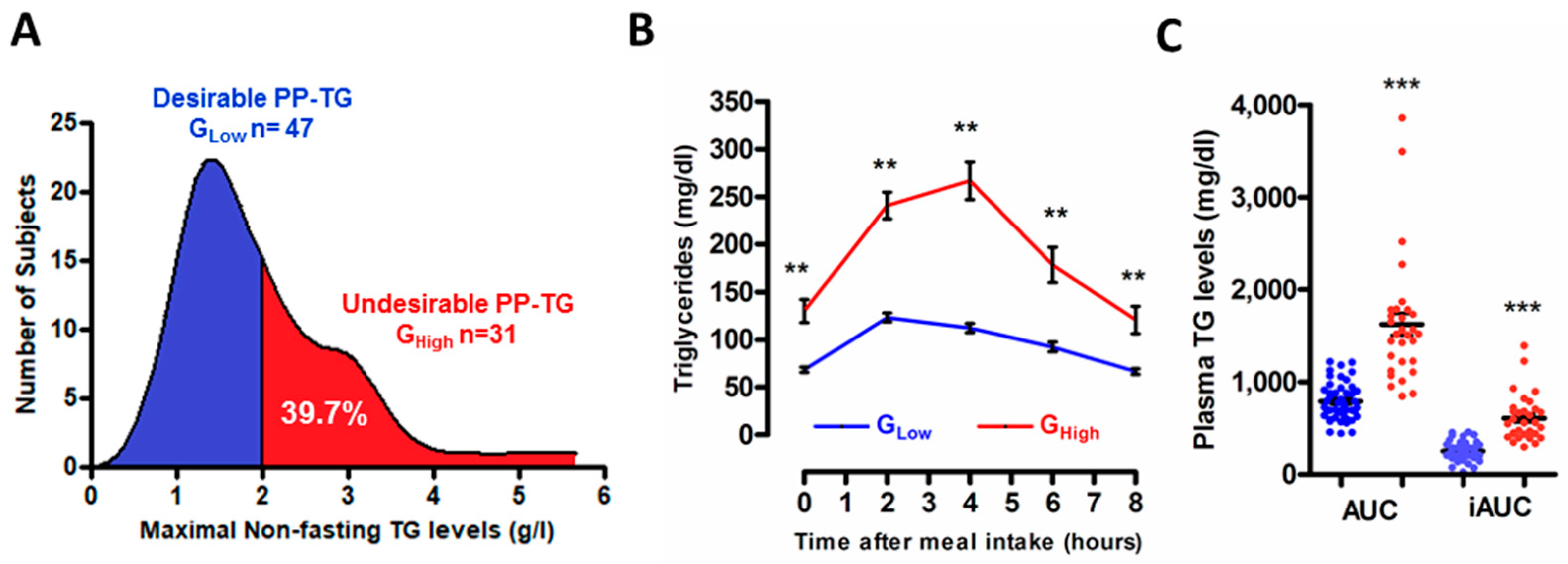
3.1. Stratification of the Study Population According to the Degree of the Postprandial Triglyceride Response
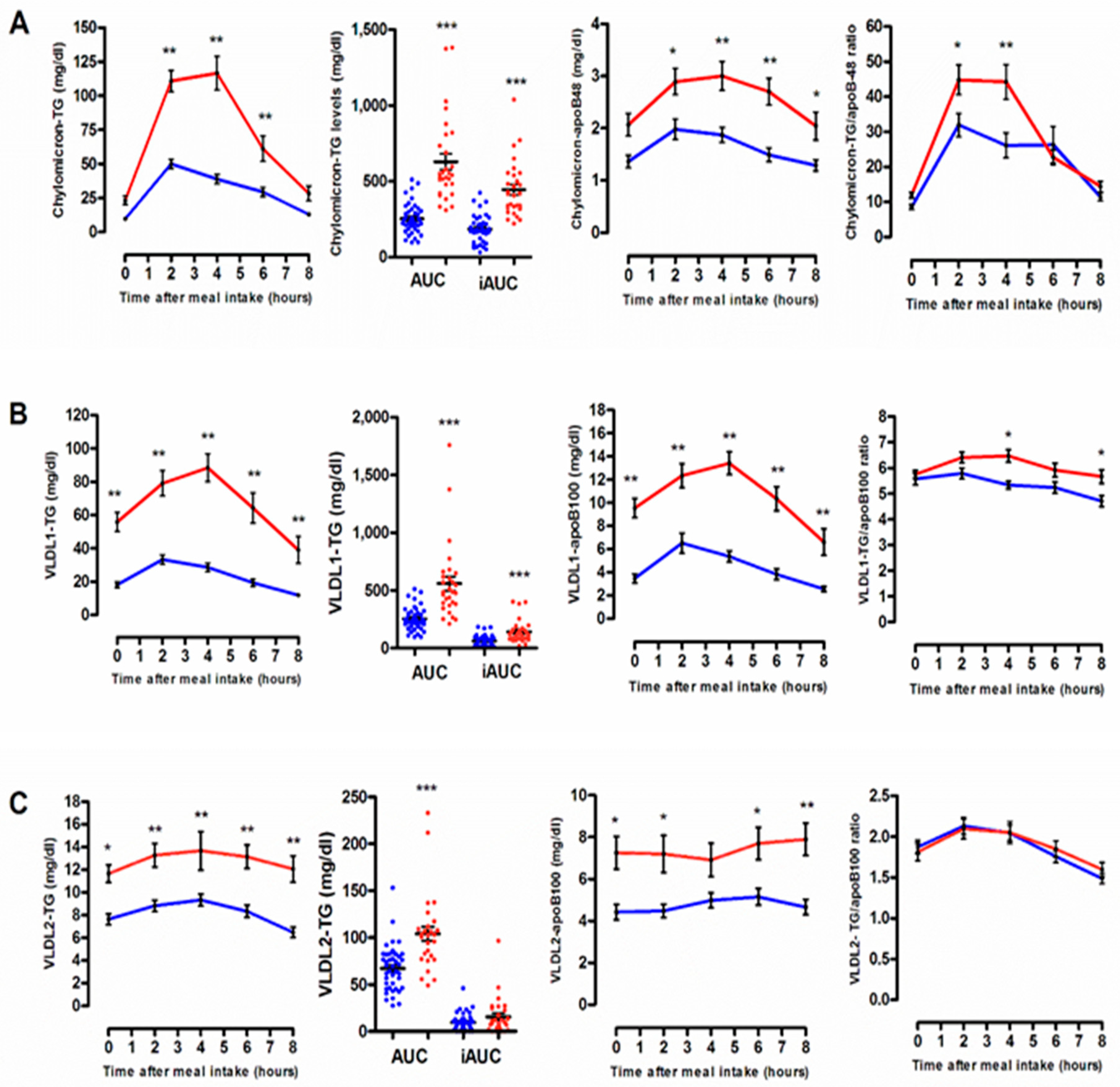
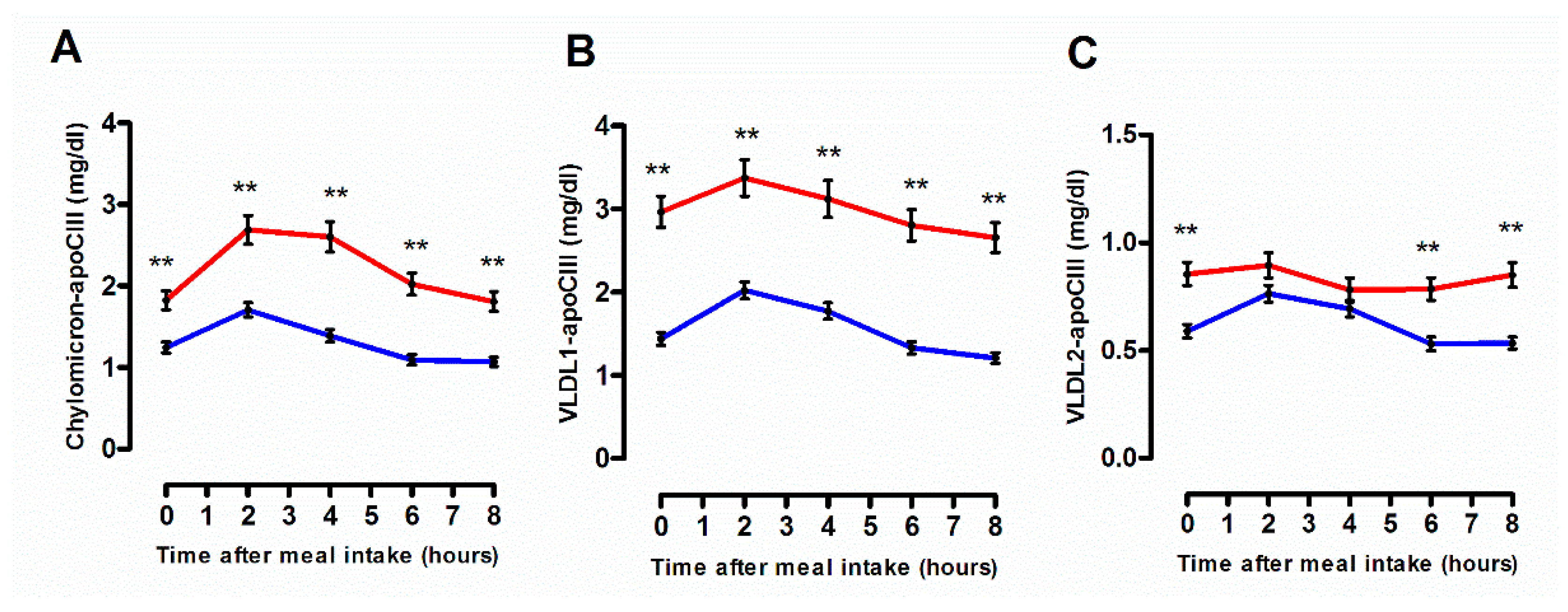
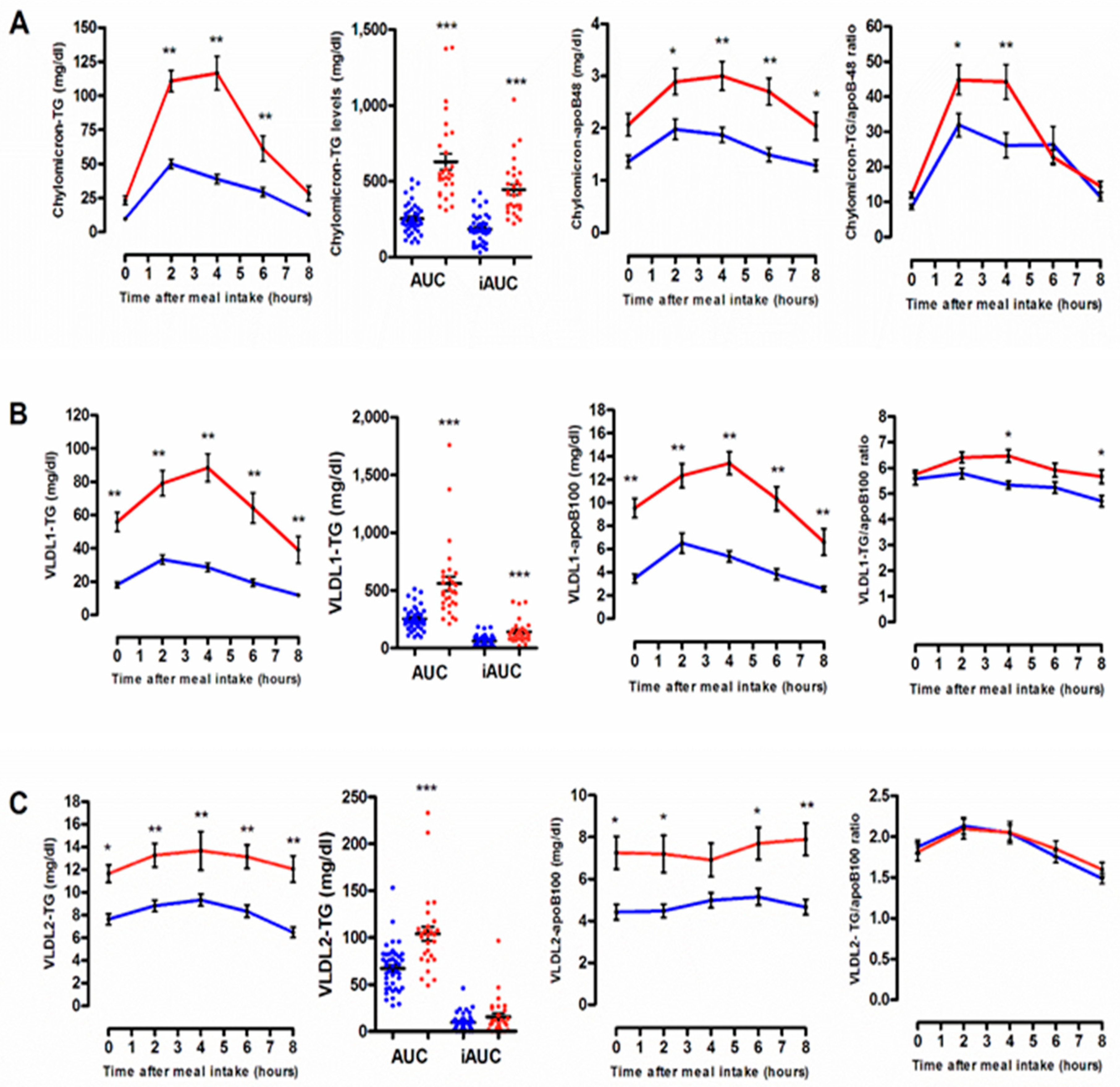
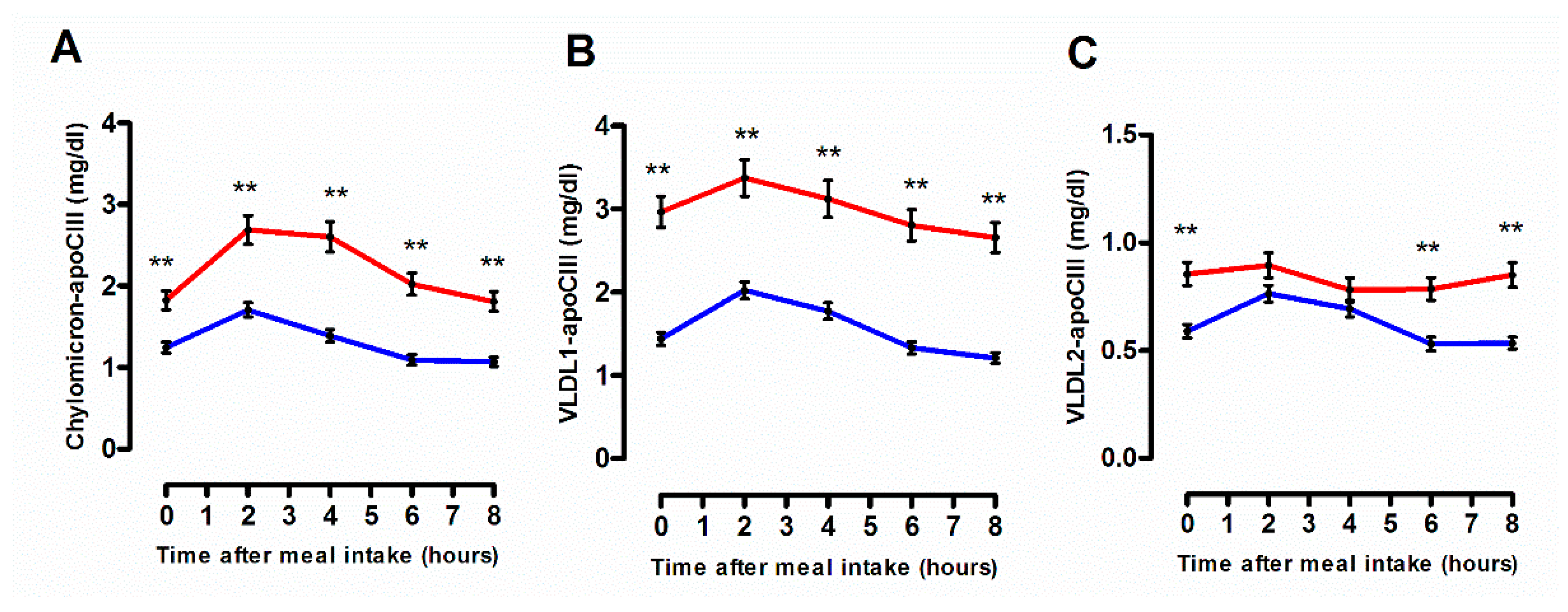
3.2. Quantitative and Qualitative Features of Plasma Lipoprotein Subspecies According to the Degree of the Postprandial Triglyceride Response
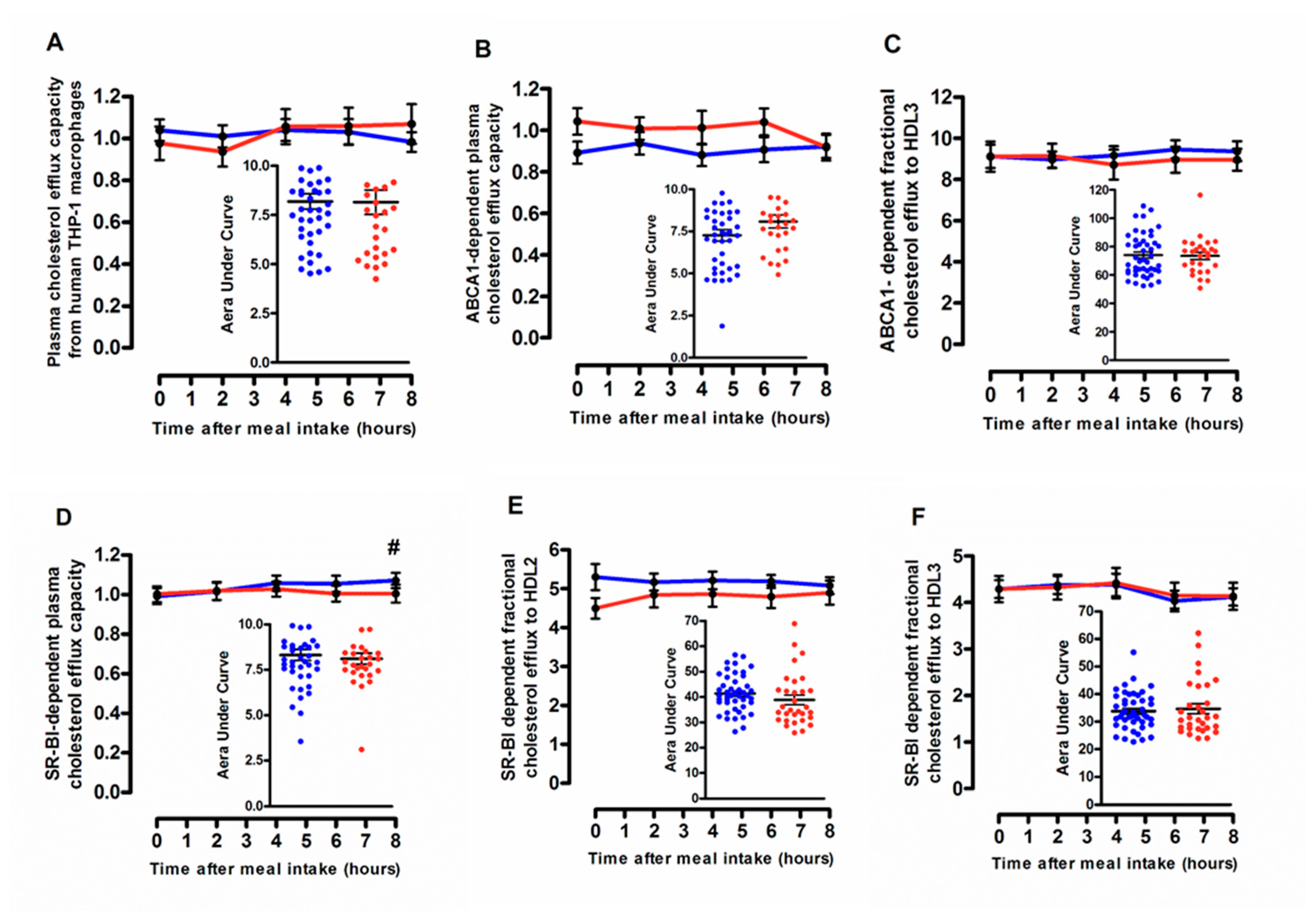
3.3. Cellular Free Cholesterol Efflux Capacity According to the Degree of the Postprandial Triglyceride Response
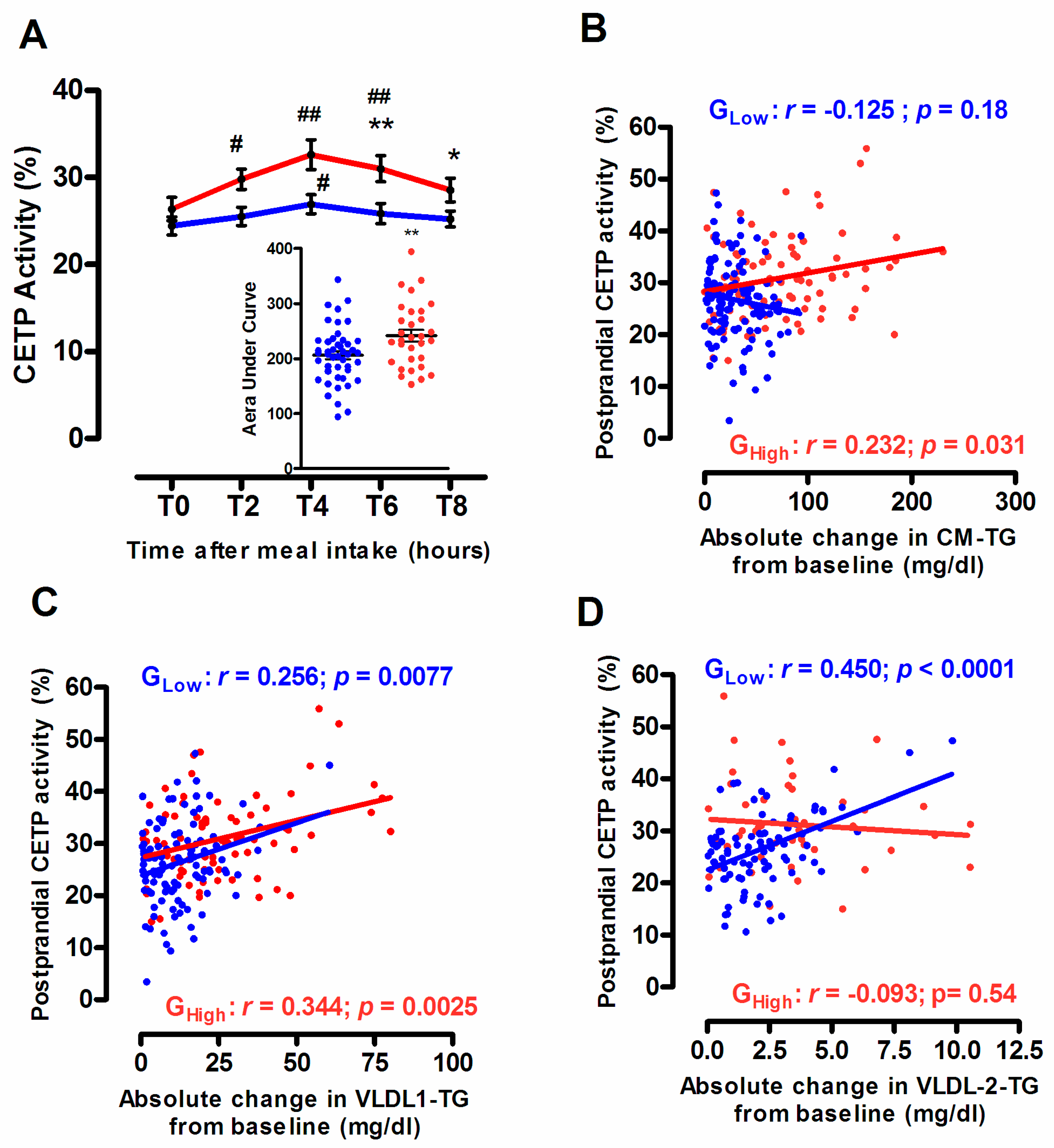
3.4. Endogenous CETP Activity According to the Degree of the Postprandial Triglyceride Response
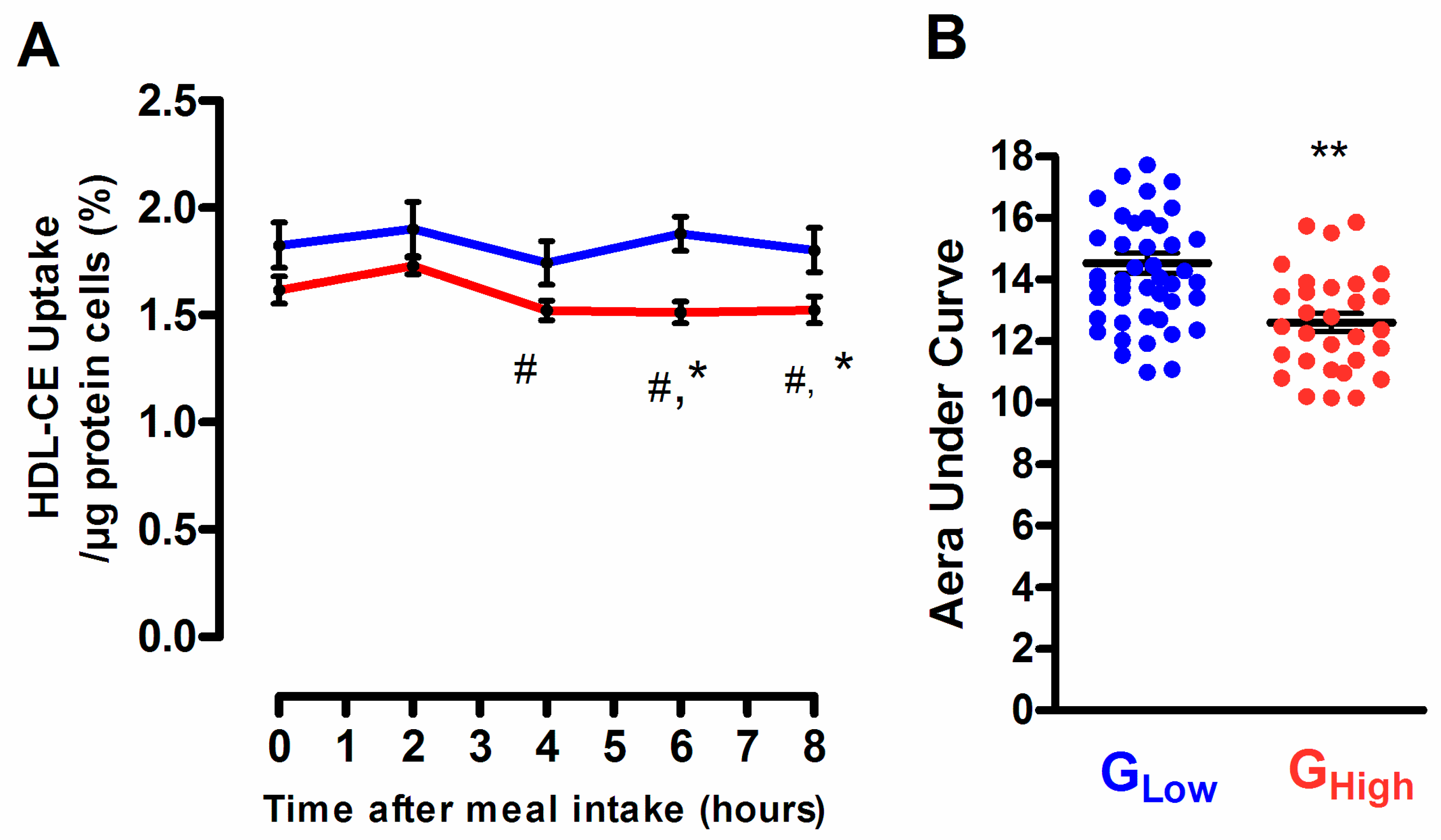
3.5. Selective HDL-CE Uptake to Hepatic Cells According to the Degree of the Postprandial Triglyceride Response
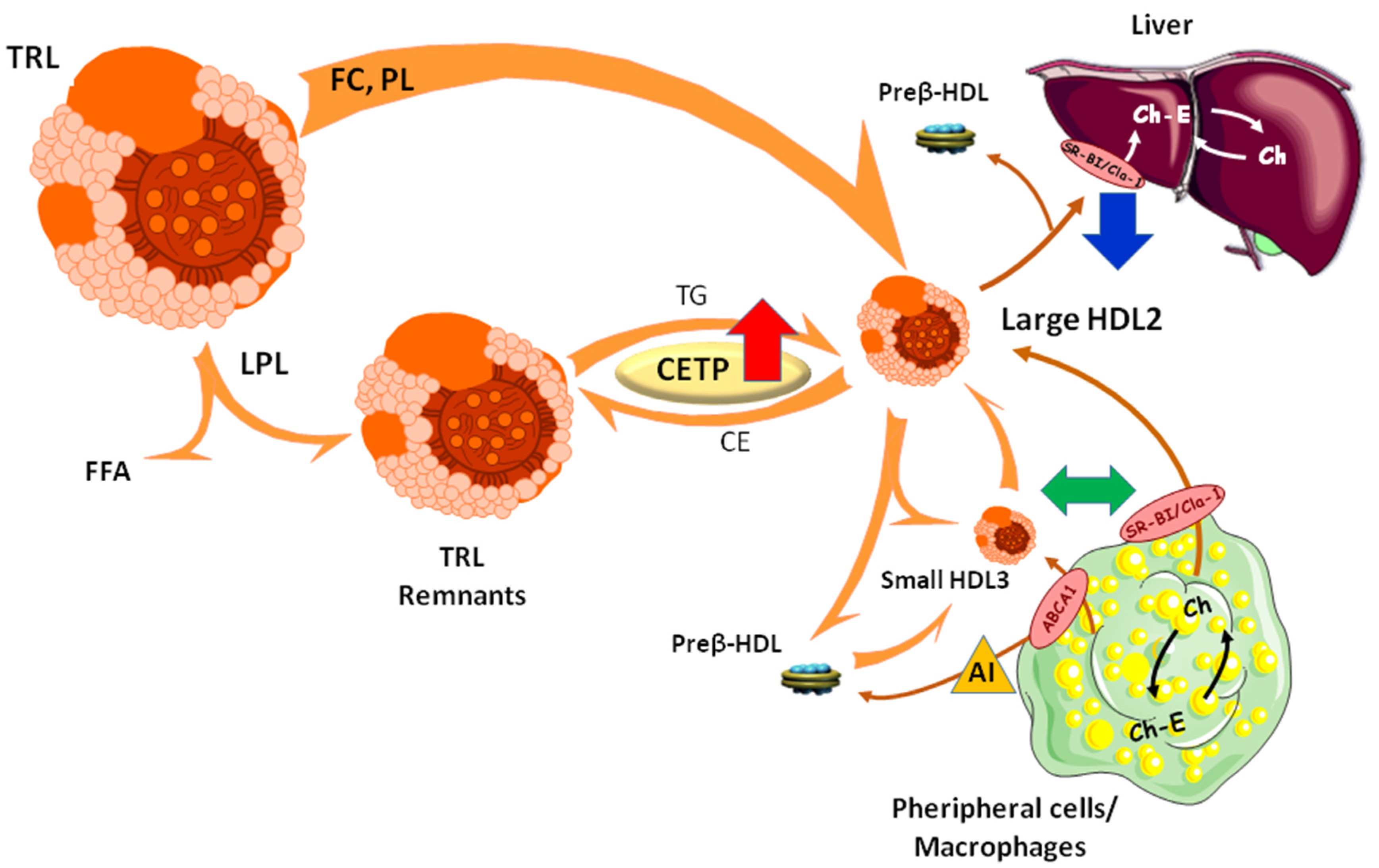
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RCT | Reverse Cholesterol Transport |
| AUC | Area under the curve |
| TRL | Triglyceride-rich lipoprotein |
| CM | Chylomicron |
| VLDL | Very-low density lipoprotein |
| LDL | Low-density lipoprotein |
| HDL | High-density lipoprotein |
| CETP | Cholesteryl ester transfer protein |
| Apo | Apolipoprotein |
References
- Khera, A.V.; Cuchel, M.; de la Llera-Moya, M.; Rodrigues, A.; Burke, M.F.; Jafri, K.; French, B.C.; Phillips, J.A.; Mucksavage, M.L.; Wilensky, R.L.; et al. Cholesterol efflux capacity, high-density lipoprotein function, and atherosclerosis. N. Engl. J. Med. 2011, 13, 127–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Eckardstein, A.; Nofer, J.R.; Assmann, G. High density lipoproteins and arteriosclerosis. Role of cholesterol efflux and reverse cholesterol transport. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 13–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, S.; Buring, J.E.; Rifai, N.; Mora, S.; Sacks, F.M.; Ridker, P.M. Fasting compared with nonfasting triglycerides and risk of cardiovascular events in women. JAMA 2007, 298, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordestgaard, B.G.; Benn, M.; Schnohr, P.; Tybjaerg-Hansen, A. Nonfasting triglycerides and risk of myocardial infarction, ischemic heart disease, and death in men and women. JAMA 2007, 298, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordestgaard, B.G.; Varbo, A. Triglycerides and cardiovascular disease. Lancet 2014, 384, 626–635. [Google Scholar] [CrossRef]
- Nakajima, K.; Nakano, T.; Tokita, Y.; Nagamine, T.; Inazu, A.; Kobayashi, J.; Mabuchi, H.; Stanhope, K.L.; Havel, P.J.; Okazaki, M.; et al. Postprandial lipoprotein metabolism: VLDL vs chylomicrons. Clin. Chim. Acta 2011, 412, 1306–1318. [Google Scholar] [CrossRef] [Green Version]
- Karpe, F.; Steiner, G.; Olivecrona, T.; Carlson, L.A.; Hamsten, A. Metabolism of triglyceride-rich lipoproteins during alimentary lipemia. J. Clin. Investig. 1993, 91, 748–758. [Google Scholar] [CrossRef] [Green Version]
- Castro, G.R.; Fielding, C.J. Effects of postprandial lipemia on plasma cholesterol metabolism. J. Clin. Investig. 1985, 75, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.; Sammett, D.; Granot, E. Mechanisms of enhanced cholesteryl ester transfer from high density lipoproteins to apolipoprotein B-containing lipoproteins during alimentary lipemia. J. Clin. Investig. 1986, 77, 1163–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassel, T.S.; Guerin, M.; Auboiron, S.; Chapman, M.J.; Guy-Grand, B. Preferential cholesteryl ester acceptors among triglyceride-rich lipoproteins during alimentary lipemia in normolipidemic subjects. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Kontush, A.; Chapman, M.J. Functionally defective high-density lipoprotein: A new therapeutic target at the crossroads of dyslipidemia, inflammation, and atherosclerosis. Pharmacol. Rev. 2006, 58, 342–374. [Google Scholar] [CrossRef] [PubMed]
- Bellanger, N.; Orsoni, A.; Julia, Z.; Fournier, N.; Frisdal, E.; Duchene, E.; Bruckert, E.; Carrie, A.; Bonnefont-Rousselot, D.; Pirault, J.; et al. Atheroprotective reverse cholesterol transport pathway is defective in familial hypercholesterolemia. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1675–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalano, G.; Julia, Z.; Frisdal, E.; Vedie, B.; Fournier, N.; Le Goff, W.; Chapman, M.J.; Guerin, M. Torcetrapib differentially modulates the biological activities of HDL2 and HDL3 particles in the reverse cholesterol transport pathway. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brites, F.D.; Bonavita, C.D.; De Geitere, C.; Cloës, M.; Delfly, B.; Yael, M.J.; Fruchart, J.; Wikinski, R.W.; Castro, G.R. Alterations in the main steps of reverse cholesterol transport in male patients with primary hypertriglyceridemia and low HDL-cholesterol levels. Atherosclerosis 2000, 152, 181–192. [Google Scholar] [CrossRef]
- Kolovou, G.D.; Anagnostopoulou, K.K.; Pavlidis, A.N.; Salpea, K.D.; Iraklianou, S.A.; Tsarpalis, K.; Damaskos, D.S.; Manolis, A.; Cokkinos, D.V. Postprandial lipemia in men with metabolic syndrome, hypertensives and healthy subjects. Lipids Health Dis. 2005, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellanger, N.; Julia, Z.; Villard, E.F.; El Khoury, P.; Duchene, E.; Chapman, M.J.; Fournier, N.; Le Goff, W.; Guerin, M. Functionality of postprandial larger HDL2 particles is enhanced following CETP inhibition therapy. Atherosclerosis 2012, 221, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Toth, P.P. Triglyceride-rich lipoproteins as a causal factor for cardiovascular disease. Vasc. Health Risk Manag. 2016, 12, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordestgaard, B.G. Triglyceride-Rich Lipoproteins and Atherosclerotic Cardiovascular Disease: New Insights from Epidemiology, Genetics, and Biology. Circ. Res. 2016, 118, 547–563. [Google Scholar] [CrossRef]
- Kolovou, G.D.; Mikhailidis, D.P.; Kovar, J.; Lairon, D.; Nordestgaard, B.G.; Ooi, T.C.; Perez-Martinez, P.; Bilianou, H.; Anagnostopoulou, K.; Panotopoulos, G. Assessment and clinical relevance of non-fasting and postprandial triglycerides: An expert panel statement. Curr. Vasc. Pharmacol. 2011, 9, 258–270. [Google Scholar] [CrossRef]
- Kolovou, G.D.; Watts, G.F.; Mikhailidis, D.P.; Pérez-Martínez, P.; Mora, S.; Bilianou, H.; Panotopoulos, G.; Katsiki, N.; Ooi, T.C.; Lopez-Miranda, J.; et al. Postprandial Hypertriglyceridaemia Revisited in the Era of Non-Fasting Lipid Profile Testing: A 2019 Expert Panel Statement, Main Text. Curr. Vasc. Pharmacol. 2019, 17, 498–514. [Google Scholar] [CrossRef]
- Guerin, M.; Egger, P.; Le Goff, W.; Soudant, C.; Dupuis, R.; Chapman, M.J. Atorvastatin reduces postprandial accumulation and cholesteryl ester transfer protein-mediated remodeling of triglyceride-rich lipoprotein subspecies in type IIb hyperlipidemia. J. Clin. Endocrinol. Metab. 2002, 87, 4991–5000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerin, M.; Lassel, T.S.; Le Goff, W.; Farnier, M.; Chapman, M.J. Action of atorvastatin in combined hyperlipidemia: Preferential reduction of cholesteryl ester transfer from HDL to VLDL1 particles. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villard, E.F.; Federspiel, M.C.; Cherfils, C.; Fesel-Fouquier, V.; Bruckert, E.; Clement, K.; Bonnefont-Rousselot, D.; Le Goff, W.; Bittar, R.; Couvert, P.; et al. Endogenous CETP activity as a predictor of cardiovascular risk: Determination of the optimal range. Atherosclerosis 2013, 227, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.; Silvain, J.; Gall, J.; Darabi, M.; Berthet, M.; Frisdal, E.; Hauguel-Moreau, M.; Zeitouni, M.; Kerneis, M.; Lattuca, B.; et al. Association of Serum Cholesterol Efflux Capacity With Mortality in Patients With ST-Segment Elevation Myocardial Infarction. J. Am. Coll. Cardiol. 2018, 72, 3259–3269. [Google Scholar] [CrossRef] [PubMed]
- Holzer, M.; Kern, S.; Trieb, M.; Trakaki, A.; Marsche, G. HDL structure and function is profoundly affected when stored frozen in the absence of cryoprotectants. J. Lipid Res. 2017, 58, 2220–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Diabetes Association. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42, S13–S28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredenrich, A.; Giroux, L.-M.; Tremblay, M.; Krimbou, L.; Davignon, J.; Cohn, J.S. Plasma lipoprotein distribution of apoC-III in normolipidemic and hypertriglyceridemic subjects: Comparison of the apoC-III to apoE ratio in different lipoprotein fractions. J. Lipid Res. 1997, 38, 1421–1432. [Google Scholar]
- Ginsberg, H.N.; Le, N.A.; Goldberg, I.J.; Gibson, J.C.; Rubinstein, A.; Wang-Iverson, P.; Norum, R.; Brown, W.V. Apolipoprotein B metabolism in subjects with deficiency of apolipoproteins CIII and AI: Evidence that apolipoprotein CIII inhibits catabolism of triglyceride-rich lipoproteins by lipoprotein lipase in vivo. J. Clin. Investig. 1986, 78, 1287–1295. [Google Scholar] [CrossRef]
- Jong, M.C.; Hofker, M.H.; Havekes, L.M. Role of ApoCs in lipoprotein metabolism: Functional differences between ApoC1, ApoC2, and ApoC3. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Weintraub, M.S.; Eisenberg, S.; Breslow, J.L. Dietary fat clearance in normal subjects is regulated by genetic variation in apolipoprotein E. J. Clin. Investig. 1987, 80, 1571–1577. [Google Scholar] [CrossRef]
- Marais, A.D. Apolipoprotein E in lipoprotein metabolism, health and cardiovascular disease. Pathology 2019, 51, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Stock, E.O.; Ferrara, C.T.; O’Connor, P.M.; Naya-Vigne, J.M.; Frost, P.H.; Malloy, M.J.; Kane, J.P.; Pullinger, C.R. Levels of prebeta-1 high-density lipoprotein are elevated in 3 phenotypes of dyslipidemia. J. Clin. Lipidol. 2018, 12, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Posadas-Sánchez, R.; Posadas-Romero, C.; Mendoza-Pérez, E.; Caracas-Portilla, N.A.; Cardoso-Saldaña, G.; Medina-Urrutia, A.; Jorge-Galarza, E.; Juárez-Rojas, J.G. Cholesterol efflux and metabolic abnormalities associated with low high-density-lipoprotein-cholesterol and high triglycerides in statin-treated coronary men with low-density lipoprotein-cholesterol <70 mg/dL. Am. J. Cardiol. 2012, 109, 636–641. [Google Scholar]
- Gall, J.; Frisdal, E.; Bittar, R.; Le Goff, W.; Bruckert, E.; Lesnik, P.; Guerin, M.; Giral, P. Association of Cholesterol Efflux Capacity With Clinical Features of Metabolic Syndrome: Relevance to Atherosclerosis. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Fournier, N.; Francone, O.; Rothblat, G.; Goudouneche, D.; Cambillau, M.; Kellner-Weibel, G.; Robinet, P.; Royer, L.; Moatti, N.; Simon, A.; et al. Enhanced efflux of cholesterol from ABCA1-expressing macrophages to serum from type IV hypertriglyceridemic subjects. Atherosclerosis 2003, 171, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Attia, N.; Ramaharo, A.; Paul, J.L.; Cambillau, M.; Beaune, P.; Grynberg, A.; Simon, A.; Fournier, N. Enhanced removal of cholesterol from macrophage foam cells to serum from type IV hypertriglyceridemic subjects. Atherosclerosis 2008, 198, 49–56. [Google Scholar] [CrossRef]
- Yassine, H.N.; Belopolskaya, A.; Schall, C.; Stump, C.S.; Lau, S.S.; Reaven, P.D. Enhanced cholesterol efflux to HDL through the ABCA1 transporter in hypertriglyceridemia of type 2 diabetes. Metabolism 2014, 63, 727–734. [Google Scholar] [CrossRef] [Green Version]
- Tatò, F.; Vega, G.L.; Tall, A.R.; Grundy, S.M. Relation between cholesterol ester transfer protein activities and lipoprotein cholesterol in patients with hypercholesterolemia and combined hyperlipidemia. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 112–120. [Google Scholar] [CrossRef]
- Mann, C.J.; Yen, F.T.; Grant, A.M.; Bihain, B.E. Mechanism of plasma cholesteryl ester transfer in hypertriglyceridemia. J. Clin. Investig. 1991, 88, 2059–2066. [Google Scholar] [CrossRef] [Green Version]
- Guerin, M.; Egger, P.; Soudant, C.; Le Goff, W.; van Tol, A.; Dupuis, R.; Chapman, M.J. Cholesteryl ester flux from HDL to VLDL-1 is preferentially enhanced in type IIB hyperlipidemia in the postprandial state. J. Lipid Res. 2002, 43, 1652–1660. [Google Scholar] [CrossRef] [Green Version]
- Guérin, M.; Dolphin, P.J.; Chapman, M.J. A new in vitro method for the simultaneous evaluation of cholesteryl ester exchange and mass transfer between HDL and apoB-containing lipoprotein subspecies. Identification of preferential cholesteryl ester acceptors in human plasma. Arterioscler. Thromb. 1994, 14, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, D.J.; Skeggs, J.W.; Morton, R.E. Elevated triglyceride content diminishes the capacity of high density lipoprotein to deliver cholesteryl esters via the scavenger receptor class B type I (SR-BI). J. Biol. Chem. 2001, 276, 4804–4811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Laccotripe, M.; Huang, X.; Rigotti, A.; Zannis, V.I.; Krieger, M. Apolipoproteins of HDL can directly mediate binding to the scavenger receptor SR-BI, an HDL receptor that mediates selective lipid uptake. J. Lipid Res. 1997, 38, 1289–1298. [Google Scholar] [PubMed]
- Bansal, N. Prediabetes diagnosis and treatment: A review. World J. Diabetes 2015, 6, 296–303. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Male Volunteers from the HDL-PP Cohort (n = 78) |
|---|---|
| Age, y | 30.1 ± 11.7 |
| BMI, kg/m² | 23.1 ± 2.7 |
| Waist circumference, cm | 83.8 ± 9.6 |
| Hip circumference, cm | 92.2 ± 8.0 |
| Waist to Hip ratio | 0.91 ± 0.07 |
| Systolic blood pressure, mmHg | 119.6 ± 9.0 |
| Diastolic blood pressure, mmHg | 77.2 ± 9.0 |
| Fasting blood glucose, mmol/L | 4.66 ± 0.45 |
| HbA1c, % | 5.32 ± 0.34 |
| Insulin, mU/L | 3.61 (1.61–6.34) |
| HOMA-IR | 0.70 (0.31–1.22) |
| Creatinine, µmol/L | 87.9 ± 9.23 |
| Creatinine Clearance, mL/min | 112.2 (101.9–122.7) |
| ASAT, UI/L | 25.9 ± 5.1 |
| ALAT, UI/L | 23.3 ± 9.6 |
| TSH, mUI/L | 1.73 ± 0.70 |
| hsCRP, mg/L | 0.58 (0.10–1.50) |
| Triglycerides, mg/dL | 64.1 (44.8–88.3) |
| Total cholesterol, mg/dL | 170.7 ± 32.6 |
| LDL-Cholesterol, mg/dL | 102.6 ± 27.6 |
| HDL-Cholesterol, mg/dL | 49.5 ± 11.9 |
| Remnant-Lipoprotein Cholesterol, mg/dL | 15.9 (12.3–22.8) |
| ApoAI, mg/dL | 127.2 ± 19.7 |
| ApoAII, mg/dL | 35.2 ± 6.12 |
| ApoB-48, mg/dL | 0.78 (0.54–1.21) |
| ApoB-100, mg/dL | 73.7 ± 24.2 |
| ApoE, mg/dL | 3.20 ± 1.33 |
| ApoCII, mg/dL | 4.02 ± 2.40 |
| ApoCIII, mg/dL | 7.30 ± 2.95 |
| Male Volunteers from the HDL-PP Cohort | |||
|---|---|---|---|
| Variables | Desirable PP-TG response (n = 47) | Undesirable PP-TG response (n = 31) | p-Value |
| Age, y | 28.3 ± 10.7 | 32.9 ± 12.7 | 0.0879 |
| BMI, kg/m² | 22.5 ± 2.2 | 24.1 ± 3.2 | 0.0207 |
| Waist circumference, cm | 81.5 ± 7.2 | 87.3 ± 11.7 | 0.0189 |
| Waist to Hip ratio | 0.89 ± 0.05 | 0.94 ± 0.08 | 0.0120 |
| Fasting blood glucose, mmol/L | 4.62 ± 0.43 | 4.73 ± 0.49 | 0.3181 |
| HbA1c, % | 5.25 ± 0.36 | 5.43 ± 0.28 | 0.0184 |
| Insulin, mU/L | 3.34 (1.00–5.77) | 4.02 (1.77–6.85) | 0.1443 |
| HOMA-IR | 0.68 (0.23–1.16) | 0.80 (0.41–1.51) | 0.1603 |
| Lipids, mg/dL | |||
| Triglycerides | 55.6 (42.1–67.3) | 87.9 (62.8–101.4) | <0.0001 |
| Total cholesterol | 163.3 ± 30.5 | 181.9 ± 33.0 | 0.0125 |
| LDL-Cholesterol | 97.5 ± 27.4 | 110.3 ± 26.6 | 0.0453 |
| HDL-Cholesterol | 52.0 ± 11.0 | 45.6 ± 12.5 | 0.0197 |
| Remnant-Lipoprotein Cholesterol | 13.1 (11.1–16.2) | 24.3 (18.0–28.3) | <0.0001 |
| Apolipoproteins, mg/dL | |||
| AI | 126.7 ± 18.5 | 127.9 ± 21.8 | 0.7911 |
| AII | 34.3 ± 5.3 | 36.6 ± 7.1 | 0.1060 |
| B-48 | 0.69 (0.46–0.95) | 1.06 (0.75–1.90) | 0.0109 |
| B-100 | 65.3 ± 19.1 | 86.3 ± 25.8 | <0.0001 |
| E | 3.03 ± 0.98 | 3.46 ± 1.71 | 0.2167 |
| CII | 3.55 ± 1.79 | 4.65 ± 2.95 | 0.1785 |
| CIII | 6.41 ± 2.30 | 8.79 ± 3.17 | 0.0008 |
| Chemical Component, % Weight | ||||||||
|---|---|---|---|---|---|---|---|---|
| Hours | Total Mass | FC | CE | TG | PL | Protein | CE/TG | |
| HDL2, d = 1.063–1.125 g/mL | ||||||||
| Desirable PP-TG response (GLow) | 0 | 153.2 ± 6.1 | 6.0 ± 0.2 | 23.7 ± 0.5 | 4.0 ± 0.2 | 21.8 ± 0.3 | 44.4 ± 0.5 | 5.97 |
| 2 | 164.2 ± 8.0 | 5.9 ± 0.2 | 22.8 ± 0.6 | 4.3 ± 0.2 | 22.2 ± 0.3 | 44.8 ± 0.4 | 5.29 | |
| 4 | 173.3 ± 7.1 # | 5.9 ± 0.2 | 22.5 ± 0.6 | 4.7 ± 0.2 ## | 22.5 ± 0.3 | 44.4 ± 0.5 | 4.79 ## | |
| 6 | 179.6 ± 5.7 ## | 6.0 ± 0.2 | 23.1 ± 0.7 | 4.5 ± 0.2 # | 22.4 ± 0.3 | 44.0 ± 0.5 | 5.10 | |
| 8 | 183.6 ± 6.8 ## | 5.9 ± 0.1 | 24.0 ± 0.5 | 3.8 ± 0.2 | 22.4 ± 0.2 | 43.9 ± 0.4 | 6.27 | |
| Undesirable PP-TG response (GHigh) | 0 | 150.6 ± 7.0 | 5.7 ± 0.2 | 22.2 ± 0.6 | 5.2 ± 0.3 * | 21.3 ± 0.3 | 45.6 ± 0.4 | 4.27 * |
| 2 | 155.8 ± 7.1 | 4.9 ± 0.2 | 22.7 ± 0.5 | 5.7 ± 0.3 * | 21.8 ± 0.3 | 44.8 ± 0.4 | 4.01 * | |
| 4 | 165.0 ± 7.8 # | 5.1 ± 0.2 | 20.9 ± 0.5 # | 6.8 ± 0.3 ##, ** | 21.8 ± 0.2 | 45.4 ± 0.4 | 3.09 ##, * | |
| 6 | 163.2 ± 6.5 # | 5.1 ± 0.2 | 20.9 ± 0.6 # | 6.1 ± 0.3 ##, ** | 22.4 ± 0.2 | 45.4 ± 0.5 | 3.40 ##, * | |
| 8 | 168.9 ± 6.9 ## | 5.2 ± 0.2 | 21.5 ± 0.6 * | 5.2 ± 0.3 * | 22.2 ± 0.3 | 45.8 ± 0.4 | 4.12 * | |
| HDL3, d = 1.125–1.21 g/mL | ||||||||
| Desirable PP-TG response (GLow) | 0 | 140.4 ± 3.8 | 3.0 ± 0.1 | 16.1 ± 0.3 | 3.6 ±0.1 | 18.3 ± 0.4 | 58.9 ± 0.5 | 4.47 |
| 2 | 136.0 ± 2.7 | 3.1 ± 0.2 | 14.9 ± 0.4 # | 3.8 ± 0.2 | 18.8 ± 0.3 | 59.3 ± 0.5 | 3.89 # | |
| 4 | 134.7 ± 3.2 | 3.2 ± 0.2 | 14.0 ± 0.4 ## | 4.1 ± 0.2 # | 18.9 ± 0.3 | 59.7 ± 0.5 | 3.41 ## | |
| 6 | 134.0 ± 3.0 | 3.2 ± 0.1 | 14.3 ± 0.4 ## | 4.3 ± 0.2 ## | 18.9 ± 0.3 | 59.2 ± 0.5 | 3.31 ## | |
| 8 | 140.1 ± 3.7 | 3.1 ± 0.1 | 15.2 ± 0.4 | 3.9 ± 0.2 | 19.3 ± 0.3 | 58.5 ± 0.5 | 3.91 # | |
| Undesirable PP-TG response (GHigh) | 0 | 153.7 ± 4.1 | 3.1 ± 0.2 | 16.1 ± 0.6 | 4.3 ± 0.2 | 17.8 ± 0.3 | 58.4 ± 0.5 | 3.71 |
| 2 | 145.1 ± 4.5 | 2.9 ± 0.1 | 15.2 ± 0.4 | 4.8 ± 0.2 * | 18.2 ± 0.4 | 58.9 ± 0.5 | 3.16 # | |
| 4 | 145.7 ± 4.0 | 2.9 ± 0.1 | 13.5 ± 0.6 ## | 5.2 ± 0.2 #, * | 18.0 ± 0.5 | 60.4 ± 1.0 | 2.58 ## | |
| 6 | 140.8 ± 3.3 | 2.8 ± 0.1 | 13.9 ± 0.4 ## | 5.2 ± 0.2 #, * | 18.9 ± 0.4 | 59.2 ±0.5 | 2.69 ## | |
| 8 | 142.3 ± 3.6 | 3.1 ± 0.1 | 14.2 ± 0.5 | 4.7 ± 0.2, * | 18.8 ± 0.4 | 59.2 ± 0.6 | 2.98 ## | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motte, A.; Gall, J.; Salem, J.-E.; Dasque, E.; Lebot, M.; Frisdal, E.; Galier, S.; Villard, E.F.; Bouaziz-Amar, E.; Lacorte, J.-M.; et al. Reduced Reverse Cholesterol Transport Efficacy in Healthy Men with Undesirable Postprandial Triglyceride Response. Biomolecules 2020, 10, 810. https://doi.org/10.3390/biom10050810
Motte A, Gall J, Salem J-E, Dasque E, Lebot M, Frisdal E, Galier S, Villard EF, Bouaziz-Amar E, Lacorte J-M, et al. Reduced Reverse Cholesterol Transport Efficacy in Healthy Men with Undesirable Postprandial Triglyceride Response. Biomolecules. 2020; 10(5):810. https://doi.org/10.3390/biom10050810
Chicago/Turabian StyleMotte, Alexandre, Julie Gall, Joe-Elie Salem, Eric Dasque, Martine Lebot, Eric Frisdal, Sophie Galier, Elise F. Villard, Elodie Bouaziz-Amar, Jean-Marc Lacorte, and et al. 2020. "Reduced Reverse Cholesterol Transport Efficacy in Healthy Men with Undesirable Postprandial Triglyceride Response" Biomolecules 10, no. 5: 810. https://doi.org/10.3390/biom10050810
APA StyleMotte, A., Gall, J., Salem, J.-E., Dasque, E., Lebot, M., Frisdal, E., Galier, S., Villard, E. F., Bouaziz-Amar, E., Lacorte, J.-M., Charbit, B., Le Goff, W., Lesnik, P., & Guerin, M. (2020). Reduced Reverse Cholesterol Transport Efficacy in Healthy Men with Undesirable Postprandial Triglyceride Response. Biomolecules, 10(5), 810. https://doi.org/10.3390/biom10050810